
抗扎伊尔型埃博拉病毒单克隆抗体的制备和鉴定
2018-07-31 05:04:14,,,,,,,
中国人兽共患病学报 2018年7期
, ,, ,, ,,
埃博拉病毒病(Ebola virus disease)是由埃博拉病毒(Ebola virus,EBOV)感染所致的一种急性传染病,病死率可高达90%[1]。EBOV自1976年在前扎伊尔的一个村庄被发现以来,已经引起多次暴发流行[1]。2013年底,埃博拉疫情在几内亚开始流行,并很快蔓延到利比里亚、塞拉利昂和尼日利亚等西非国家[2],2014年8月8日,世界卫生组织(WHO)宣布,西非暴发的埃博拉疫情是国际关注的突发公共卫生事件。
埃博拉病毒属于丝状病毒科埃博拉病毒属[3],埃博拉病毒属包括扎伊尔埃博拉病毒(Zaire ebolavirus,ZEBOV)、苏丹埃博拉病毒(Sudan ebolavirus,SEBOV)、雷斯顿埃博拉病毒(Reston ebolavirus,REBOV)、塔伊森林埃博拉病毒(Tai Forest ebolavirus,TAFV)和本迪布焦埃博拉病毒(Bundibugyoe ebolavirus,BEBOV)等6种病毒[4-5],其中本迪布焦型、扎伊尔型和苏丹型曾造成非洲的埃博拉暴发流行。埃博拉病毒基因组是线性、单股负链的RNA,大小约19 kb,相对分子质量为4.2×106Da[6-7]。埃博拉病毒RNA基因组可编码7种病毒蛋白,包括 VP30、VP35、 RNA依赖性RNA聚合酶、NP、VP40、糖蛋白、VP24[8-10]。与病毒结构蛋白(VP30, VP35, RNA-dependent RNA polymerase)和膜蛋白(VP40、glycoprotein and VP24)相比,NP蛋白在埃博拉病毒中丰度高,检测灵敏度强,是疾病诊断与监测中的靶标分子。由于NP靶蛋白C末端110个氨基酸片段具有很强的抗原性,因此可成为制备单克隆抗体的候选靶基因[11]。
2014-2016年西非的埃博拉出血热疫情主要是由扎伊尔型埃博拉病毒引起的。由于当地医疗条件滞后,不能及时、有效地进行病毒感染的诊断,对患者处理不当,导致疫情广泛传播[12]。随着贸易全球化、气候变暖、人类活动范围的扩大等,增加了自然或人为感染EBOV的概率,尽管我国还未有埃博拉病毒输入性感染的病例,但开展特异性好和安全的现场快速诊断技术研究对埃博拉病毒病的早期诊断和预防尤为重要。由于埃博拉病毒是危害等级为一类病原微生物,国内没有原发或输入病例,且暂无可分离培养埃博拉病毒的生物安全四级实验室,很难得到流行毒株。