
花后干旱对西藏青稞叶片水势、光合生理、籽粒表型和产量的影响
2018-07-25 06:42侯维海王建林胡单冯西博
中国农业科学 2018年14期
侯维海,王建林,胡单,冯西博
(西藏农牧学院植物科学学院,西藏林芝 860000)
0 引言
【研究意义】干旱是植物遭遇非生物胁迫因素中最复杂和最具破坏性的环境因子,引起植物形态、生理和代谢等多个方面的改变[1],最终影响光合作用和产量形成[2]。花后灌浆期是作物产量形成的关键时期,也是对土壤干旱最敏感的时期[3]。通过合理的灌溉措施维持灌浆期叶片光合生理对产量形成具有重要影响[4]。在西藏高原,青稞主产区多集中在海拔高、辐射强、降水少、蒸发量大的干旱和半干旱地区,该地区占西藏耕地面积的2/3。青稞花后灌浆期常遇到阶段性不同程度的干旱,导致青稞产量和品质下降[5]。另外,随着全球气候变化的影响,干旱发生的频率和严重性逐渐增加[6],干旱已成为西藏高原青稞高产和稳产的限制因素之一。因此,研究高原环境下干旱胁迫对青稞花后灌浆期叶片水势、光合生理、产量及籽粒表型关系,系统揭示花后干旱对青稞造成的影响,并提供无损、准确和快速判断青稞受旱情况的方法,将有助于指导青稞花后节水灌溉,并为抗旱品种筛选提供参考。【前人研究进展】干旱胁迫导致土壤水分亏缺,引起植物体内水分含量变化,进而影响叶片蒸腾作用带来的水分散失和体温调控[7]。叶片水势可直接反映植物体内水分状况,是评价植物水分亏缺最敏感的度量单位[8],与土壤含水量呈线性负相关,并与气孔导度存在密切关联[9]。另外,叶温冷却值与植物体内自由水含量和束缚水含量显著相关,能反映叶片水分状况,也可衡量植物受旱程度[7]。当植物发生缺水现象时,光合器官生理活性表现出高度敏感的复杂变化[10]。传统的叶片气体交换技术研究表明,干旱胁迫诱导植物叶片气孔关闭,导致气孔导度(gs)、净光合速率(Pn)、胞间 CO2浓度(Ci)和蒸腾速率(Tr)降低,最终引起作物光合能力的下降[11-13]。但当干旱胁迫达到一定严重程度时,简单的叶片气体交换仅能反映植物“表观”光合能力,而叶绿素荧光(CHIF)技术能反映“内在”光系统性能[14-15]。据研究,干旱胁迫导致植物光合器官PSII反应中心受损,光合电子传递能力降低,原初光能的捕获效率下降,光合作用受到不同程度抑制。大麦在严重或持久的干旱胁迫下,ΦPSII下降,而qN上升[16]。干旱胁迫降低小麦旗叶实际量子产量、ETR和NPQ值[17]。玉米随着干旱胁迫程度加剧,ΦPSII、ETR和qP均下降,而qN先升后降[18]。胡麻Fv/Fm、ΦPSII、qP明显下降,而qN明显上升[19]。随着干旱胁迫增加,结缕草叶绿素初始荧光(Fo)和qN逐渐增加,Fm、ΦPSII、Yield、ETR和qP呈逐渐下降趋势。彩叶草随着干旱胁迫增加,叶绿素含量、叶片含水量、Fo、Fv/Fm和 Fv/Fo表现逐渐降低趋势[20]。总之,干旱胁迫引起光合器官的破坏,反映为光合生理参数的改变,导致光合能力降低。在实践中常将叶片气体交换和叶绿素荧光技术有机结合,准确反映植物“表观”和 “内在”光系统状态和性能,完整解释外界环境胁迫对光合器官的影响。光合器官是植物生长、发育、干物质积累和产量形成的基础。以往研究表明,小麦[21-22]和大麦[23-24]花后干旱可导致植株整体发生改变,包括衰老加速、气孔导度降低,光合作用受阻,灌浆时间缩短,穗数、穗粒数、千粒重、籽粒产量的降低。大多数作物的产量以收获籽粒为主,籽粒品质与其大小、饱满度等表型性状有关,如粒长、粒宽、粒面积等,同时与籽粒粒级有关。