
新疆部分生态区亚洲飞蝗地理种群形态变异研究
2018-07-30 05:15:42族米娜牙森沙力阿孜古丽阿布力孜徐光青古丽孜热曼合木提涂雄兵
新疆农业科学 2018年5期
族米娜,牙森·沙力,阿孜古丽·阿布力孜,徐光青,古丽孜热·曼合木提,涂雄兵
(1.新疆农业大学农学院/农林有害生物监测与安全防控重点实验室,乌鲁木齐 830052;2.阿勒泰地区治蝗灭鼠指挥部办公室,新疆阿勒泰 836700;3. 中国农业科学院植物保护研究所/植物病虫害生物学国家重点实验室,北京 100193)
0 引 言
【研究意义】亚洲飞蝗(Locustamigratoriamigratoria)属于直翅目Orthoptera 蝗总科Acridoidea斑翅蝗科Oedipodidae,飞蝗属LocustaLinnaeus[1]。是农业生产中危害最为严重的世界性害虫之一,历史上与水灾、旱灾并称为我国三大自然灾害[2]。主要为害禾本科与莎草科牧草为主[3]。亚洲飞蝗分布于亚洲地区西南部及欧洲地区的南部、蒙古、朝鲜、日本北部及中国的北部均有分布[4]。张学祖(1955)报道中最早提到新疆的飞蝗为迁移飞蝗即亚洲飞蝗。据历史记载亚洲飞蝗在新疆发生最早于1941年开始。20世纪50~70年代,30年间曾发生4次大的飞蝗灾害。累计发生年数达15年。1977年亚洲飞蝗扩散面积最大达到2×104hm2,农牧业生产受到最大破坏[5]。亚洲飞蝗在新疆分布范围较广,主要分布于塔城地区、伊犁地区、昌吉、玛纳斯、库尔勒等30个县市[6-7]。因昆虫的迁飞,迁徙习性,不同的地理环境是否会引起昆虫形态上的变异,自国外学者Uvarov[8]提出利用形态测量的方法测量飞蝗的变型现象。中国学者陈永林[9]也通过测量散居型飞蝗标本,对中国3个飞蝗亚种进行了比较研究。陈若篪等[10]通过测定其前翅、后足腿节、上颚口针、及产卵瓣长度,得出不同的地区褐飞虱形态上有明显的变异。郑福山等[11]通过数值分类学研究了12个不同地理种群的菱角萤叶甲数量性状的变异与地理分布之间的关系。索中毅等[12]运用几何形态学方法对国内外30个地区700头白星花金龟鞘翅和前胸背板进行了比较分析,阐明了不同地理种群的白星花金龟地理形态变异。飞蝗主要以迁飞危害为主,其一个世代内可以聚集迁飞长达 2 575 km[13]。新疆地大物博又因独特的地理环境条件,南疆地区由于天山的阻隔,塔里木盆地被周围的高山围住北冰洋的湿润空气难以越过天山等周围的山脉,降水量极低,空气异常的干燥,绿洲地带贫瘠。北疆地区由于准噶尔盆地周围的山脉比较低矮,缺口较多北冰洋与大西洋气流可以流通,空气较为湿润。富有优质的草原与牧场。因南北疆气候条件的差异,导致南疆与北疆的迥异的地理环境条件[14]。动物体型变异的原因与一系列环境因子有关[15]。又因新疆各地区环境条件不同,以期明确新疆不同地理种群亚洲飞蝗的形态是否存在地理分化,为研究亚洲飞蝗形态特征与生态适应性的关系提供理论依据。【前人研究进展】前人对亚洲飞蝗研究报道主要以其生物学特性[16-17]、取食特性[18]、有丝分裂与减数分类观察[19]、染色体核型分析[20]、发生器结构与鸣声时域[21]等方面进行了深入研究。【本研究切入点】对亚洲飞蝗形态特征与地理变异相关的研究文献较少,甚至国内尚未报道。不同的地理种群昆虫存在相应的地理变异现象,研究新疆亚洲飞蝗是否也存在地理变异情况。【拟解决的关键问题】以亚洲飞蝗主要分布的6个地理种群为材料,利用数值分类学方法,对散居型亚洲飞蝗进行形态测量,研究不同地理种群散居型亚洲飞蝗形态上是否存在地理变异。
1 材料与方法
1.1 材 料
2015~2016年7~9月分别在新疆富蕴县、托克逊县、玛纳斯县、哈巴河县实验种群、哈密市黄田庙尔沟、和硕县等县市亚洲飞蝗核心分布区域使用扫网法采集散居型成虫,哈巴河县实验种群来自于阿勒泰市科克苏湿地保护区,在阿勒泰哈巴河实验基地养虫室进行饲养至成虫。共采集散居型亚洲飞蝗成虫标本为495头。列出具体采集信息。表1
表1 亚洲飞蝗成虫标本采集信息
Table 1 Information of collection of Locusta migratoria L.