因此本研究依据文献合成扎伊尔型埃博拉病毒核蛋白多肽免疫动物,进行细胞融合并筛选杂交瘤细胞,制备单克隆抗体,构建4种埃博拉病亚型真核表达载体(pFLAG-CMV-Zaire-NP, pFLAG-CMV-Reston-NP, pFLAG-CMV-Tai Forest-NP、pFLAG-CMV-Sudan-NP)转染HEK293T细胞表达相应重组核蛋白,并以重组核蛋白为抗原对制备的抗扎伊尔型病毒核蛋白的单克隆抗体进行特异性鉴定,为埃博拉病毒病的现场快速诊断方法的建立奠定基础。
1 材料与方法
1.1材料 大肠埃希菌DH5α、HEK293T细胞均为中国人民解放军军事医学研究院未知病原分析室保存;pFLAG-CMV-Zaire-NP、pFLAG-CMV-Reston-NP、pFLAG-CMV-Tai Forest-NP、pFLAG-CMV-Sudan-NP质粒由生工生物工程有限公司依据我们提供的核酸序列合成;质粒提取试剂盒购自QIAGEN公司;DMEM购自北京康彻赛尔生物公司;胎牛血清购自CLESON公司;X-tremeGENE HP DNA Transfection Reagent 购自Roche公司;30%丙烯酰胺购自北京普利莱公司;抗Flag标签小鼠单抗购自北京锐抗生物公司;IRDye 800CW 羊抗鼠IgG购自LI-COR公司;缓冲盐等均为国产分析纯试剂。
1.2扎伊尔型埃博拉病毒NP优势抗原表位预测和抗原多肽合成 通过生物信息学软件DNAstar Protean预测埃博拉病毒NP氨基酸序列(ZEBOV-NP)的二级结构、亲疏水性及抗原性,并用BLAST进行同源性分析,筛选优势抗原表位,进而送上海生物技术公司人工合成多肽免疫原。
1.3 扎伊尔型埃博拉病毒单克隆抗体的制备
1.3.1实验动物的免疫 将人工合成的扎伊尔型NP多肽免疫原与弗氏完全佐剂乳化后,皮下多点注射6~8周龄BALB/c小鼠。每隔2周免疫1次,加强免疫3次。融合前3 d经腹腔注射加强免疫一次。
1.3.2细胞融合 用正常小鼠的腹腔巨噬细胞作为饲养细胞,取免疫小鼠的脾细胞与SP2/0细胞,在聚乙二醇(polyethylene glycol, PEG)的作用下进行细胞融合。融合细胞终止后离心并弃掉上清,加入预温的选择培养基HAT (1%谷氨酰胺+1%氯霉素和卡那霉素+2% HAT+20%血清),继续培养,约7~10 d,当有融合细胞出现时,进行筛选克隆。