但迄今为止,国内外相关研究鲜见报道。【本研究切入点】前人虽对干旱胁迫下小麦、水稻、玉米等作物光合、叶绿素荧光参数及产量性状有过研究,但在高原环境下系统深入研究干旱胁迫对青稞水势、光合特征、叶绿素荧光特性和产量的相关报道还较少,同时,与之紧密相关的籽粒表型性状及群体粒级的精细剖析也极少见。【拟解决的关键问题】本研究在高原环境下对干旱胁迫的青稞花后叶片水势、光合生理特性、产量因素、籽粒表型性状及群体粒级进行系统分析,以期获得以上指标对干旱胁迫的响应程度并建立响应曲线,探索判别青稞受旱程度的简单、准确和快速的方法,为西藏高原青稞生产提供精准的田间灌溉量,也为深入系统地了解干旱胁迫对青稞光合生理和产量提供理论基础。
1 材料与方法
1.1 试验材料与处理
选取西藏农区主栽春青稞品种“喜马拉雅22号”为供试材料,2016年3月20日在西藏农牧学院实习农场干旱棚内进行试验(经度94°25′,纬度29°59′,海拔2 960 m)。采用盆栽种植,人工模拟春青稞花后干旱胁迫。盆高30 cm,底部直径20 cm,盆面内径40 cm。每盆装质地相同的0—20cm耕层土壤28.6 kg,其类型为砂壤土,含有机质9.72 g·kg-1、全氮1.18 g·kg-1、全磷 0.32 g·kg-1、全钾 12.10 g·kg-1、碱解氮 83.60 mg·kg-1、速效磷 37.45 mg·kg-1、速效钾81.70 mg·kg-1。盆装土前,将土壤和肥料混伴,其中,氮磷钾混伴配比量分别为 75 mg N·kg-1,100 mg P2O5·kg-1,
100 mg KCl·kg-1。将饱满、大小一致、无病害种子按照每盆3行播于盆中,待3叶期,每盆边行定苗4株,中间行定苗5株,共13株。于开花后第3天进行干旱胁迫处理,以田间最大持水量的 75%—85%为对照(CK),设3个干旱胁迫处理水平,分别为轻度干旱胁迫(田间最大持水量为 65%—75%,CK灌水量的3/4,LD);中度干旱胁迫(田间最大持水量的 50%—55%,CK灌水量的2/4,MD);重度干旱胁迫(田间最大持水量的35%—45%,CK灌水量的1/4,HD)。控水开始时测定土壤含水量以达到预设水平,此后每隔2 d于16:00称重补水,持续处理13 d,随后进行相关指标测定。每个处理重复5次。控水前水分管理同对照,注意病虫草害发生,并于5月中下旬喷施三锉酮防治锈病。
1.2 测定项目
1.2.1 叶片水势(LWP)测定 选择晴朗无风天气,于11:30取受光方向一致的旗叶、倒2叶为测定对象,用打孔器打出10个直径为1 cm碎圆片,放置于WP4C型露点水势仪的样品盒内,进行水势测定。
1.2.2 旗叶蒸发冷却值和光合参数测定 采用Li-6400XT便携式光合仪(Li-Cor,Inc,USA)并以开放式气路系统方式对青稞旗叶蒸发冷却值和光合参数进行测定。设置气体流速为 500 μmol·s-1,3 cm×2 cm LED红蓝光光源提供1 300 μmol·m-2s-1的光合有效辐射(PAR),获得瞬时净光合速率(Pn)、气孔导度(gs)、胞间CO2浓度(Ci)、蒸腾速率(Tr)、大气CO2浓度(Ca)、空气温度(Tair)和叶片温度(Tleaf)。依公式计算气孔限制度Ls=1-Ci/Ca,叶蒸发冷却值ΔT=Tleaf-Tair。