采集地点Sampling site种群Population代码Code经度(E)Longitude纬度(N)Latitude样本数(头)Sample阿勒泰地区富蕴县库尔特乡Fuyun County Kurt Township in Aletai area富蕴FY89°52'92″47°00'33″135吐鲁番地区托克逊县Toksun County, Turpan area托克逊TKX88°68'61″42°79'38″71巴州地区和硕县蝴蝶沟Heshuo County of Mongolia Autonomous Prefecture of Bayinguoleng和硕HX86°88'36″42°28'80″69阿勒泰地区哈巴河县实验种群Experimental population in Habahe County of Aletai哈巴河HBH86°41'98″48°06'67″58昌吉地区玛纳斯县平原林场The plain forest farm of Manasi County in Changji玛纳斯MNS86°21'92″44°31'73″96哈密地区黄田庙尔沟Mioer gully of Huang Tian in Hami area哈密HM93°74'86″42°80'31″66
1.2 方 法
1.2.1 测量指标
根据第四届国际蝗虫学会议(1936)所规定的测量标准,以游标卡尺(精确度:0.01 mm)测量了495头亚洲飞蝗成虫标本的形态指标。形态参数包括体长(Length, L)、前翅长度(Elytron length, E)、后足股节长度(Posterior femur length , F)、头高( Height of head,CH),头宽(Maximum width of head, C)前胸背板长(Pronotum length,P)、前胸背板宽度(Pronotum width, M)、前胸背板高度(Pronotum height,H)后足胫节长度(Tibia length, T)、颜顶角(Fastigium,A)。图1
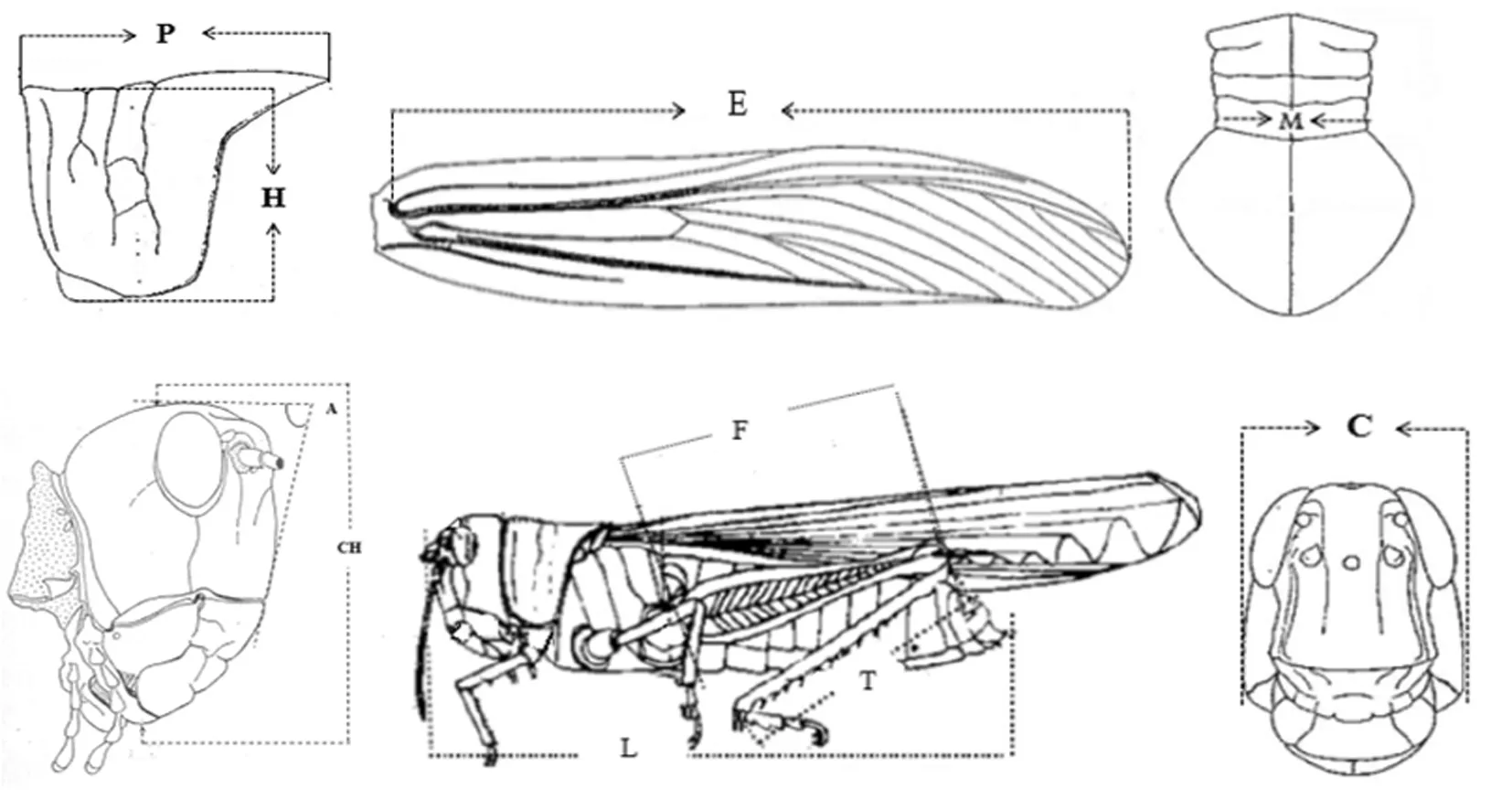
图1 10种形态指标的形态测量标准
Fig.1 Standard scheme of morphometric measurements of 10 morphometric measurements(Diesh,1953, 陈永林,1978)
1.2.2 各性状之间的相关性
将亚洲飞蝗6个地理种群共有体长、前翅长度、前胸背板长度、前胸背板高度、前胸背板宽度、后足股节长度、后足胫节长度、头宽、头高、颜顶角等10个性状测量值进行Pearson系数相关性分析。
运用Pearson相关系数分析散居型亚洲飞蝗不同性别10个形态参数之间的相关性。
1.2.3 聚类分析与主成分分析
