
Recent insights into physiological responses to nutrients by the cylindrospermopsin producing cyanobacterium,Cylindrospermopsis raciborskii*
2018-08-02 02:50MicheleBURFORDAnusuyaWILLISAnnCHUANGXiaoMANPhilipORR
Michele A. BURFORD , Anusuya WILLIS, Ann CHUANG, Xiao MAN, Philip T. ORR
Australian Rivers Institute, Griffith University, Australia
Abstract The harmful cyanobacterium Cylindrospermopsis raciborskii is a widespread species increasingly being recorded in freshwater systems around the world. Studies have demonstrated some key attributes of this species which may explain its global dominance. It has a high level of flexibility with respect to light and nutrients, being capable of growth under low and variable light conditions. However,it is the strategy with respect to nutrient utilization that has received more attention. Unlike many bloom forming species, the dominance of this species is not simply linked to higher nutrient loads. In fact it appears that it is more competitive when phosphorus and nitrogen availability is low and/or variable. An important component of this flexibility appears to be the result of within-population strain variability in responses to nutrients, as well as key physiological adaptations. Strain variability also appears to have an effect on the population-level cell quota of toxins, speci fically cylindrospermopsins (CYNs). Field studies in Australia showed that populations had the highest proportion of toxic strains when dissolved inorganic phosphorus was added, resulting in stoichiometrically balanced nitrogen and phosphorus within the cells. These strategies are part of an arsenal of responses to environmental conditions, making it a challenging species to manage.However, our ability to improve bloom prediction will rely on a more detailed understanding of the complex physiology and ecology of this species.
Keyword: nitrogen; phosphorus; cylindrospermopsins; strains
1 INTRODUCTION
Much of the focus on drivers of cyanobacterial blooms has been on nutrient loads. It is well established that excessive nutrient loads are responsible for many of the world’s algal blooms,including affecting the duration and magnitude of blooms (Glibert and Burford, 2017). At the simplest level, scienti fic thinking is that more nutrient inputs equals more frequent, bigger, longer-lasting blooms.However, we also now know that the relationship between nutrient pollution and blooms is more complex than traditionally thought. This is highlighted by the inability to accurately predict blooms (see Glibert et al., 2010; Oliver et al., 2012).
The success of a cyanobacterial blooms re flects physiological adaptations of species and strains within species, environmental conditions and interactions with co-occurring organisms. The toxic cyanobacterium, Cylindrospermopsis raciborskii is no exception. It is increasingly being recorded as present or in bloom proportions in lakes, reservoirs and rivers throughout the world (Padisák, 1997; Sinha et al., 2014). In Australia it produces the hepato- and cytotoxins, cylindrospermopsins (CYNs). It was first deemed a harmful bloom species after a toxic bloom event in 1979 when human casualties developed symptoms including vomiting and tender hepatomegaly (Byth, 1980; Hawkins et al., 1985). We now know that CYNs are also produced by strains in other areas of the world, e.g. New Zealand (Wood and Stirling, 2003) and Asia (Li et al., 2001; Wimmer et al., 2014). In contrast, in South America, strains produce saxitoxins while in other regions of the world, species may be non-toxic (Lagos et al., 1999;Neilan et al., 2003). Much of our understanding of this species has focussed on Australia and South America due to the toxic nature of blooms in these regions.
In recent years, scienti fic studies of this species has increased signi ficantly. In 2006, Web of Science had 24 publications with C. raciborskii as a topic, while ten years later this was up to 78. Much of the research has focussed on understanding the drivers of blooms via ecological, physiological and molecular studies.As our knowledge grows, some key themes are emerging. They include the challenge of understanding physiological responses in the face of a high level of variability in responses to environmental conditions.Additionally, the role of nutrients in promoting the species, and its toxin levels have received attention in an attempt to manage blooms. This review focusses on these topics, while acknowledging that there are other drivers, e.g. physical conditions, allelopathy(e.g. Figueredo et al., 2007; Rzymski et al., 2014)which are also likely to play a key role.
2 A FOCUS ON STRAINS
A review of the species by Burford et al. (2016)examined the role of a range of environmental conditions, including nutrients, in promoting blooms of C. raciborskii. Comparison of field studies of this species has given con flicting results regarding the importance of nutrients in promoting blooms. In the case of phosphorus, field and bioassay studies have shown both positive and negative correlations between phosphorus concentrations and C. raciborskii cell densities (Bonilla et al., 2012; Kokociński and Soininen, 2012; Muhid et al., 2013; Soares et al.,2013; Aguilera et al., 2017). Chislock et al. (2014)also showed that C. raciborskii can dominate under a range of phosphorus levels: Posselt et al. (2009) also found this but only when nitrogen levels were not elevated. Additionally, Dolman et al. (2012) showed higher C. raciborskii cell concentrations at high nitrogen:phosphorus ratios, relative to Red field(1958) ratios of 16:1. Therefore, there are no consistent findings regarding nutrient availability and C. raciborskii blooms from these field studies.