1.2.3 旗叶叶绿素荧光参数测定 于9:00—11:30,取受光方向一致的旗叶,利用OS5P便携式脉冲调制叶绿素荧光仪(OPTI-science,USA)的3种测量模式,获取叶绿素荧光动力学参数。所有指标均技术重复6 次,生物学重复3次,并采用交叉测量(先每处理测一次,再循环 2次),保证各处理间的测定尽量在同一时间进行,以消除气象差异。在Fv/Fm模式下,叶片预先暗适应30 min,获得初始荧光(Fo)、最大荧光(Fm)、可变荧光(Fv),计算出PS II的最大量子效率Fv/Fm=(Fm-Fo)/Fm;在Yield模式下,叶片充分光照下适应30 min,获得稳态荧光(Fs)、光下最大荧光(Fms)、Yield=PS II的实际光量子产量ΦPSII=(Fms-Fs)/Fms;Kinetic模式下,叶片充分光照下适应30 min,获得光化学淬灭(qP)、非光化学淬灭(qN或 NPQ)、光合电子传递的相对速率(ETR)
1.2.4 室内考种 各处理材料成熟后,单独收获,分别脱粒,获取千粒重、籽粒产量、单株粒重、干物质积累量等数据。
1.2.5 籽粒表型性状和粒级分析 各处理取全部籽粒进行无影拍照,获得高质量电子图片。利用Image-pro plus软件分析籽粒表型性状(图1),测定粒二维面积、粒宽、粒长、粒直径、粒周长、粒圆度值指标。
为定量分析干旱胁迫对籽粒大小影响,以粒二维面积(Ga)为指标,将籽粒分为小粒(Ga<0.18 cm2)、中粒(0.18 cm2<Ga≤0.23 cm2)、大粒(Ga>0.23 cm2)3个等级,分析不同干旱胁迫对粒级的影响。
图1 按照粒二维面积大小对籽粒进行分级Fig. 1 Classification of seeds according to the size of two-dimensional area of grain
1.3 数据处理
利用 R软件进行简单线性回归分析和作图,SPSS 20进行Pearson相关分析。
2 结果
2.1 干旱胁迫对青稞不同叶水势(LWP)影响
与CK相比,LD处理的旗叶和倒2叶水势略有下降,降幅分别为0.24和0.04 Mpa(图2)。随着干旱胁迫加深,旗叶和倒2叶LWP明显下降,其中MD处理较CK降幅分别为0.82、0.61 Mpa,HD处理较CK降幅分别为2.26、1.32 Mpa。Pearson相关分析显示,干旱胁迫与旗叶和倒 2叶水势的相关系数均为0.92(P<0.05)。以干旱胁迫程度为自变量x,旗叶和倒2叶LWP为因变量y建立回归曲线(图3-A,图3-B),其回归方程分别为y=4.32x-5.94和y=2.73x-4.6,两者均为一次线性增函数。说明干旱胁迫均可明显引起旗叶和倒2叶LWP变化,且前者LWP对干旱胁迫更敏感。

图2 不同干旱胁迫下青稞旗叶和倒2叶水势Fig. 2 Water potential changes of flag leaf and the 2nd leaf from top in hulless barley under different drought stress
2.2 干旱胁迫对青稞旗叶蒸发冷却值(ΔT)影响
随着干旱胁迫的加重,旗叶ΔT呈逐渐增大趋势(图 3-C),其中 MD和 HD处理引起旗叶ΔT升高达1.67℃和2.97℃。同时,两者可用简单线性回归方程y=-3.57x+3.66描述,表明随着干旱胁迫的加重,植物冷却系统调控力度增加,阻碍了气体交换和热量散失,从而引起植物体温上升。相关分析表明,ΔT与干旱胁迫存在显著负相关(r=-0.978,P<0.05)。
2.3 干旱胁迫对青稞旗叶叶片光合参数的影响