将雌性与雄性亚洲飞蝗共有显著差异性的体长、前翅长度、前胸背板长度、前胸背板高度等四个性状平均值进行系统聚类和主成分分析。
将雌性与雄性亚洲飞蝗共有显著差异性的性状体长、前翅长度、前胸背板长度、前胸背板高度四个性状进行系统聚类和主成分分析。
1.3 数据处理
运用SAS8.0软件对亚洲飞蝗10项形态参数进行单因素方差分析及Ducan(新复极差法)多重比较,运用Pearson相关系数分析各形态参数相关性,并对每个形态参数平均值进行系统聚类分析(Hierarchi Cluster Analysis,HCA)和主成分分析(Principal Component Analysis,PCA)。
2 结果与分析
2.1 形态差异
研究表明,分布于富蕴县、托克逊县、和硕县、哈巴河县实验种群、玛纳斯县、哈密市黄田庙尔沟6个地理种群雌性亚洲飞蝗成虫之间在体长、前翅长度、后足股节长度、前胸背板长度、前胸背板宽度、前胸背板高度、后足胫节长度等性状存在极显著差异(P<0.01),而富蕴县、托克逊县、和硕县、哈巴河县实验种群、玛纳斯县地理种群在头宽、颜顶角、头高三个性状未存在显著差异(P>0.05)。同样分布于6个地理种群雄性亚洲飞蝗在体长、前翅长度、前胸背板长度、前胸背板高度、等性状之间存在极显著差异(P<0.01)。而后足股节、前胸背板宽度、后足胫节长度、等性状不同地理种群之间未存在显著性差异(P>0.05)。雌性与雄性亚洲飞蝗成虫均在体长、前翅长度、前胸背板长度、前胸背板高度等性状上存在显著性差异,在头宽与头高两个性状之间未存在显著性差异。表2

2.2 各性状相关性
研究表明,亚洲飞蝗不同地理种群雌性成虫形态参数可知,体长与前翅长度、头高之间存在极显著正相关性、相关系数为0.968。与前胸背板长度、前胸背板高度之间存在显著正相关性相关系数为0.872、0.841。相关系数均大于0.5,说明这些性状的变化对体长的大小有一定影响。前翅长度与前胸背板长度、前胸背板高度、后足股节长度、前胸背板宽度、后足胫节长度、头宽等性状之间未存在显著相关性,说明前翅长度的大小除了体长大小与头高以外其他性状无关。前胸背板长度与前胸背板高度、前胸背板宽度、后足胫节长度之间具有极显著正相关性,相关系数为0.969、0.934、0.937。与后足股节长度、头宽、头高等性状之间具有显著正相关性,相关系数为0.866、0.906。前胸背板高度与前胸背板宽度和头宽之间具有极显著正相关性,相关系数为0.937、0.949。与后足股节长度、后足胫节长度之间存在显著正相关性,相关系数为0.847、0.895、。后足股节长度与前胸背板宽度、后足胫节长度之间存在极显著正相关性相关系数为0.925、0.980。前胸背板宽度与后足胫节长度、头宽两个性状之间存在极显著正相关性,相关系数为0.944、0.931。与头高之间存在显著相关性,相关系数为0.869。表3
