
阪崎肠杆菌培养上清蛋白引起HepG2肝细胞的基因表达变化
2018-08-02 08:42由淑萍任立松笋陈泽良王
癌变·畸变·突变 2018年4期
由淑萍 任立松 马 龙 贺 笋陈泽良王 嵘 刘 涛*
(1.新疆医科大学公共卫生学院,新疆 乌鲁木齐 830011;2.新疆天康畜牧生物技术股份有限公司,新疆 乌 鲁木齐 830032;3.中国解放军69337部队,新疆 额 敏 834601;4.新疆医科大学护理学院,新疆 乌 鲁木齐 830011)
自Franklin[1]首次分离并报道以来,阪崎肠杆菌(Cronobacter sakazakii)已经多次引起重大食品安全事件,造成严重的后果[2-3]。2007年新疆地区在进出口食品中检测并分离到阪崎肠杆菌[4]。课题组在前期研究中发现阪崎肠杆菌新疆分离株经酪蛋白氨基酸酵母浸膏培养液(casamino acid yeast extract medium,CAYE)培养后,其纯化上清蛋白在一定条件下能够对乳鼠各组织器官产生损伤,尤其对肝组织损伤较为严重[5-7],为进一步探索该上清蛋白损伤肝组织的分子机制,本课题组纯化阪崎肠杆菌培养上清蛋白,将上清蛋白加入HepG2肝细胞培养液中,8 h后收集细胞并采用高通量测序法对实验组及对照组细胞基因进行检测,分析不同组中基因的差异表达、转录本水平定量分析、GO功能富集分析、KEGG通路富集分析,并对RPS18、RFC4基因进行荧光定量PCR验证,现报道如下。
1 材料与方法
1.1 菌株与细胞
阪崎肠杆菌新疆分离株IQCC10418,由中国检验科学院赵贵明老师惠赠,HepG2细胞购于武汉普诺赛生物公司。
1.2 实验仪器与试剂
CO2培养箱购于Thermo公司;核酸分析仪购于Biochrom公司;CAYE培养液购于北京陆桥公司,配制按照Quintero-villegas等[8]提供的方法进行,胎牛血清、高糖DMEM培养基、细胞培养6孔板购于Gibco公司,Trizol试剂购于Qiagen公司,实时荧光定量仪Light Cycler 2.0购于美国Roche罗氏公司,BCA蛋白质定量检测试剂盒购于Sangon Biotech公司。
1.3 方法
1.3.1 阪崎肠杆菌培养液上清蛋白纯化 采用CAYE培养基于CO2培养箱中37 ℃条件下培养阪崎肠杆菌;12 000 g、4 ℃离心CAYE培养液5 min;0.45 µm膜过滤上清,根据滤过后上清液的体积,按照1∶1比例加入饱合硫酸铵液并搅拌30 m in;12 0 00 g 、4 ℃ 再次进行离心,取沉淀;磷酸盐缓冲液(pH=7.0)悬浮,4 ℃ 透析。
1.3.2 蛋白质浓度测定 BCA蛋白质定量试剂盒检测纯化产物中蛋白质浓度,用蒸馏水稀释为含蛋白浓度为1.6 μg/mL的水溶液。