花后干旱对青稞光合参数的影响如图4所示。LD处理下,旗叶Pn、gs、Ci分别较CK下降2%、7%、6%,Tr和Ls较CK增加1%和10.1%;MD处理下,旗叶Pn、gs、Ci、Tr分别较CK下降45%、72%、65%,67%,而Tr增加53.31%;HD处理下,旗叶Pn、gs、Ci、Tr分别较 CK 下降 88%、97%、99%,98%,而Tr增加63.4%。在 MD和 HD处理下,gs、Ci、Tr下降明显,而Tr的变化极小,其中gs分别降至0.06和0.01 mol H2O·m-2·s-1;Ci分别降至85.43 和1.56 μmol·mol-1;
Tr分别降至 1.36和0.08 g·m-2·h-1。以上表明,中、重度干旱胁迫导致气孔几乎关闭,Ls达到最大值,引起Ci和Tr显著下降,此时的水分含量已是维持青稞生命活动的极限水量。相关分析显示,干旱胁迫程度与gs和Ci的相关系数分别是0.967和0.964,均达显著水平(P<0.05),而与Ls的相关系数是-0.963,呈显著负相关(P<0.05)。
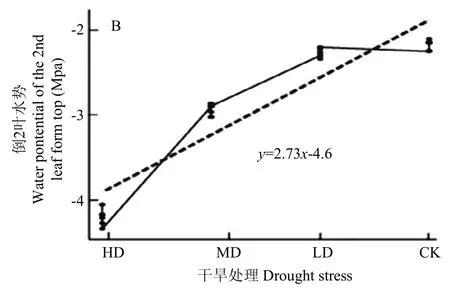
图3 干旱胁迫程度与叶水势和叶温的简单回归模型Fig. 3 Simple regression model of leaf water potential, leaf temperature and drought stress degree
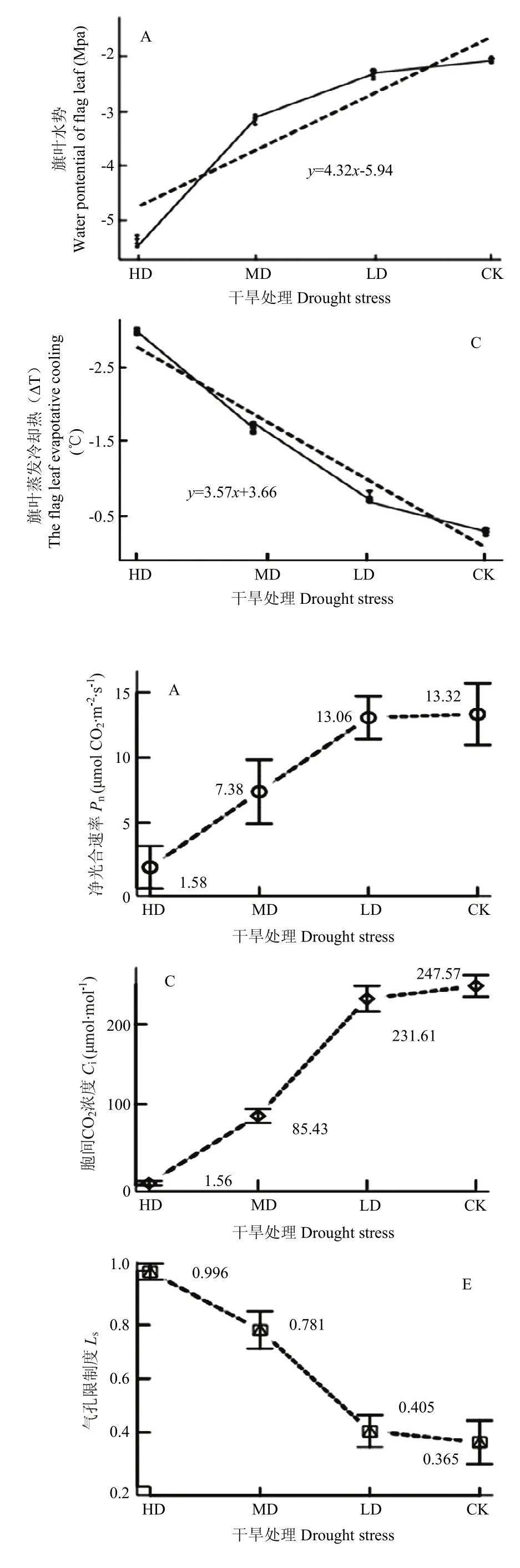
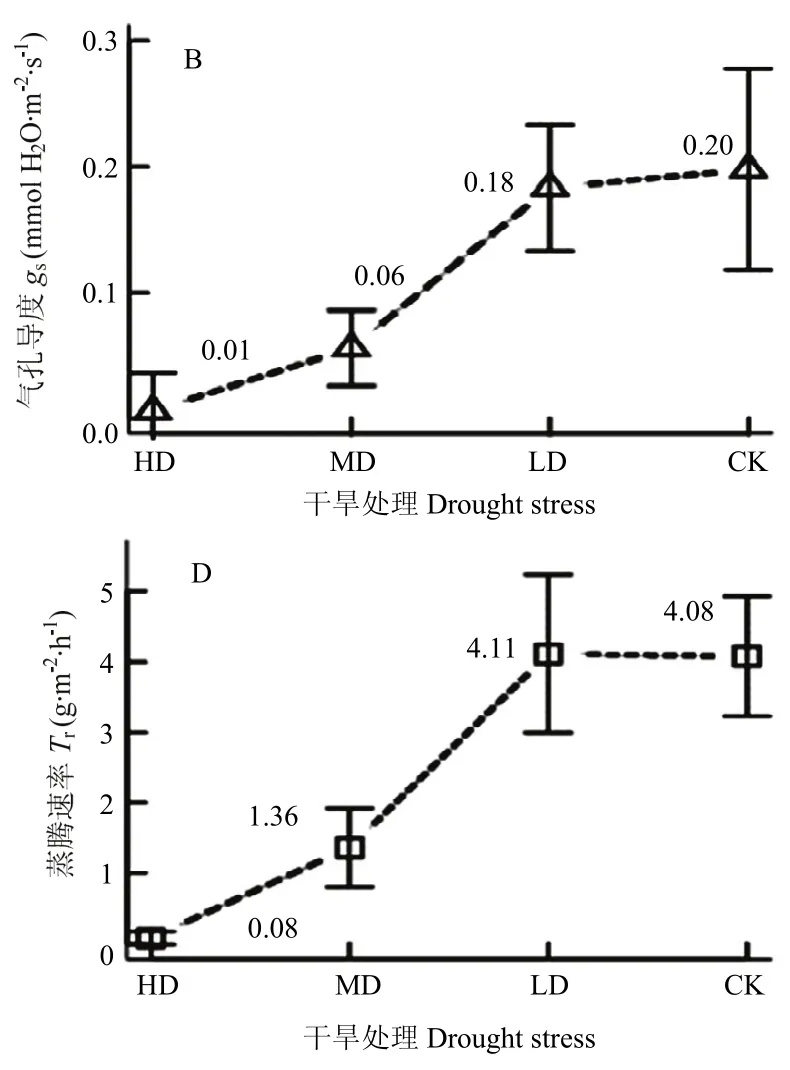
图4 干旱胁迫下青稞旗叶光合参数变化Fig. 4 Variation of photosynthetic parameters of flag leaf for hulless barley under different drought stress
2.4 干旱胁迫对青稞旗叶叶绿素荧光参数(ChIF)影响
2.4.1 干旱胁迫对初始荧光(Fo)、最大荧光(Fm)及PS II的最大量子产量(Fv/Fm)的影响 LD、MD和HD干旱胁迫处理导致Fo较CK升高2.9%、6.8%和13.5%(图5-A),Fo的增加表明PSII反应中心破坏或者可逆失活[25];Fm的大小反映了 PSII的电子传递能力,Fm的变化与Fo变化趋势相反(图5-B),即随着干旱胁迫增加,Fm较CK分别下降3.4%、16.8%和31.6%。在CK条件下,Fo最小,而Fm最大,表明此时的水分对青稞光合最适宜的。随着干旱胁迫的加剧,青稞旗叶PSII的电子传递能力逐渐下降。相关分析显示,干旱胁迫程度与Fm和Fo的相关系数分别为0.971和-0.981(P<0.05),表明干旱胁迫呈度与 Fm呈显著正相关,而与Fo呈显著负相关。
如图5-C所示,CK 的Fv/Fm值为0.78,LD、MD和HD处理的Fv/Fm值分别为0.76、0.71和0.63,Fv/Fm值的减小呈先慢后快的趋势。相关分析表明,干旱胁迫与Fv/Fm呈显著正相关,其相关系数达到0.966(P<0.05)。该结果表明,不同干旱处理下Fv/Fm值的降幅不同,但均导致青稞旗叶PSII活性中心受损,光合作用原初反应过程受抑制。
2.4.2 干旱胁迫对青稞旗叶实际光量子产量(ΦPSII)和电子传递速率(ETR)的影响 ΦPSII反映了植物实际光合效率,ETR为最大潜在相对电子传递速率,两者在无光抑制的光照条件下直接测得。由图 5-D和图5-E所示,青稞旗叶ΦPSII和ETR在LD、MD和HD处理间差异显著,均表现为LD>MD>HD。相关分析显示,干旱胁迫程度与ΦPSII和ETR呈极显著正相关,相关系数分别是 0.995和 0.992(P<0.05)。