亚洲飞蝗雄性形态参数相关性分析表明,体长形态参数与前胸背板高度、前胸背板宽度性状之间存在极显著正相关性,相关系数为0.937、0.936。与后足胫节长度、头宽、头高等性状之间存在显著正相关性,相关系数为0.859、0.879、0.896。这说明除了前翅长度、前胸背板长度、后足股节长度以外的性状对雄性成虫体长的变化有密切的联系。前翅长度与前胸背板长度、头宽两个性状之间存在极显著正相关性,相关系数为0.929、0.973。与前胸背板高度、后足股节长度、前胸背板宽度、后足胫节长度之间存在显著正相关性,相关系数为0.902、0.876、0.880、0.880。说明除了体长、头高、颜顶角以外的性状对雄性成虫前翅长度的大小有较大的影响。前胸背板长度与前胸背板高度、前胸背板宽度、后足股节长度、后足胫节长度、头宽之间均存在极显著正相关性,相关系数为0.933、0.980、0.933、0.964、0.934。说明大多数性状对前胸背板长度变化存在很大的影响。前胸背板高度与前胸背板宽度、后足胫节长度、头宽、头高等性状之间存在极显著正相关性,相关系数为0.953、0.966、0.970、0.928。与后足股节有显著正相关性,相关系数为0.847。后足股节长度与后足胫节长度之间存在极显著正相关性,相关系数为0.984,与头宽之间存在显著相关性,相关系数为0.888。前胸背板宽度与头宽、头高之间存在极显著相关性,相关系数为0.956、0.929,与后足胫节长度之间存在显著相关性,相关系数为0.864。后足胫节长度与头宽之间存在极显著相关性,相关系数为0.924。表4
2.3 种群数量性状系统聚类
通过系统聚类将亚洲飞蝗6个地理种群形成6个分类单元(Operational taxonomic units,简称OTU),研究表明,由6个操作单元可形成2个类群。第Ⅰ类群由阿勒泰地区富蕴县库尔特乡与巴州地区和硕县蝴蝶沟、哈巴河县实验种群组成,第Ⅱ类群由吐鲁番地区托克逊县种群与昌吉地区玛纳斯县平原林场种群、哈密市黄田庙尔沟种群组成。第Ⅰ类群中巴州地区和硕县蝴蝶沟与哈巴河县实验种群聚为一小类,第Ⅱ类群中昌吉地区玛纳斯县平原林场种群和哈密市黄田庙尔沟种群聚类为一小类。图2
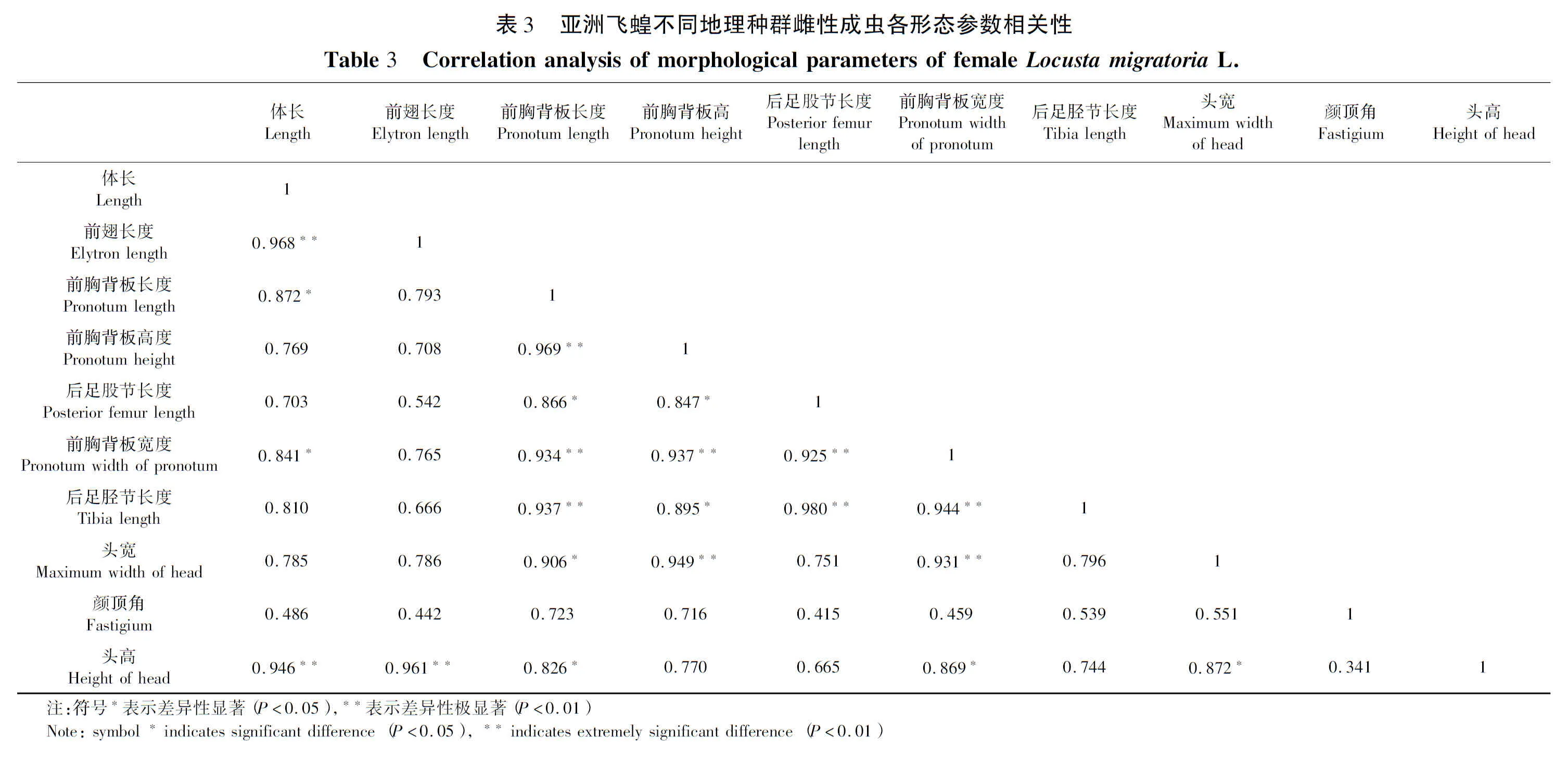

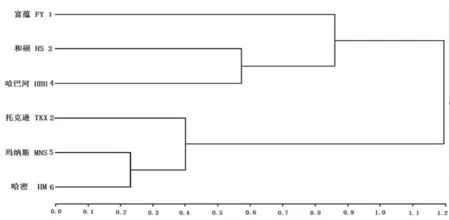
图2 亚洲飞蝗地理种群系统聚类分析树系
Fig.2 The Locusta migratoria L tree diagram of Hierarchical Cluster Analysis
2.3 种群数量性状主成分