1.3.3杂交瘤细胞的筛选 将融合后的细胞铺于96孔培养板中,培养7 d后,倒置相差显微镜下可观察到细胞较大,浑圆透亮,融合细胞集落呈葡萄串状,即杂交瘤细胞。未融合细胞逐渐死亡,裂解。吸取有杂交瘤细胞生长的细胞孔的培养上清,间接ELISA检测抗体分泌。将阳性克隆孔用有限稀释法进行培养。经过细胞单克隆株的筛选,共得到能分泌抗扎伊尔型核蛋白单克隆抗体的杂交瘤细胞12株,编号分别为ZNP1F12、ZNP2G4、ZNP3B3、ZNP3F10、ZNP5E3、ZNP6E1、ZNP12C3、ZNP4F2、ZNP14E2、ZNP15F11、ZNP9G8、ZNP7D5。
1.3.4小鼠腹水的制备 取8~10周 BALB/c小鼠腹腔注射灭菌液体石蜡,7 d后每只小鼠腹腔注射1×106个杂交瘤细胞,待注射后7~10 d,小鼠腹部明显隆起时收集腹水,离心收集上清,传统正辛酸-饱和硫酸铵法提取腹水单抗[13-14],紫外分光光度法测量蛋白浓度。
1.3.5腹水效价测定与单克隆抗体分型 采用间接ELISA方法检测腹水效价。将含单克隆抗体的腹水上清从1∶1 600开始用PBS连续倍比稀释(按照1∶2),加到包被有埃博拉病毒NP抗原多肽的KLH的板上,每个样品平行做两份,PBS为阴性对照。加入终止液,酶标仪测定OD450的吸光值,阳性反应的最大稀释度为所测样品的效价。采用小鼠单抗快速分型试剂盒(Pierce Rapid Isotyping Kits-Mouse)对单克隆抗体亚型进行鉴定。
1.4抗扎伊尔型埃博拉病毒单克隆抗体的特异性鉴定
1.4.1埃博拉病毒真核表达载体的构建 根据埃博拉病毒四型NP基因序列,用OVERLAP方法形成模板DNA,再用PCR方法得到双链DNA,亚克隆到pFlag-CMV-2载体,构建4种埃博拉病毒高效表达的重组质粒(pFLAG-CMV-Zaire-NP、pFLAG-CMV-Reston-NP、pFLAG-CMV-Tai Forest-NP、pFLAG-CMV-Sudan-NP),将4种质粒转化DH5α菌株,并用含Amp的平皿筛选重组质粒,重组质粒测序正确后,用X-tremeGENE HP DNA Transfection Reagent高效真核转染试剂瞬时转染到HEK293T细胞,培养24~48 h后收集细胞。
1.4.2埃博拉病毒真核表达蛋白的鉴定 提取4种转染重组质粒的HEK293T细胞总蛋白,并以未转染重组质粒的HEK293T细胞作为阴性对照,100 ℃加热5 min后离心收集上清液,并用BCA微量蛋白测定试剂盒测定蛋白浓度,取10 μg细胞的蛋白样品经SDS-PAGE电泳分离后,电转移至PVDF膜,经含5%的脱脂牛奶TBST溶解封闭过夜后,分别与加入一抗(鼠源性Flag单克隆抗体)和二抗(羊抗鼠IgG)孵育后,用LI-COR Odyssey 红外荧光扫描成像系统扫描条带后进行图像分析。