2.4.3 干旱胁迫对青稞旗叶荧光淬灭系数影响 光化学淬灭(qP)在CK和LD条件下几乎一致,之后随着干旱程度加重,qP下降较快,而非光化学淬灭(NPQ)表现出与qP相反的趋势,其值逐渐增加(图5-F、G)。相关分析显示,干旱胁迫程度与 qP显著正相关,而与 NPQ呈显著负相关,相关系数分别为0.959和-0.98(P<0.05)。以上结果表明,干旱胁迫增加了PSII的天线色素耗散热的能力,降低了干旱胁迫对PSII的破坏。
2.5 不同干旱胁迫对青稞千粒重(TKW)、单穗粒重(SGW)、干物质积累量(DMC)、籽粒产量(GW)和经济系数(EC)的影响
如表1所示,LD、MD、HD处理下,青稞TKW比 CK分别下降 19.12%、25.78%、41.06%;SGW分别下降5.08%、14.28和31.25%;DMC分别下降4.32%、9.35%、28.06%;GW 分别下降 8.47%、18.64%、44.07%;EC分别降低 4.35%、10.25%、22.25%。相关分析表明,干旱胁迫程度分别与TKW、SGW、GW和EC呈显著正相关,其相关系数依次为0.984、0.955、0.954和0.958(P<0.05)。
以干旱胁迫程度x为自变量,分别与应变量y(TKW、GW、DMC、Ec、SGW)做简单回归分析(图6),发现x和y两者之间存在明显的相关性,呈一次函数变化。干旱对TKW的拟合回归方程为y=26.46x+匀23.495;对GW拟合回归方程为y=33.6x+27.5;对DMC拟合回归方程为y=49.6x+93.5;对EC拟合回归方程为y=0.12x-0.31;对 SGW 拟合回归方程为y=2.65x+2.12。拟合方程的斜率值均为正值,表明自变量x与应变量y均呈正相关。这一结果与相关性分析相似。

表1 不同干旱胁迫下的青稞产量参数Table 1 Analysis on yield reduction of hulless barley yield under different drought stress
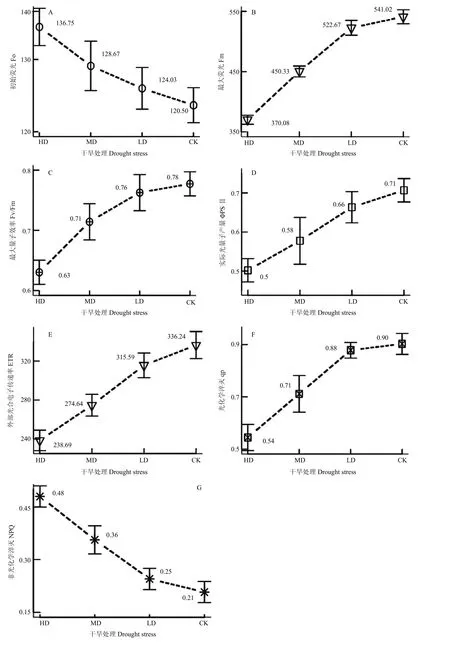
图5 不同干旱胁迫下青稞旗叶叶绿素荧光参数Fig. 5 Chlorophyll fluorescence parameters of flag leaf form hulless barley under different drought stress
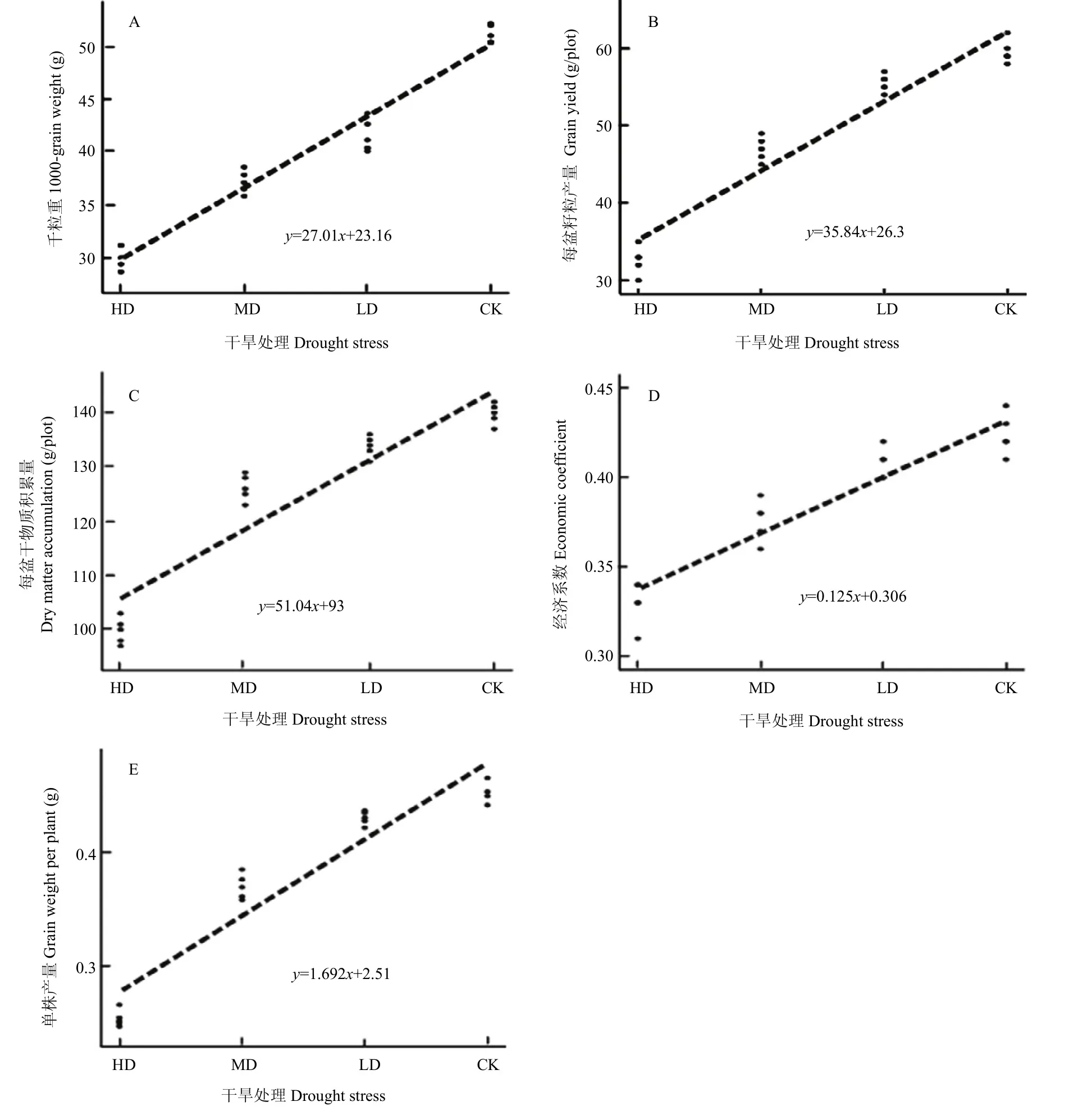
图6 干旱胁迫程度与青稞产量参数的简单回归模型Fig. 6 A simple regression model for production parameters of hulless barley and drought stress degree
2.6 不同干旱胁迫下,籽粒表型的相关分析及回归分析
利用Image-pro plus软件对不同干旱胁迫的青稞籽粒表型性状进行精细分析(表2)。不同干旱胁迫下,青稞籽粒表型表现出明显规律性变化(图7)。随着干旱胁迫增加,粒二维面积、粒周长、粒长、粒宽均有减少,而粒圆度值增大。表明干旱改变了籽粒的大小,影响了籽粒的性状,随着干旱胁迫的加剧,这种改变能力越大。相关分析表明,干旱胁迫对青稞籽粒表型性状影响明显,其中干旱对粒周长影响呈极显著正相关,相关系数为0.994(P<0.01);对粒面积、粒直径和粒长呈显著正相关,相关系数分别为0.976,0.977和0.988(P<0.05);与粒宽呈正相关,相关系数为 0.914(P>0.05),而与粒圆度值呈负相关,相关系数是-0.939(P>0.05)。