研究表明,前三个主成分占总信息量的99%,第一个主成分Prin1=(0.365L+0.364E+0.348P+0.332H)雌性+(0.329L+0.351E+0.369P+0.368H)雄性,特征值为7.118 6,说明总变异值为88.99%是体长、前翅长度、前胸背板长度、前胸背板高度在形态构建上的综合作用。并且以上各个指标的系数相接近,因此,是其形态指标及其比值的加权平均,代表了亚洲飞蝗6个地理种群在新疆的分化程度。第二个主成分Prin2=(-0.105L-0.175E+ 0.467P+ 0.575H)雌性+(-0.606L-0.010E+ 0.064P -0.196H)雄性,特征值为0.617 0,能说明总变异的7.74%是雌性前胸背板长度、前胸背板高度与雄性前胸背板长度的作用。在雌性前胸背板长度、前胸背板高度和雄性前胸背板长度上有正系数。在雌性体长、前翅长度和雄性体长、前翅长度和前胸背板高度有负系数。其雌性前胸背板长度、前胸背板高度与体长、前翅长度和雄性体长、前翅长度、前胸背板高度是相互对比。Prin2雌性前胸背板长和雌性前胸背板高系数最大为0.467、0.575,起着显著的作用,作用系数最小的为雄性前胸背板长度,为0.064。其他参数系数为负值,因此,在Prin2中起着负作用。 第三个主成分Prin3=( -0.424L-0.401E-0.020P-0.173263H)雌性+(0.004L+0.762E+0.216P+0.046H)雄性,特征值0.194 2,总变异值为2.43%。 是雄性体长、前翅长度、前胸背板长度和前胸背板高度的作用。在雄性体长、前翅长度、前胸背板长度、前胸背板高度有正系数,在雌性体长、前翅长度、前胸背板长度、前胸背板高度有负系数。其雌性的体长、前翅长度、前胸背板长度、前胸背板高度四个参数系数值与雄性体长、前翅长度、前胸背板长、前胸背板高度是相互对比。Prin3中雄性前翅长度起着显著作用,作用系数为0.762。作用最小的是雄性体长,作用系数为0.004。雌性的参数系数都为负值,在Prin3中起负作用。
研究表明,亚洲飞蝗不同地理种群数量性状及其参数平均值运算而得到前三个主成分得分,分别以Prin1和Prin2、Prin1和Prin3、Prin2和Prin3绘制二维排序图。表5,表6
表5 亚洲飞蝗地理种群数量性状主成分参数
Table 5 Principal component parameters of Locusta migratoria L
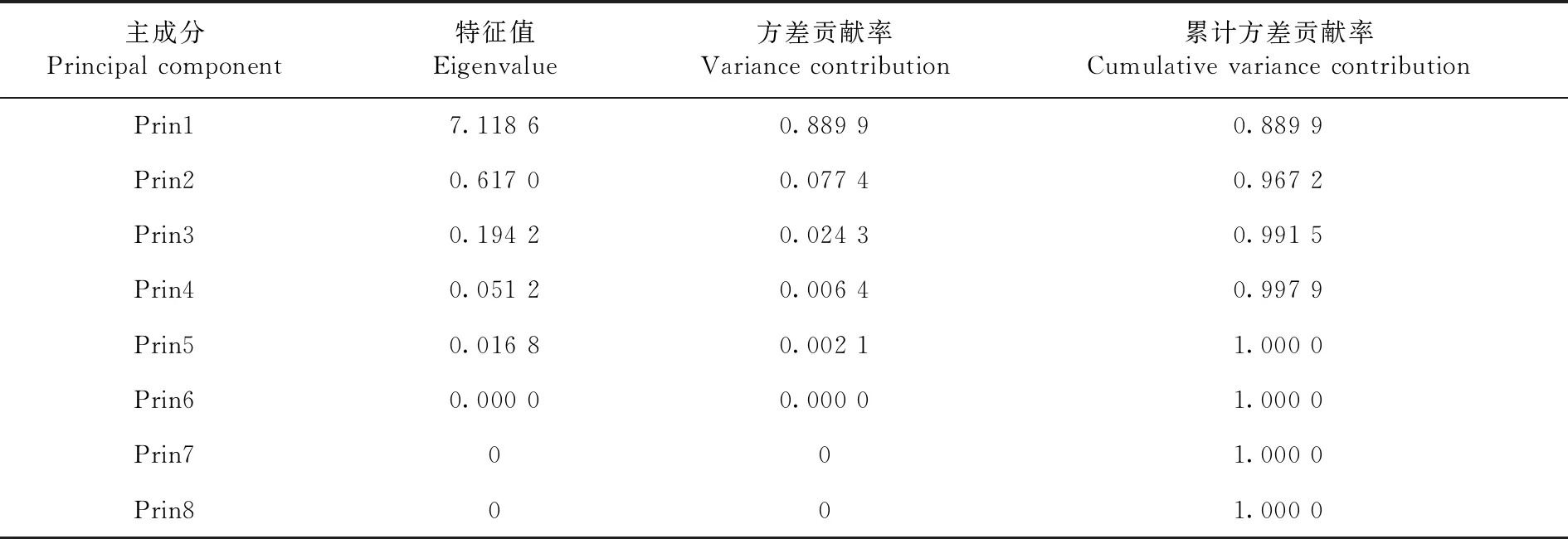
主成分Principal component特征值Eigenvalue方差贡献率Variance contribution累计方差贡献率Cumulative variance contributionPrin17.118 60.889 90.889 9Prin20.617 00.077 40.967 2Prin30.194 20.024 30.991 5Prin40.051 20.006 40.997 9Prin50.016 80.002 11.000 0Prin60.000 00.000 01.000 0Prin7001.000 0Prin8001.000 0
