
6个新疆地方梨品种S基因型鉴定
2018-08-01 07:53张校立艾沙江买买提徐叶挺王继勋
江苏农业科学 2018年13期
张校立, 艾沙江·买买提, 徐叶挺, 许 娟, 邓 莉, 王继勋
(新疆农业科学院园艺作物研究所/农业部新疆地区果树科学观测试验站,新疆乌鲁木齐 830091)
新疆地方梨品种属于新疆梨系统。新疆梨系统是梨五大栽培系统之一,代表品种是著名的库尔勒香梨(PyrusbretschneideriRehd)。库尔勒香梨维吾尔语名乃西米提或乃西普提,蒙古语名为开登木,原产新疆库尔勒地区,在新疆已有1 400多年的栽培历史,其果实香味浓郁、皮薄肉脆、清甜爽口、细嫩多汁、耐贮藏性强,是新疆名特优果品之一。截至2016年,库尔勒香梨种植面积达7.39万hm2,产量达128.04万t[1],香梨产业已成为库尔勒地区的主导产业。库尔勒香梨自交不亲和,S基因型为S22S28[2],自交结实率低,需要配置授粉品种,但不同的授粉品种会对香梨果实产生显著的花粉直感效应,突出表现为不同花粉授粉后果实萼片脱落或宿存,萼片脱落后的幼果发育成为“母梨”,萼片宿存的幼果发育成为“公梨”,二者不但外在品质果形上差异很大,而且内在品质和口感上也差异明显,市场价格差异非常显著。徐胜利等研究表明,选择新疆梨作为授粉品种能够明显改善香梨果形和果实品质[3]。鉴定新疆地方梨品种S-等位基因对库尔勒香梨合理授粉品种的选配具有重要的理论意义。梨自交不亲和性基因的研究始于20世纪50年代,国外最早研究的是Kikuchi、Machida等日本学者,采用广泛的田间授粉杂交试验,从日本梨中鉴定了7个S等位基因,命名为S1、S2、S3、S4、S5、S6、S7[4-6]。国内最早研究的是乌云塔娜,2003年,从中国白梨中分离鉴定了7个新的S基因,分别记为S17、S19、S20、S21、S22、S26、S27等位基因[7]。谭晓风等分别从白梨、沙梨及秋子梨等亚洲梨种群分离鉴定了34个S等位基因[8]。从2003年开始至今采用PCR-RFLP、DNA测序、杂交检测等方法开展了中国梨自交不亲和性的研究,发现了包括S1~S10基因在内的42个S基因,从中国梨中克隆了S12、S13、S15、S17~S22、S25~S30、S32、S34、S35、S38~S40、S42、S46等S基因全长cDNA序列,确定了近500个梨品种的S基因型。近500个品种中大部分都是白梨系统、秋子梨系统、沙梨系统的品种,新建梨系统的品种非常少,目前,只有陈慧等鉴定了黄句句(S22S34)、伊犁红句句(S22S28)、早熟句句(S17S19)、乃希木特阿木提(S19S28)、沙01号(S22S28)、斯尔克甫梨(S22S28)、墨梨(S26Sb)、贵德长把(S19Sb)等8个新疆梨系统品种S-基因型[9]。新疆梨地方品种是我国特有的梨树资源,新疆地方梨品种S基因型的鉴定研究比较少,大部分新疆地方梨品种的S基因型还未鉴定,我们研究的6个新疆地方梨品种的基因型目前尚未有人报道。我们利用DNA测序技术鉴定了6个新疆地方梨品种的S基因型。6个品种S基因型的鉴定,以期丰富我国梨品种S基因型信息;为库尔勒香梨筛选最佳的授粉梨品种提供理论基础,并可为库尔勒香梨的丰产栽培技术提供科学依据,也为香梨品种的遗传改良奠定基础。
1 材料与方法
1.1 材料
供试品种为褐色句句梨、霍城冬黄梨、八月梨、库尔勒黄酸梨、奎克阿木特2号、也历克阿木特。品种均来自新疆农业科学院轮台国家果树资源圃。于2017年4月上旬采集各品种幼嫩叶片置于液氮中,带回实验室后于-80 ℃冷冻保存。