对粒二维面积(Ga)分级显示(表3),随着干旱胁迫增加,小粒(Ga<0.18 cm2)二维面积占比明显增加,中粒(0.18cm2<Ga≤0.23 cm2)二维面积占比先增后降,大粒(Ga<0.23 cm2)占比明显降低。该结果进一步说明,干旱对籽粒大小具有明显影响,导致小粒明显增加,而大粒明显降低。
为定量描述干旱胁迫对籽粒表型性状影响程度,利用简单回归分析建立了以干旱胁迫程度x为自变量,以种子表型性状为因变量y的线性回归模型(图7)。结果显示,干旱对粒二维面积的回归方程为y=0.076x+0.159;对粒直径的回归方程为y=0.092x+0.449;对粒周长的回归方程为y=0.291x+1.585;对粒圆度值的归回方程为y=-0.048x+1.247;对粒长的回归方程为y=0.118x+0.677;对粒宽的回归方程为y=0.075x+0.328。由此可见,干旱对籽粒表型性状影响差异较大,根据回归方程斜率值大小,推断干旱对粒周长影响最大,其次为粒长和粒直径,再次为粒面积和粒宽。

表2 不同水分胁迫对青稞籽粒表型性状参数的影响Table 2 Phenotypic character parameters of hulless barley grain under different drought stress
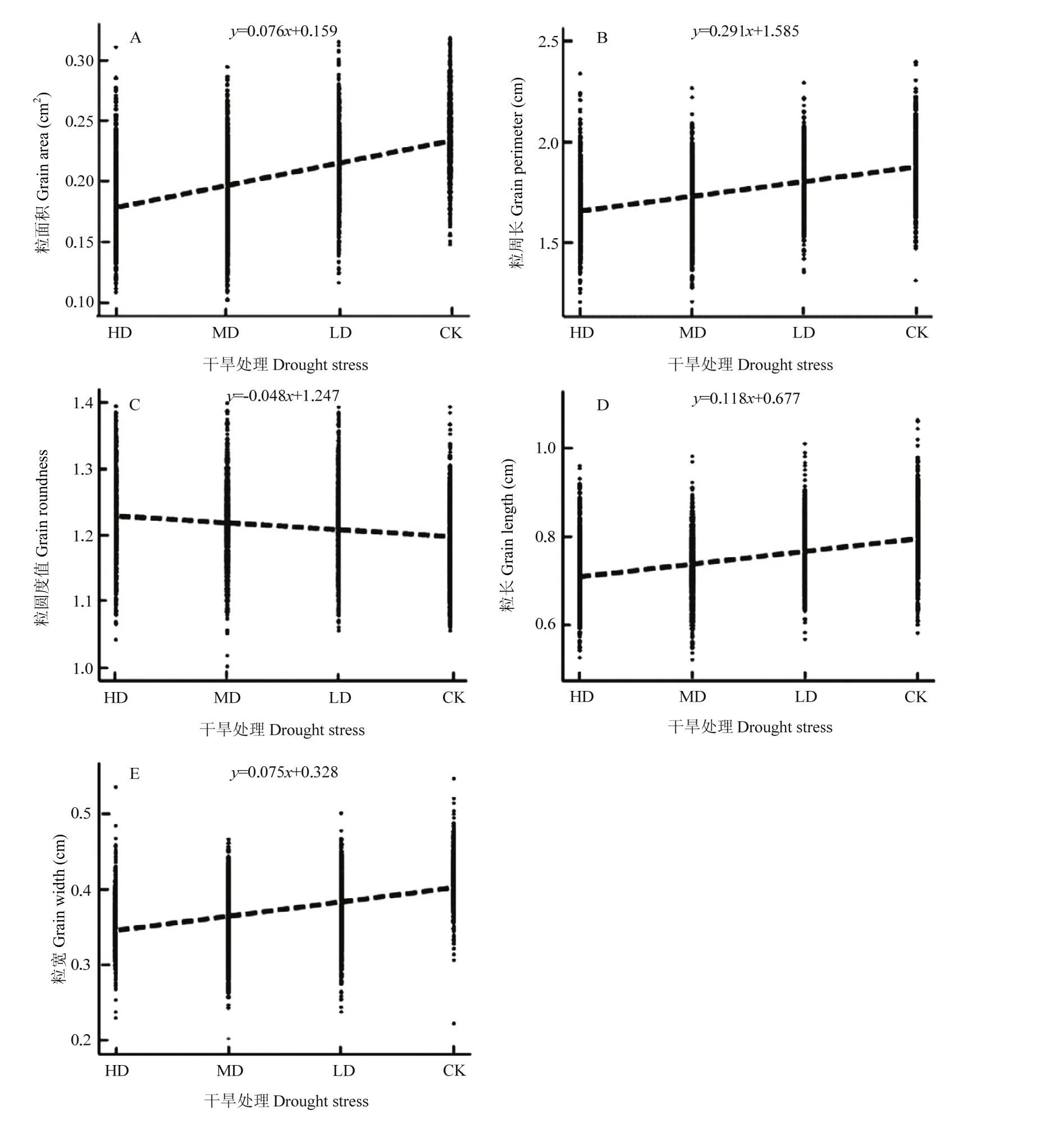
图7 干旱胁迫程度与青稞籽粒表型的简单回归模型Fig. 7 A simple regression model for the phenotypic of hulless barley and drought stress degree

表3 不同干旱处理对青稞籽粒粒级影响Table 3 Effect of different drought stress on grain classification of hulless barley under different drought stress
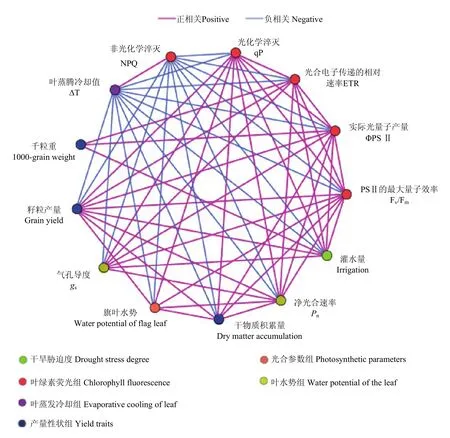
图8 干旱胁迫、叶水势、光合参数、叶绿素荧光参数和产量指标之间的Pearson相关网络图Fig. 8 Pearson correlation network diagram on drought stress, leaf water potential, photosynthesis parameter, chlorophyll fluorescence parameter, and yield index
2.7 干旱胁迫程度、叶水势、光合、叶绿素荧光参数及产量因子的Pearson相关网络图
ΔT和NPQ与大部分参数呈直接负相关,而与个别参数虽无直接相关,但存在间接相关(图 8)。灌水量与gs、qP、ETR、ΦPSII、Fv/Fm和籽粒产量呈直接正相关;旗叶水势与qP、Fv/Fm、Pn、籽粒产量和干物质积累量呈直接正相关;Pn与ΔT和NPQ呈直接负相关,除与灌水量存在间接正相关外,皆与其他指标呈直接正相关。gs与Pn、灌水量、Fv/Fm、ΦPSII、ETR和qP呈直接正相关,而与ΔT和NPQ呈直接负相关;千粒重与ETR、ΦPSII和灌水量呈直接正相关。以上结果表明,在干旱胁迫下,青稞光合生理、产量指标等参数间关系比较复杂,既存在直接影响,也具有间接影响。