表6 亚洲飞蝗地理种群数量性状主成分得分
Table 6 Principal compoent of Locusta migratoria L

种群代号主成分得分Prin1Prin2Prin3富蕴FY4.014 58-0.369 810.037 48托克逊TKX-1.552 431.305 490.299 02和硕HS2.239 250.615 97-0.469 15哈巴河HBH0.183 55-0.603 490.671 48玛纳斯MNS-2.232 57-0.648 24-0.448 14哈密HM-2.652 38-0.299 92-0.090 69
研究表明,Prin1和Prin2组合而成的二维排序图中,哈密县(HM)、和玛纳斯县(MNS)地理种群形成一组,其哈密县(HM)与玛纳斯县(MNS)地理种群的形态指标和比值的综合效应接近。哈巴河县实验种群(HBH)、和富蕴县(FY)地理种群形成一组,两者虽形成一组,但其形态指标相差较大。体型较小的托克逊县(TKX)、和体型较大的和硕县(HS)地理种群单独形成一组,其中托克逊县(TKX)与 富蕴县(FY)、哈巴河县(HBH)地理种群之间的差异明显大于和硕县(HS)与 富蕴县(FY)、哈巴河县实验种群(HBH)地理种群之间的差异。和硕县(HS)与哈密市黄田庙尔沟(HM)、玛纳斯县(MNS)地理种群之间的差异明显大于托克逊县(TKX)与哈密市黄田庙尔沟(HM)玛纳斯县(MNS)地理种群之间的差异。哈巴河县实验种群(HBH)与哈密市黄田庙尔沟(HM)、玛纳斯县(MNS)之间的差异小于哈巴河县实验种群(HBH)与和硕县(HS)、富蕴县(FY)、托克逊县(TKX)之间的差异。图3
研究表明,Prin2和Prin3组合而成的二维排序图与 Prin1和Prin2组合而成的二维排序图相似。哈巴河县实验种群(HBH)、与富蕴县(FY)地理种群虽形成一组,但富蕴县(FY)与哈密市黄田庙尔沟(HM)地理种群之间的差异小于与哈巴河县实验种群(HBH)之间的差异。哈密市黄田庙尔沟(HM)、与玛纳斯县(MNS)地理种群形成一组,玛纳斯县(MNS)与托克逊县(TKX)地理种群之间的差异明显大于哈密市黄田庙尔沟(HM)地理种群与托克逊县(TKX)地理种群之间的差异。托克逊县地理种群(TKX)与和硕县(HS)地理种群单独形成一组。和硕县(HS)地理种群与哈巴河县实验种群(HBH)、玛纳斯县(MNS)、富蕴县(FY)、哈密市黄田庙尔沟(HM)地理种群之间的差异要小于托克逊县(TKX)地理种群与它们之间的差异。图4