1.3.3 感染HepG2肝细胞、提取总RNA 含10%胎牛血清的DMEM(高糖)完全培养液培养HepG2细胞至长满培养瓶,细胞呈单层致密状时,常规胰蛋白酶消化传代,24 h换液,72 h再传代;实验调整HepG2细胞浓度至1×106/mL,于6孔培养板中分别加入1 000 μL蒸馏水及1 000 μL纯化产物稀释液(终浓度为1.6 μg/mL),建立对照组与实验组,每组3个复孔,8 h后,加胰酶回收;采用Trizol试剂盒提取总RNA;琼脂糖凝胶电泳分析样品RNA完整性及是否存在DNA污染;并检测RNA纯度[D(260)/D (280)比值]。
1.3.4 cDNA文库构建及高通量测序 用M-MLV逆转录酶和随机引物合成第一链cDNA,用DNA聚合酶合成第二链cDNA,送Novogene公司illumina芯片测序平台进行高通量测序,采用GoMiner数据库进行Gene Ontology(GO)富集分析,采用KEGG数据库进行Pathway分 析。
1.3.5 差异表达基因分析 采用DESeq2软件统计比较实验组和对照组的cDNA文库,RNA-seq测序技术进行差异显著性分析,本实验中两组间转录本的log2(Fold Change)绝对值大于1作为差异显著性标准。
1.3.6 实时荧光定量PCR 由于高通量测试结果信息量巨大,只能提供初步筛选结果,不能十分准确分析单个基因的状态及表达情况。因此进行进一步实时荧光定量PCR(quantitative real-time PCR,qPCR)验证十分必要。本次我们将选取log2(Fold Change)值为-1.10 且与核糖体密切相关、调节因子发挥重要作用的RPS18基因(ribosomal protein S18)和log2(Fold Change)值为2.13且与调控DNA复制与修复的功能分子RFC (replication factor C)亚基4进行验证。β-actin为内参基因。RPS18基因(引物序列5´-3´):P1,tggatcactcggataaatgcggtaact;P2, tggatcactcggataaatgcggtaact(GenBank登 录 号AY75781.1)。 RFC4基 因 : P1, ggaactggaaaaacatcca ctattttg; P2, ctgagcagctgaggtcatagaatctgc(GenBank登 录号CR536561.1)。反应体系共20 µL,Platinum®SYBR®Green qPCR SuperMix-UDG 10 µL,上游引物(10 µmol/L)0.4 µL,下游引物(10 µmol/L) 0.4 µL,模板 2 µL,纯水7.2 µL。反应参数为:95 ℃、15 s,95 ℃、10 s,55℃、10 s,50个循环。
2 结果
2.1 总RNA提取结果
琼脂糖电泳结果表明,提取的RNA无DNA污染。经检测各组的D(260)/D(280)的值均在1.8~2.0之间,表明提取的总RNA纯度符合实验要求。
2.2 差异表达基因