1.2 试剂
ExTaq、pMD18-T Vector、DNA回收试剂盒,购自北京全式金生物技术(TransGen Biotech)有限公司;DNA提取试剂盒,购于天根科技生化有限公司;其他药品均购自上海生工生物工程技术服务有限公司。
1.3 基因组DNA提取
参照植物基因组DNA提取试剂盒说明书(柱式植物DNAout2.0,天根科技生化有限公司)提取梨基因组DNA,在1%琼脂糖凝胶、1×TAE缓冲液、3~5 V/cm下电泳,电泳结果用上海复日FR-980凝胶图成像系统照相,检测DNA样品。
1.4 PCR扩增及检测
根据已报道的梨的S基因相关信息,获知梨S基因的一级结构由5个保守区、1个高频可变区及1个内含子组成。高频可变区是花粉识别的部位。S基因多态性主要源于高频可变区的差异,通常根据高频可变区的差别判断不同的S基因。因此,根据S基因一级结构特点,利用S基因高变区(hypervariable region,HV)的保守序列设计合成引物。
正向引物PF:5′-TTTACGCAGCAATATCAG-3′;
反向引物PR:5′-AC(A/G)TTCGGCCAAATAATT-3′。
在江南等扩增反应体系及反应过程[10]的基础上略作调整,使用ExTaq酶保证扩增结果的可靠性,扩增产物在1.4%琼脂糖凝胶上于100 V电泳30~45 min,上海复日FR-980凝胶图像系统分析拍照并记录分析。
1.5 PCR产物回收、克隆、测序
利用凝胶回收试剂盒回收目的DNA片段,与PEASY-T5载体16 ℃条件下过夜连接,连接产物转化至感受态大肠杆菌Trans1-T1,涂于SOC/Amp50固体培养基上37 ℃过夜。挑选阳性克隆,菌液PCR筛选带有目的片段长度的克隆。每个S基因片段挑选3个阳性克隆,送上海生工生物工程技术服务有限公司测序。测序结果利用Blast与GenBank中已知的S-RNase基因序列比较,确定其S基因型。
2 结果与分析
2.1 S基因特异性扩增
采用S等位基因特异性引物PF和PR对6个梨品种基因组DNA进行PCR扩增,琼脂糖电泳检测结果见图1。

从图1可以看出,每个梨品种扩增都得到2条S基因特异片段,大约在300~1 500 bp。褐色句句梨、霍城冬黄梨、八月梨、奎克阿木特2号、也历克阿木特等5个品种都扩增出明显的2条带;库尔勒黄酸梨扩增出来的2条带不是太明显,说明这个品种的S等位基因片段大小相差很小。
2.2 梨品种PCR扩增产物的聚丙烯酰胺凝胶电泳分离
将PCR扩增产物进一步进行聚丙烯酰胺凝胶电泳分离,PCR产物在聚丙烯酰胺凝胶上电泳随迁移率的不同,表现为不同位置的条带。从图2可以看出,6个品种在聚丙烯酰胺凝胶上均呈现2条明显的条带,褐色句句梨、霍城冬黄梨、八月梨、奎克阿木特2号4个品种的2条带都在300~500 bp之间;褐色句句梨、霍城冬黄梨的1条约450~500 bp 之间的扩增条带在聚丙烯酰胺凝胶上的位置相同,它们的S基因的碱基序列是否相同有待进一步分析;也历克阿木特的1条带在1000~1 500 bp内,另1条带在600~1 000 bp 内。
2.3 梨品种S基因型的鉴定