研究表明,Prin1和Prin3组合而成的二维排序图虽也与 Prin1和Prin2、 Prin2和Prin3组合而成的二维排序图相似,但和硕县(HS)地理种群与富蕴县(FY)地理种群之间的差异要小于与哈巴河县实验种群(HBH)之间的差异,托克逊县(TKX)地理种群与玛纳斯县(MNS)地理种群、哈密市黄田庙尔沟(HM)地理种群之间的差异小于托克逊县地理种群与哈巴河县实验种群(HBH)、富蕴县(FY)地理种群、和硕县(HS)地理种群之间的差异。图5

图3 6个操作单元主成分分析二维排序(Prin1&Prin2)
Fig.3 Ordination on principal component coordinates Prin1 and Prin2

图4 6个操作单元主成分分析二位排序(Prin2&Prin3)
Fig.4 Ordination on principal component coordinates Prin1 and Prin3

图5 6个操作单元主成分分析二位排序(Prin1&Prin3)
Fig.5 Ordination on principal component coordinates Prin2 and Prin3
3 讨 论
通过形态测量方法,将新疆6个地理种群采集的495头散居型亚洲飞蝗成虫10个性状测量值进行了形态差异、及种群分化方面的研究。形态差异分析结果得出雌性亚洲飞蝗所有性状均值都大于雄性飞蝗,这与李涛等[22]研究中华稻蝗形态特征分析结果相似。其富蕴县、和硕县、哈巴河县实验种群、玛纳斯县、托克逊县、哈密市黄田庙尔沟等6个地理种群雌雄亚洲飞蝗成虫个体均在体长、前翅长度、前胸背板长度、前胸背板高度等4个性状上存在地理差异。其中雌性成虫体长、前翅长度、后足股节长度、前胸背板长度、前胸背板宽度、前胸背板高度、后足胫节长度等性状测量值存在地理差异,而雄性成虫在体长、前翅长度、前胸背板长度、前胸背板高度、颜顶角等性状上存在地理差异。说明不同地理环境条件下雌性与雄性成虫形态变异程度不一致[23]。
在前人研究中直翅目昆虫蟋蟀Teleogryllusemma和螽斯Ephippigerterrestris也存在不同地理种群雌性与雄性个体体型变异程度不一致的现象[24-25]。对亚洲飞蝗6个地理种群10个性状之间进行相关性分析,亚洲飞蝗成虫各个性状之间都存在相互促进,共同变异的现象。将雌性与雄性共有差异性的指标体长、前翅长度、前胸背板长度、前胸背板高度作为主要形态指标参数进行聚类分析与主成分分析,探明散居型亚洲飞蝗不同地理种群之间的形态变异。
前人研究中对不同地理种群烟蚜[26]、菱角萤叶甲[11]等形态特征差异聚类分析与主成分分析结果都一致,而牙森·沙力等[27]研究不同地理种群散居型西藏飞蝗数量性状系统聚类结果与主成分分析结果不一致,研究系统聚类与主成分结果也存在差异性。在聚类分析结果中分为两个类群:富蕴县地理种群、和硕县地理种群、哈巴河县实验种群聚为一个类群,其中和硕县地理种群与哈巴河县实验种群聚为一小类群,因富蕴县地理种群形态参数与和硕县与哈巴河县实验种群有较大的差异性单独为一小类群。