经高通量测序及基因差异表达结果分析表明,与对照组比较,共有5 120个差异表达基因,其中2 155个上调,2 965个下调。长链非编码RNA(lncRNA)为226个,占差异表达基因总数的4.41%,说明差异表达基因中非编码序列较少。表1为前5位表达上调的基因,表2为前5位表达下调的基因。
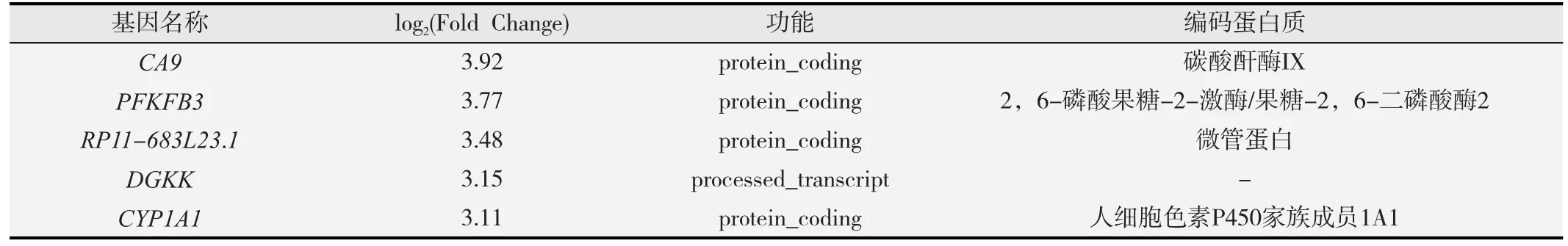
表1 前5位表达上调的基因
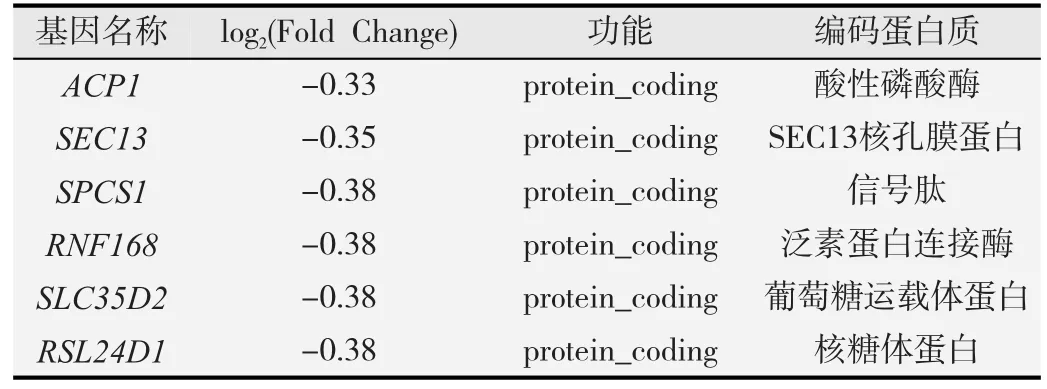
表2 前5位表达下调的基因
2.3 差异表达基因涉及到的信号通路
GO和KEGG pathway分析表明,经上清蛋白作用后,基因差异表达涉及到219个信号通路,GO富集分析显示,差异基因主要参与核分裂、有丝分裂、复制等多个生物学过程(见图1)。Pathway分析显示,具有显著富集的信号通路有5个,差异基因主要参与DNA的复制、蛋白转运、精氨酸和脯氨酸代谢等(见表3)。

图1 GO富集分析结果
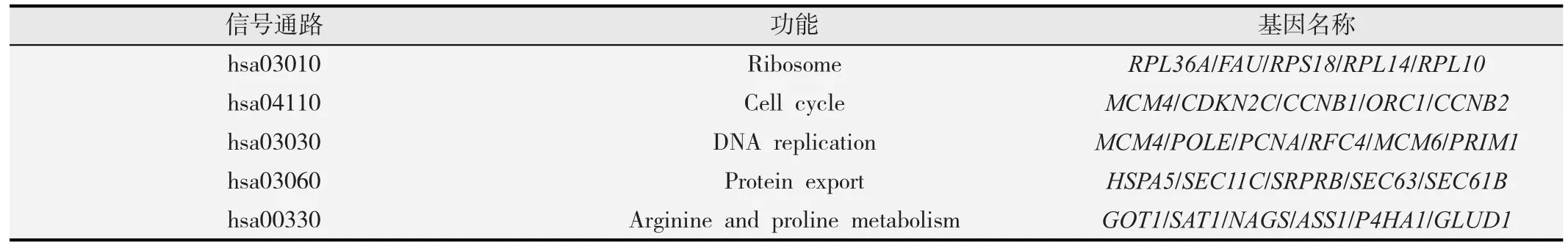
表3 基因差异表达参与的信号通路
2.4 qPCR验证结果
qPCR结果见图2和图3,可见选取的RPS18、RFC4 mRNA检测结果与高通量测序的结果相同。与对照组比较,实验组RPS18 mRNA表达呈现明显下降趋势,RFC4 mRNA表达则显著上升,差异具有统计学意义(P<0.05)。
3 讨论
目前,国内外对于阪崎肠杆菌的污染途径、致病因子、致病机理等报道非常有限。2003年,Pagotto[9]首次描述了某种类肠毒素化合物可能是阪崎肠杆菌产生的一种毒力因子。阪崎肠杆菌在培养过程中产生的肠毒素是其致病的重要因素,侵入脑微血管上皮细胞后,其外膜蛋白A和外膜蛋白X能够穿过血脑屏障,并存在于脑巨噬细胞中影响细胞因子的分泌,导致严重的大脑疾病,如新生儿脑膜炎、脑梗塞、脑囊肿等,是阪崎肠杆菌的重要致病因子[10-13]。本研究室在对阪崎肠杆菌新疆株的研究过程中发现,编号为IQCC 10418号菌株在经CAYE培养后,其培养上清中产生某种活性蛋白[14]能够对机体产生损伤,推测可能与阪崎肠杆菌致病有关。
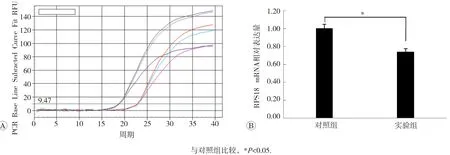
图2 两组HepG2细胞中RPS18 mRNA检测结果分析
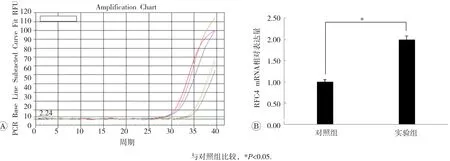
图3 两组HepG2细胞中RFC4 mRNA检测结果分析
本研究发现阪崎肠杆菌培养上清蛋白侵袭HepG2肝细胞,通过提取总RNA、反转录、高通量测序,异常表达基因有5 120个,包括2 155个表达上调基因,2 965个表达下调基因。CA9基因表达量较高,CA9基因编码碳酸酐酶蛋白,是锌金属酶家族中一员,与机体的钙代谢、酸平衡等有关;差异表达下调明显的ACP1基因,编码Acid phosphatase 1蛋白,该蛋白属于磷酸酪氨酸蛋白家族成员之一,与酪氨酸代谢相关。还有部分差异表达基因如CALR基因、TMEM 基因等编码的蛋白涉及细胞的一些重要功能,如信号转导、钙代谢等功能[15]。