1.4.3抗体特异性的鉴定 用4种埃博拉病毒真核表达的重组蛋白作为抗原,1.3.4中小鼠腹水单抗为一抗,用免疫印迹法鉴定单抗的特异性,并筛选出能够分泌高特异性抗体的杂交瘤细胞株。
2 结 果
2.1根据扎伊尔型埃博拉病毒核蛋白抗原表位预测结果设计抗原多肽 通过DNAStar Protean软件分析ZEBOV NP氨基酸序列N或C末端抗原峰,表明EBOV-NP最主要的抗原峰集中在C端的420~739 aa区间,呈连续高峰,其N端仅有少量的抗原高峰。通过BLAST对所选的氨基酸序列进行同源性比较,选取特异性较好的氨基酸肽段。根据生物信息软件预测分析结果设计了ZEBOV NP特异性抗原多肽(aa611-630: YRDHSEKKELPQDEQQDQDH)。
2.2腹水效价测定 通过间接ELISA方法检测所制备腹水效价(见表1),表明杂交瘤细胞株腹水抗体效价分别介于1∶104~1∶105。使用小鼠单抗分型试剂盒对扎伊尔型细胞株分泌的抗体型别进行检测,如表1:其中有1株IgG2a型(ZNP9G8),2株IgM型(ZNP3B3、ZNP4F2),2株IgG2b型(ZNP14E2、ZNP3F10),其余7株为IgG1型。
表1 埃博拉病毒NP单克隆抗体分型及腹水效价测定
Tab.1 IsotyPes and antibody titers of aseite fiuid of Ebola virus NP-specific mAbs
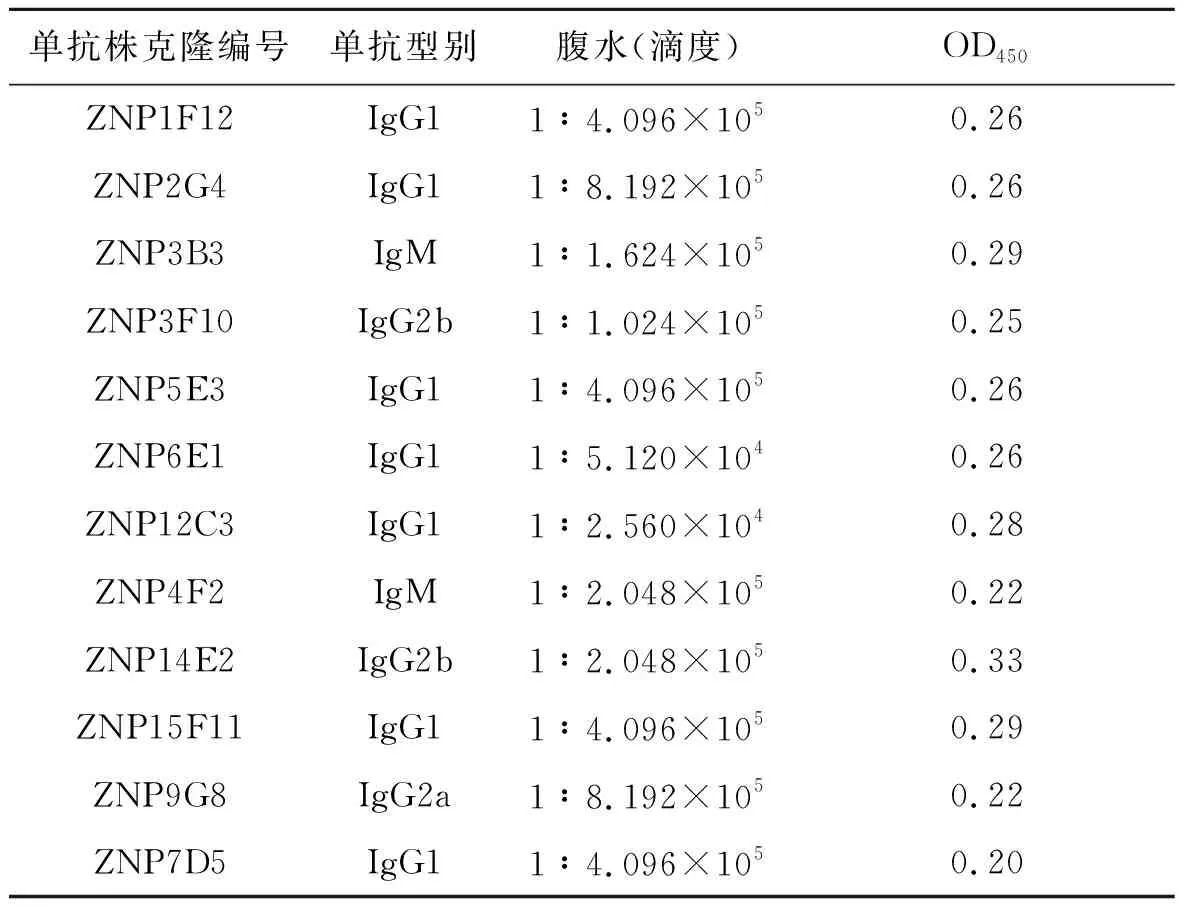
单抗株克隆编号单抗型别腹水(滴度)OD450ZNP1F12IgG11∶4.096×1050.26ZNP2G4IgG11∶8.192×1050.26ZNP3B3IgM1∶1.624×1050.29ZNP3F10IgG2b1∶1.024×1050.25ZNP5E3IgG11∶4.096×1050.26ZNP6E1IgG11∶5.120×1040.26ZNP12C3IgG11∶2.560×1040.28ZNP4F2IgM1∶2.048×1050.22ZNP14E2IgG2b1∶2.048×1050.33ZNP15F11IgG11∶4.096×1050.29ZNP9G8IgG2a1∶8.192×1050.22ZNP7D5IgG11∶4.096×1050.20
2.3埃博拉病毒4种亚型NP重组质粒的构建和真核重组表达蛋白的鉴定 4种重组表达质粒的DNA测序结果表明插入的序列完全正确,成功构建了4种埃博拉病毒NP蛋白的重组质粒。将构建成功的4种埃博拉病毒NP重组质粒瞬时转染HEK293T细胞,Western blot鉴定4种NP融合蛋白的表达。如图1:埃博拉病毒四种亚型的NP融合蛋白分子量大小介于95~120 kDa之间,均检测到一条特异性蛋白印迹条带,而未转染重组质粒的对照组无相应条带出现。说明埃博拉病毒4种亚型的NP重组蛋白真核表达成功。