聚丙烯酰胺凝胶电泳后回收目的条带,将克隆后的阳性克隆送上海生工生物工程技术服务有限公司测序,将12条测定的结果序列使用生物信息学软件进行Blast搜索、同源性比较,根据分析GenBank内S基因的碱基序列及同源性,确定6个新疆地方梨品种的S基因型。从表1可以看出,6个品种中S基因的核苷酸序列与GenBank中已登录的梨S基因有极高的同源相似性,褐色句句梨的S基因与S28、S4基因的相似性达98%、99%;霍城冬黄梨的S基因与S28、S4基因的相似性达100%、99%;八月梨S基因与S28、S4基因的相似性达99%、100%;库尔勒黄酸梨的S基因与S22、S35基因的相似性达100%、100%;奎克阿木特2号的S基因与S4、S16基因的相似性达100%、100%;也历克阿木特S基因与S18、S12基因的相似性达94%、100%。

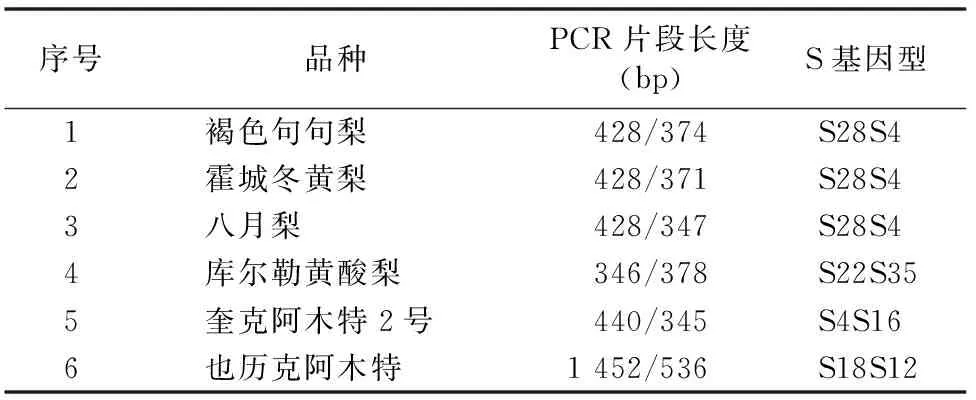
表1 6个新疆地方梨品种的S基因型
注:系统为新疆梨;引物为PF/FR。
3 讨论与结论
本研究应用PCR技术和对S基因特异性扩增条带的测序,通过DNA序列分析确定了6个新疆地方梨品种的S基因型。鉴定结果为:褐色句句梨S28S4、霍城冬黄梨S28S4、八月梨S28S4、库尔勒黄酸梨S22S35、奎克阿木特2号S4S16、也历克阿木特S18S12。同为新疆地方梨品种,它们之间既有相同之处,也有不同之处,如褐色句句梨、霍城冬黄梨、八月梨等3个品种具有相同的基因型S28S4,这3个品种间不能相互作授粉品种,因为它们具有相同的基因型,如果相互间作授粉品种,就会出现不结果或结果率极低;库尔勒黄酸梨、奎克阿木特2号、也历克阿木特3个品种的基因型不相同,这3个品种间可以相互作授粉品种。褐色句句梨、霍城冬黄梨、八月梨3个品种具有相同的基因型,并且都具有库尔勒香梨的S28,库尔勒黄酸梨具有库尔勒香梨的S22基因,表明褐色句句梨、霍城冬黄梨、八月梨、库尔勒黄酸梨与库尔勒香梨在起源上相关性比较大,奎克阿木特2号、也历克阿木特与库尔勒香梨在起源上相关性比较小。鉴定出的6个新疆地方梨品种分别含有S4、S12、S16、S18、S22、S28、S35等S基因,这些基因分别属于白梨系统、沙梨系统和西洋梨系统的S基因,初步推论新疆地方梨品种可能是白梨系统、沙梨系统和西洋梨系统的杂种,杂种后代又经过长期的自然杂交,互相演化而来成现在的品种。在果树育种学上,沈德绪等认为,新疆梨可能是西洋梨与白梨的杂交种[11]。新疆地方梨品种具体的起源与分类还需对其更多品种的S-基因进行鉴定,并需要进一步的分析与研究。
6个新疆地方梨品种基因型的鉴定,增加了我国6个新疆梨品种的S基因型信息,可为库尔勒香梨授粉品种的筛选新疆地方梨品种提供理论依据。褐色句句梨、霍城冬黄梨、八月梨3个品种具有库尔勒香梨的S28基因,库尔勒黄酸梨具有库尔勒香梨的S22基因,表明褐色句句梨、霍城冬黄梨、八月梨和库尔勒黄酸梨不太适宜作库尔勒香梨的授粉品种;奎克阿木特2号与也历克阿木特可以作为库尔勒香梨的授粉品种,但其授粉后对库尔勒香梨品质的影响,还需开展进一步研究。
猜你喜欢
今日农业(2022年16期)2022-09-22
铁道通信信号(2019年2期)2019-03-26
烟台果树(2019年1期)2019-01-28
中国三峡(2017年4期)2017-06-06
中南大学学报(自然科学版)(2016年2期)2017-01-19
系统工程与电子技术(2016年2期)2016-04-16
中国光学(2015年1期)2015-06-06
新疆农垦科技(2014年6期)2014-02-28
海岸工程(2014年4期)2014-02-27
电视技术(2010年1期)2010-08-10