The use of culture studies to examine nutrient utilization strategies is also problematic. A review by Lakeman et al. (2009) examined the issue of strain variability in phytoplankton cultures. There are many challenges in interpreting culture studies in an ecological context, including changes in physiological responses of strains as a result of genetic changes after an extended period of culturing, and differences in culture conditions.
Strains can also vary in their physiological responses across geographic ranges, but a recent study has also shown that strains also within waterbodies. For example, Willis et al. (2016a) found that 24 isolates of C. raciborskii from one bottle of lake water, grown under the same environmental conditions, had a 2.5 fold difference in their CYNs cell quotas. Additionally, there was variation in the ratio of two forms of CYNs present in strains, as well as the growth rate and morphology.
A recent study also showed that strain variability can be just as signi ficant as species differences in determining the growth response to light and temperature (Xiao et al., 2017). This study compared responses of eight C. raciborskii strains and four Microcystis aeruginosa strains isolated across two adjacent reservoirs. The data from this study was then modelled to show that the winners of competition between species can be highly variable, depending on strains present, as well as light and temperature conditions (M. Xiao unpubl. data). There was no absolute ‘winner’ under all conditions as there were always strains predicted to coexist with the dominant strain. The uncertainty in prediction of species competition outcomes was due to the substantial variability of growth responses within and between strains.
The fact that such a degree of variability occurs with strains within populations may also help explain why there is so much variationin responses of isolates from different geographic regions to the same physicochemical variables, including nutrients, in published studies. Additionally, in the case of C. raciborskii, it seems that a key attribute that contributes to its success is the high level of physiological variability between strains, giving this species flexibility under variable environmental conditions.
3 NITROGEN
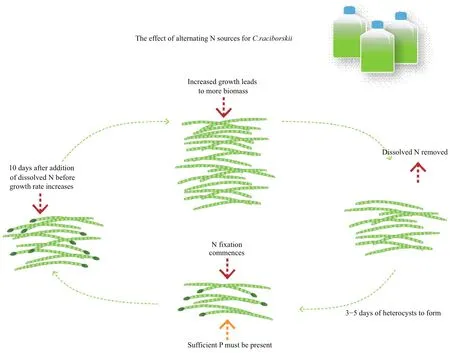
Fig.1 Diagram showing utilization of dissolved nitrogen and nitrogen fixation under a range of conditions by C. raciborskii(from Willis et al., 2015, 2016b)
The ability of cyanobacteria to fix atmospheric nitrogen has often been proposed as a key factor explaining cyanobacterial dominance in freshwater systems where dissolved nitrogen levels are low.Cylindrospermopsis raciborskii also has the ability to fix nitrogen, but studies have questioned whether it promotes C. raciborskii blooms (Moisander et al.,2012). There is a preference for dissolved inorganic(ammonium, nitrate) and organic (urea) forms of nitrogen over nitrogen fixation, as demonstrated by a range of studies (Saker and Neilan, 2001; Burford et al., 2006; Ammar et al., 2014; Stucken et al., 2014).This species remains effective at scavenging dissolved nitrogen, even under low concentrations. Nitrogen fixation is an energetically expensive process so use of dissolved forms of nitrogen, especially under lower light conditions, is an efficient strategy. As a result,dissolved forms may provide the nitrogen source throughout much of the year (Burford et al., 2006).However, despite nitrogen fixation being energetically expensive, it remains a useful strategy under dissolved nitrogen stress giving a competitive advantage over non- fixing cyanobacteria and eukaryotic algae.
The role of nitrogen fixation in supporting growth and dominance of C. raciborskii has been examined in more detail in culture studies. It appears that nitrogen fixation is a relatively inefficient process. Heterocytes are required for nitrogen fixation but a study showed that it can take 3–5 days for development of heterocytes once dissolved nitrogen becomes limiting (Fig.1,Willis et al., 2016b). Cylindrospermopsis raciborskii produces only terminal heterocytes but can compensate with highly efficient transfer of nitrogen from the heterocyte to, and between vegetative cells in the filament (Plominsky et al., 2013, 2015). Once heterocytes are produced, nitrogen fixation has been shown to occur immediately, but growth is slower than with dissolved nitrogen (Fig.1). Subsequent addition of growth-media level concentrations of dissolved nitrogen caused heterocyte loss after about 5 days, halting nitrogen fixation (Kenesi et al., 2009;Willis et al., 2016b). This contrasted with a field mesocosm study where lower levels of dissolved nitrogen addition increased heterocyte production,provided sufficient phosphorus was available (Burford et al., 2014). These results also highlight that culture studies can give a distorted view of how cells respond physiologically to nutrients under typical field concentrations.