3 讨论
3.1 叶水势与干旱胁迫的关系分析
植物叶水势与受旱胁迫直接相关[26],与土壤水分含量呈良好的线性关系[13,27]。本研究表明,青稞功能叶水势可灵敏反映其受旱程度,且旗叶水势较倒2叶对干旱胁迫更敏感,同时相关分析显示叶水势与气孔导度呈间接正相关(图 7)。据研究,叶水势的降低与气孔开闭程度有关[27-28]。由此推测,生理活性较旺盛的旗叶遇到干旱胁迫时,气孔导度下降快,阻碍了蒸腾作用,导致蒸腾拉力减弱和膨压下降,土壤水向顶部运输压力不足,从而旗叶较倒2叶水势下降更多。因此,青稞旗叶水势可直观、快速判断整个植株持水状况,反映土壤水分亏缺程度,可用于快速和精确地指导节水灌溉。
3.2 叶蒸发冷却值与干旱胁迫的关系分析
作物通过气孔的蒸腾作用散失水分,以降低体温维持正常的生理活动。当土壤水分不足时,会影响蒸腾作用水分散失和体温调控,叶蒸发冷却值(ΔT)可衡量植物受干旱胁迫程度的大小[9]。本研究发现,ΔT与干旱胁迫呈显著负相关(P<0.05)且呈一次增函数,同时相关性网络图显示(图7),ΔT与gs、ΦPSII、Fv/Fm呈负相关。这可能与干旱胁迫诱导气孔导度下降,影响植物与外界环境的气体和热量交换,造成叶冷却系统受损有关。相似的研究结果也在干旱胁迫的山毛榉、高粱等植物中报道过[8,29]。因此,ΔT和LWP均可作为衡量土壤水分或植物受旱程度的直接指标,甚至ΔT比LWP更敏感。
3.3 光合参数与干旱胁迫的关系
植物光合器官对干旱胁迫敏感,干旱胁迫下植物光合作用的反应过程和机制与植物生长阶段、基因型、干旱程度等因素有关[30]。本研究在青稞花后灌浆期进行干旱胁迫,且品种相同,因此,干旱胁迫对青稞光合参数的变化应当取决于干旱程度。本研究发现,随着干旱胁迫的加重,青稞旗叶Pn、gs、Ci和Tr均表现出不同程度的下降趋势,而Ls呈逐渐增加趋势,其中各参数快速下降或上升的拐点均发生在中度干旱胁迫附近,推测当土壤含水量为最大田间持水量的 50%时,光合作用将明显降低。相关分析表明,干旱胁迫与gs和Ci呈显著正相关,而与Ls呈显著负相关;与Pn和Tr没有直接相关性,但存在间接正相关。结果进一步表明,干旱对气孔调控显著,gs降低,Ls增加,维持了组织水分含量。本研究结果与大麦、小麦在干旱胁迫下植株的衰老,叶绿素的分解、光合作用的抑制相似[3,24],也进一步说明干旱胁迫加速了青稞叶片衰老,引起整个植株持绿性下降,导致叶片捕光面积减少,气孔导度下降并伴随着胞间 CO2浓度降低,进而导致光合作用的下降[30-31]。
3.4 叶绿素荧光参数与干旱胁迫的关系
叶绿素荧光常用于监测、定量分析和比较植物对胁迫的反应。当植物受到干旱胁迫时,叶片气孔关闭导致光抑制产生,引起 PSII反应中心损伤,使得叶绿素荧光参数发生改变[33-34]。本研究发现,随着干旱胁迫的加重,青稞旗叶 Fo逐渐升高,在中度和重度干旱胁迫下,Fo升高较快,Fm则表现出与Fo相反的变化趋势。说明轻度干旱胁迫对青稞旗叶 PSII反应中心的破坏较小,随着干旱胁迫加剧,PSII反映中心遭到了不易逆转的破坏或可逆失活,其色素吸收的能量以热和荧光散失的形式增加,而用于光合作用的能量逐渐减少。Fv/Fm能快速和有效反映PSII最大光化学量子效率。低的 Fv/Fm 表明植物处于逆境胁迫条件下,PSII受到光抑制或下调。因此,Fv/Fm值的大小可指示作物的健康状况。