托克逊县地理种群、玛纳斯县地理种群、哈密市黄田庙尔沟地理种群为一个类群,托克逊地理种群与哈密市黄田庙尔沟地理种群聚为一小类群。因玛纳斯县地理种群与哈密市黄田庙尔沟地理种群性状之间的相似性较接近,与托克逊县地理种群有较大的差异性,托克逊县地理种群单独为一小类群。而主成分分析结果中,Prin1&Prin2、Prin2&Prin3、Prin1&Prin3二维排序图中分布区域为大陆性暖温带荒漠气候托克逊县地理种群与暖温带大陆性干旱气候和硕县地理种群单独分布,而大陆性寒温带干旱气候富蕴县地理种群与大陆性北温带寒冷气候的哈巴河县实验种群分布在一起,属温带大陆干旱性气候的玛纳斯县与哈密市黄田庙尔沟地理种群分布在一起。相互归类的地理种群虽有相似的气候特征,但形态特征也具有一定的差异性。也与牙森·沙力[27]等研究散居性西藏飞蝗9个地理种群形态特征数量分析结果中得出的结论相似。
以近年来亚洲飞蝗主要分布危害的部分主要生态区为主采集散居型标本,未能全面采集亚洲飞蝗在新疆所分布的所有区域内的虫源,致使全面反映新疆各个生态区亚洲飞蝗形态变异的研究存在局限性。塔城、伊犁、喀什、阿克苏等地区县市分布的亚洲飞蝗虫源形态变异研究及形态上的变异是否与所处的生态区域环境条件有关尚不明确,待进一步研究证明。
4 结 论
通过采集亚洲飞蝗在新疆分布的部分主要生态区域包括富蕴县、哈巴河县实验种群、和硕县、托克逊县、玛纳斯县、哈密市农十三师黄田庙尔沟散居型标本,利用数值分类学方法测量与分析亚洲飞蝗10个形态指标得出雌性与雄性亚洲飞蝗6个地理种群之间同在体长、前翅长度、前胸背板长度、前胸背板宽度4个性状存在地理差异性,头高与头宽两个性状6个地理种群之间不存在地理差异。雌性与雄性亚洲飞蝗不同地理种群之间的性状变异存在不一致性。散居型亚洲飞蝗分布的部分主要生态区域之间存在明显的形态变异现象。
MorphologicalVariationofGeographicalPopulationsofAsianLocustamigratoriainSomeEcologicalAreasinXinjiang
猜你喜欢
民间故事选刊·上(2022年1期)2022-02-09 09:52:22
小小说月刊·下半月(2020年9期)2020-10-09 11:22:13
小学阅读指南·低年级版(2020年6期)2020-07-26 14:30:50
华夏地理(2017年5期)2017-08-16 07:47:44
绿色科技(2017年3期)2017-03-14 20:11:52
保健与生活(2016年2期)2016-04-11 21:19:22
新农村(2016年10期)2016-03-06 05:37:46
故事大王(2014年5期)2014-06-25 01:42:31
草食家畜(2012年2期)2012-03-20 13:22:38
意林(2005年18期)2005-05-30 08:39:35