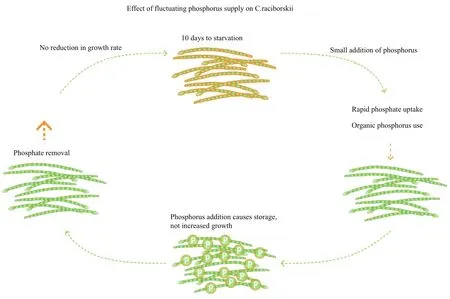
Fig.2 Diagram showing effect of fluctuating phosphorus availability on C. raciborskii (from Willis et al., 2015, 2017)
The length of trichomes, or cells per trichome, vary between strains of C. raciborskii with implications for nitrogen fixation (Willis et al., 2016b). In some studies, trichome length has been found to change with environmental conditions (Plominsky et al.,2013; Amaral et al., 2014), while in others it appears to be constant (Willis et al., 2015). As a result of producing only terminal heterocytes (Plominsky et al., 2013), variation in trichome length results in differences in the number of vegetative cells supplied with nitrogen from each heterocyte (Willis et al.,2015). This may mean that strains with long trichomes have a lower nitrogen supply under N- fixing conditions, which could result in a lower growth rate,and disadvantage these cells. Conversely, heterocyte and nitrogenase production are energetically expensive, thus a lower number of vegetative cells per heterocyte could be a disadvantage. Indeed one study found the growth rate of cultures under N- fixing conditions was higher in the strain with longer trichomes than others (Willis et al., 2015). More research is needed to more fully elucidate these strain differences. For example, although the terminal heterocytes of C. raciborskii appear to have a higher nitrogenase activity compared to Dolichospermum,which has intercalary heterocytes (Plominsky et al.,2013), it is unknown if strains vary in their heterocyte N- fixing capacity.
These studies highlight that bloom formation is most likely in systems with sufficient dissolved nitrogen availability. Nitrogen fixation only occurs if there are extended periods of dissolved nitrogen limitation, and is only used as a mechanism for cell maintenance.
4 PHOSPHORUS
The early paradigm of phosphorus utilization by cyanobacteria (and other microalgae) was encapsulated in two key empirical equations: Monad and Droop (Monod, 1942; Droop, 1983). The theory is that algal biomass increases as phosphorus availability (internal and external) increases until a maximum biomass is reached. However, it is increasingly clear that many species do not adhere to these relationships. Cylindrospermopsis raciborskii is an example of this, having a range of strategies to maximise utilization of phosphorus (Fig.2). Indeed, it appears from these studies that the success of this species is the result of adaptability to low and variable phosphorus supply, rather than the ability to outcompete other species under increasing phosphorus availability.
Some years ago researchers became aware that there are cyanobacterial species that can alternate between passive and active dissolved inorganic phosphorus uptake in response to changes in phosphorus availability (Falkner et al., 1995; Richie et al., 2001). Passive uptake occurred at higher dissolved inorganic phosphorus availability, while below a threshold, active uptake was the mechanism for dissolved inorganic phosphorus use. This resulted in much more rapid uptake of dissolved inorganic phosphorus than passive uptake.
Early work on phosphorus utilization by a European strain by C. raciborskii showed that it is capable of uptake rates of high dissolved inorganic phosphorus(Isvánovics et al., 2000). More recently, studies with Australian strains of C. raciborskii measured the highest reported dissolved inorganic phosphorus uptake rates for a cyanobacterial species, re flecting active uptake (Willis et al., 2017). This was substantiated in studies showing upregulation of the active uptake genes, Pst (A. Willis unpubl. data). In the field, a threshold concentration of 5 μg/L dissolved inorganic phosphorus was identi fied as a switch from passive to active uptake in C. raciborskii-dominated populations (Prentice et al., 2015).