大部分作物健康的光合组织Fv/Fm平均值约在0.83,低于该值表明植物处于生理修复状态[35]。例如,在水分[36]、温度[37]、病原物侵染的生物胁迫[38]等外界环境因素胁迫下,Fv/Fm明显下降。本研究发现,在正常灌水条件下,青稞旗叶LWP为-2.09 Mpa时,Fv/Fm为0.78,低于前人报道的0.83。推测青稞可能处于生理修复状态,这与西藏林芝地区低氧、低温、持续阴雨等特殊的气候环境有关,也可能归因于高原作物在长期的进化、演化和选择中形成与低地作物相差异的光合适应模式。此后,随干旱胁迫增加,青稞旗叶LWP和Fv/Fm呈逐渐降低趋势,说明青稞已受到水分胁迫,并引起 PSII的损伤,光合初反应受到抑制。中度和重度干旱胁迫下,Fv/Fm有一定程度的下降,可作为干旱胁迫引起植物可看见的生理紊乱之前的一种潜在提示,这在山毛榉植物中也得到了验证[29]。
有研究学者认为,Fv/Fm对早期轻度干旱不敏感,而Fs、ΦPSII、NPQ能比Fv/Fm更敏感地捕捉轻度干旱胁迫对光合器官造成的损伤[39]。前人研究表明,水分胁迫并未显著影响小麦和硬粒小麦的 PSII 的原初光化学反应[40]。但相反的结论认为Fv/Fm、NPQ 、qP、Fo均有改变[41]。本研究在轻度干旱胁迫下,发现旗叶Fs、ΦPSII、NPQ变幅比Fv/Fm更大,表明以上参数对干旱胁迫非常敏感,这与前人研究成果相似。在中度和重度干旱胁迫下,旗叶ΦPSII、ETR、qP逐渐降低,表明PSII反应中心受到破坏,光合电子传递活性降低,原初光能的捕获效率下降,光合作用受到不同程度抑制;而Fs和NPQ呈增大趋势,反映PSII天线色素吸收的过量光能以热和发射荧光的形式耗散,从而保护光合器官,已有研究认为这是植物适应干旱胁迫的一种保护机制[42]。
3.5 产量性状、种子表型和群体粒级与干旱胁迫的关系
青稞花后干旱处理下,千粒重、单穗粒重、籽粒产量、经济系数、干物质积累量均有不同程度的降低,并引起籽粒表型参数下降,表现为籽粒横向变窄、纵向缩短、粒二维面积降低,同时群体粒级发生改变,小粒占比明显增加,而大粒占比呈明显下降。上述性状均属于数量性状遗传,相关性网络图显示(图7),干旱胁迫与各观测参数联系紧密,既存在直接相关也存在间接相关,其中干旱胁迫与gs、qP、ETR、ΦPSII、Fv/Fm、籽粒产量和千粒重均呈直接正相关,而与ΔT和NPQ呈直接负相关。该结果表明,干旱胁迫引起光合器官破坏,导致碳同化能力下降,最终造成青稞产量性状,种子表型和群体粒级改变,这与前人研究结果一致[23-24]。
4 结论
在不同的梯度干旱胁迫下,灌溉水量与叶水势和叶蒸发冷却值(ΔT)呈线性相关,两者均能灵敏反映干旱胁迫程度(或土壤水分)大小,可代表青稞受旱程度;干旱胁迫引起青稞光合器官的破坏,导致光合及叶绿素荧光参数的改变;产量参数、籽粒表型性状、群体粒级均随干旱胁迫的加重而逐渐降低。
猜你喜欢
作物学报(2022年9期)2022-07-18
星星·诗歌原创(2022年1期)2022-04-16
干旱地区农业研究(2021年6期)2021-12-08
干旱地区农业研究(2021年6期)2021-12-08
今日农业(2021年17期)2021-11-26
西藏人文地理(2020年4期)2020-11-19
农村实用技术(2020年9期)2020-11-19
翠苑(2019年4期)2019-11-11
浙江农业科学(2019年11期)2019-11-09
湖北林业科技(2019年4期)2019-09-16