M:蛋白分子量标准;1:Zaire-NP; 2:Reston-NP;3: Tai Forest-NP;4: Sudan-NP;5:对照图1 免疫印迹法检测四种埃博拉病毒真核重组蛋白的表达Fig.1 Detection of eukaryotic recombinant protein of four Ebola virus by Western blot
2.4抗扎伊尔型埃博拉病毒NP蛋白的单克隆抗体特异性的鉴定 为获得特异性较高单抗细胞株,将转染表达的Zaire-NP、Reston-NP、Tai Forest-NP、Sudan-NP蛋白进行电泳,转膜后封闭,与收集的阳性克隆细胞上清一起4 ℃过夜孵育,分别对获得的12株抗EBOV NP单克隆细胞株进行免疫印迹分析。如图2所示,有9株(ZNP1F12、ZNP2G4、ZNP3B3、ZNP5E3、ZNP6E1、ZNP12C3、ZNP4F2、ZNP9G8、ZNP7D5)与其他型别Reston-NP、Tai Forest-NP、Sudan-NP抗原另存在不同程度的交叉反应(图2),特异性不强。另外3株(ZNP3F10、ZNP14E2、ZNP15F11)单克隆细胞株只与Zaire-NP抗原产生反应,与外3种亚型的埃博拉病毒NP蛋白抗原无交叉反应,表明这3株细胞分泌的单克隆抗体特异性较高。
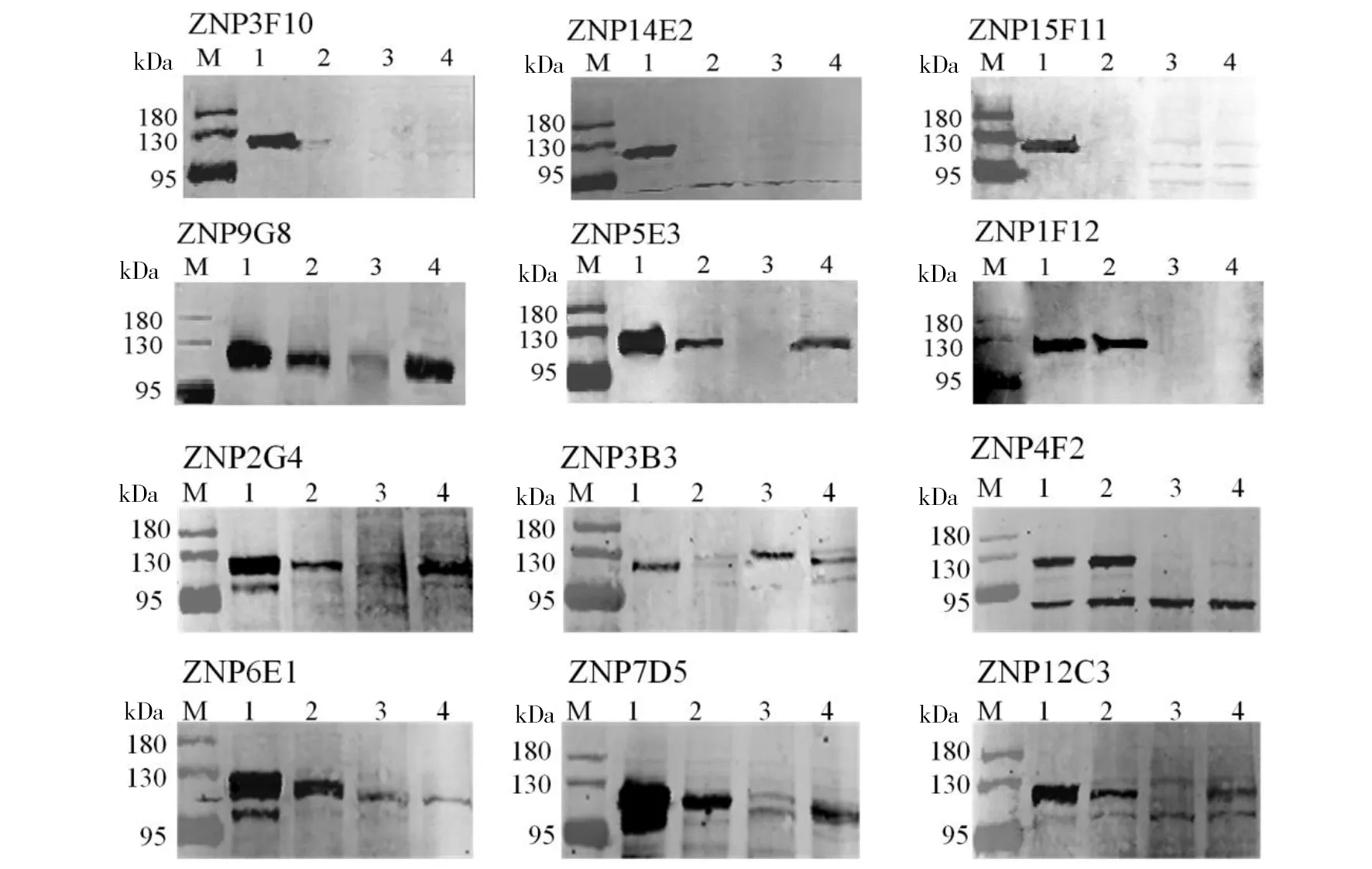
M:蛋白分子量标准;1:Zaire-NP; 2:Reston-NP;3: Tai Forest-NP;4: Sudan-NP图2 免疫印迹法检测抗扎伊尔型单克隆抗体的特异性Fig.2 Detection of the specificity of monoclonal antibodies against Zaire Ebola virus by Western blot
3 讨 论
本研究依据扎伊尔型埃博拉病毒核蛋白序列合成多肽免疫小鼠,成功制备出抗扎伊尔型埃博拉病毒单克隆抗体,并对制备单克隆抗体进行了效价测定和单抗分型。同时用4种埃博拉病毒亚型重组载体(pFLAG-CMV-Zaire-NP、pFLAG-CMV-Reston-NP、pFLAG-CMV-Tai Forest-NP、pFLAG-CMV-Sudan-NP)的真核表达蛋白为抗原对单克隆抗体的特异性和交叉反应进行鉴定,最终成功筛选出特异性高的抗扎伊尔型单克隆抗体杂交瘤细胞3株,这3株细胞分泌的抗体效价高,且都属于IgG型抗体,相对IgM型抗体较稳定,效价不易下降。