Other studies have also shown that pulses of dissolved inorganic phosphorus, rather than constant additions, result in faster growth of C. raciborskii(Marinho et al., 2013; Amaral et al., 2014). Indeed,studies of C. raciborskii with constant dissolved inorganic phosphorus addition showed no change in growth rate until starvation occurred, at which time cultures died (Willis et al., 2017). This suggests that the mechanism used by C. raciborskii to provide sufficient phosphorus is phosphorus storage, rather than varying growth rates. Conversely the relatively high concentrations of phosphorus typically used in growth media can result in detrimental effects on C. raciborskii growth, e.g. Willis et al. (2015) found highest growth rates when growth media concentrations were half or lower. Studies have also shown the ability to utilize organic phosphorus as an alternative source, when dissolved inorganic phosphorus was depleted (Bai et al., 2014). This is not, however, unique to C. raciborskii, as it has been described for a number of cyanobacterial species.
Strains of C. raciborskii vary in their phosphorus storage capacity and carbon:phosphorus ratios, and these physiological differences result in differences in the amount of time cultures can grow in P-free medium, and at a maximum growth rate (Willis et al.,2017). Amaral et al. (2014), demonstrated strain differences in response to P additions, with strain MVCC19 having a more flexible physiology (i.e.changes in μmax) compared to CCMP1973, which conversely had bigger changes in morphology.Differences in alkaline phosphatase activity has also been found to vary between strains, with Chinese strains showing an increase in activity with P starvation (Wu et al., 2012), while Australian strains upregulated the alkaline phosphatase gene but did not increase activity in response to P starvation (A. Willis,unpubl. data). The genome composition, in terms of P metabolism genes, is identical in all strains analyzed so far (Sinha et al., 2014), so it is likely that differential transcription causes the differences in how strains use phosphorus.
There is also an interaction between phosphorus availability and population-level toxin quotas in C. raciborskii. A study by Burford et al. (2014) found a shift in dominance towards toxic strains with phosphorus addition to mesocosms, possibly as a result of a change to a balanced nitrogen:phosphorus stoichiometry in cells. This contrasts with nitrogen addition, where there is no effect on toxin production,or strain dominance. This also contrasts with another well studied toxic species, Microcystis aeruginosa,where the addition of nitrogen has been shown to preferentially promote toxic strains (Gobler et al.,2016).
Zooplankton grazers can have both negative and positive effects on cyanobacteria. The negative effect is grazing which reduces cell densities, while positive effects include preferentially grazing noncyanobacterial algal species, thus promoting cyanobacteria. Additionally, grazers can excrete nutrients when grazing, promoting algal, including cyanobacterial growth. In the case of C. raciborskii, it appears to gain an advantage from the presence of phosphorus excreted by zooplankton. Studies in both the laboratory and mesocosms in an Australian reservoir have shown that zooplankton preferentially grazed higher quality, non-toxic algal species over C. raciborskii, and released phosphorus which subsequently stimulated growth of C. raciborskii(Hong et al., 2013, 2015).
Of course, nitrogen and phosphorus have interacting effects on cyanobacteria, including C. raciborskii. Willis et al. (2015) found that highest growth rates were achieved for three Australian strains when nitrogen was added to cultures, with little or no phosphorus addition, suggesting an existing store of phosphorus remained within the cells. In contrast, no nitrogen addition, or nitrogen plus phosphorus addition typically resulted in lower growth rates. Studies have also showed that heterocyte production to allow nitrogen fixation can be limited by phosphorus availability (Kenesi et al., 2009;Burford et al., 2014).
5 CONCLUSION
In summary, our understanding of nutrient physiology in C. raciborskii has increased substantially in recent years due to a number of studies. It is clear that one strategy used by this organism for success in nutrient utilization is about flexibility in response to variable nutrient availability,rather than the ability to outcompete other species under increasing nutrient load. It should be stressed that although nutrients are an obvious driver of blooms, these must be considered in the context of other environmental factors, such as physical conditions and allelopathy (Antunes et al. 2015).
 Journal of Oceanology and Limnology2018年4期
Journal of Oceanology and Limnology2018年4期
- Journal of Oceanology and Limnology的其它文章
- Editorial Statement
- Response of Microcystis aeruginosa FACHB-905 to different nutrient ratios and changes in phosphorus chemistry*
- In fluence of light availability on the speci fic density, size and sinking loss of Anabaena flos- aquae and Scenedesmus obliquus*
- Application of first order rate kinetics to explain changes in bloom toxicity—the importance of understanding cell toxin quotas*
- Regime shift in Lake Dianchi (China) during the last 50 years*
- The effects of electrochemical oxidation on in-vivo fluorescence and toxin content in Microcystis aeruginosa culture*