在差异基因的GO富集和pathway分析[16-18]中还发现,差异表达基因参与了219个信号通路,阪崎肠杆菌侵袭HepG2肝细胞后,其差异基因的表达多为染色体、着丝粒、核糖体等变化,核糖体可从转录和翻译两个水平上调节相关基因表达,从而调控微生物的次级代谢,同时,核糖体结构改变,其代谢产物合成的调控系统也会出现变化,诱导真核细胞结构变化及有丝分裂[19-20];在差异表达基因的信号通路中,同样涉及DNA的复制、蛋白转运、精氨酸和脯氨酸代谢等多种通路,其中参与差异表达基因最多为Ribosome(hsa03010),与蛋白的翻译、转运等功能相关,参与DNA的修复,调控细胞增殖、凋亡和分化。
进一步进行结果验证表明,RPS18是由RPS18基因编码的一种蛋白,参与组成核糖体的40S小亚基,通过与起始因子以及其他核糖体蛋白相互作用,在翻译起始过程中作为一种调节因子发挥重要的作用。此外,RPS18核糖体蛋白是一种公认的Ca2+/钙调蛋白激活的蛋白激酶II底物。研究表明,RPS18参与了阪崎肠杆菌培养上清蛋白引起的HepG2肝细胞的病变,能够调节HepG2肝细胞的毒力和致病力,参与了信号转导过程。复制因子C(replication Factor C,RFC)是与DNA复制和修复过程密切相关的蛋白,RFC在ATP存在的情况下,能将PCNA和DNA聚合酶δ装配到附有引物的模板上,从而有效的延伸DNA复制链;而RFC4基因对抗DNA复制损伤修复能力具有重要的调控作用,如RFC4等基因下调可减缓肝癌细胞的增殖[21]。研究表明,RFC4基因上调,参与了DNA复制的过程,导致了阪崎肠杆菌培养上清蛋白引起的HepG2肝细胞病变的发生。
综上,从阪崎肠杆菌培养上清中分离纯化的蛋白质,可产生HepG2肝细胞病变,推测该蛋白质的生物活性可能与RPS18、RFC4基因等差异表达,引起的核糖体、染色体、着丝粒等的变化相关,参与DNA的复制与修复、蛋白转运、精氨酸和脯氨酸代谢等多条信号通路。
猜你喜欢
内蒙古民族大学学报(自然科学版)(2022年2期)2022-11-22
生物化学与生物物理进展(2022年9期)2022-09-22
科学导报(2022年29期)2022-05-26
卫星应用(2022年3期)2022-05-23
中国典型病例大全(2022年11期)2022-05-13
中国典型病例大全(2022年11期)2022-05-13
中国典型病例大全(2022年13期)2022-05-10
昆明医科大学学报(2022年2期)2022-03-29
昆明医科大学学报(2022年1期)2022-02-28
昆明医科大学学报(2021年3期)2021-07-22