现有的埃博拉病毒的检测技术包括病毒的分离培养、IgM和IgG抗体的检测、荧光定量PCR、恒温核酸检测技术、免疫组化及免疫荧光等技术等[15]。然而,埃博拉病毒分离培养需生物安全四级实验室,分离培养时间长,并且埃博拉病毒传染性强,潜伏期短,病情进展快,病死率高,急需建立快速检测方法;而IgM和IgG抗体的检测及免疫组化技术等方法需要高效价和特异性好的单克隆抗体,因此制备出特异性高的埃博拉单克隆抗体并筛选出能分泌埃博拉单克隆抗体的杂交瘤细胞尤为重要。
目前已知的埃博拉病毒属的5个种中,扎伊尔型对人致病性最强,致死率为60%~90%;苏丹型、本迪布焦型次之,致死率为40%~60%[10];雷斯顿、塔伊森林型对黑猩猩致病性强,但对人致病性较弱,至今尚无引起疾病或死亡的报道。单克隆抗体制备过程长和工作量大,埃博拉病毒抗体特异性的鉴定当然最好是选择4种亚型的埃博拉活病毒,但埃博拉病毒属于危害等级为一类的病原微生物生物,操作需在生物安全P4级实验室完成,因此在活毒株来源有限条件下,利用基因工程技术合成重组NP蛋白是目前实验室进行埃博拉免疫学研究的主要途径。然而在实际应用中,抗体最终识别的是病毒的天然蛋白,因此对抗原的设计要求更加严格,根据多肽序列设计原则,本研究设计的ZEBOV-NP抗原肽段纯度较高,与载体蛋白偶联后免疫效果良好。而重组蛋白真核表达是获取大量目的蛋白的途径,并且哺乳动物表达系统表达的外源性蛋白的生物学活性最高,其抗原性更接近天然蛋白,因此利用真核表达的蛋白抗原代替不易得到的强毒性活病毒来鉴定抗体的特异性在抗体制备研究中应用广泛,准确可靠[16-17]。本研究成功制备效价高和特异性好的抗扎伊尔型埃博拉病毒抗体,为建立快速检测埃博拉病毒的方法奠定基础。
猜你喜欢
中国现代医药杂志(2020年10期)2020-12-14 07:20:14
现代检验医学杂志(2016年3期)2016-11-15 01:59:28
西南医科大学学报(2015年1期)2015-08-22 13:01:46
医学研究杂志(2015年6期)2015-07-01 17:41:11
医学研究杂志(2015年3期)2015-06-10 06:41:52
特产研究(2015年1期)2015-04-12 06:36:20
微生物与感染(2015年1期)2015-02-28 17:42:36
发明与创新(2015年9期)2015-02-27 10:38:44
癌变·畸变·突变(2015年3期)2015-02-27 06:15:07
中国卫生(2014年12期)2014-11-12 13:12:46
