
西部传统发酵乳品中乳酸菌筛分及其亚硝酸盐降解能力
2018-07-24 08:45张兴吉葛武鹏李小鹏
食品科学 2018年14期
张兴吉,葛武鹏,*,刘 阳,王 瑞,王 智,李小鹏
(1.西北农林科技大学食品科学与工程学院,陕西 杨凌 712100;2.陕西百跃优利士乳业有限公司,陕西 西安 710000;3.咸阳市食品药品检测检验中心,陕西 咸阳 712000)
我国传统发酵乳品资源丰富,西部牧区是我国传统发酵乳品的主要产区,生活在新疆、青海、甘肃、内蒙古、西藏等牧区的牧民利用马乳、牛乳、骆驼乳、羊乳等为原料,通过传统的发酵方式自制成各类发酵乳品食用[1-3]。西部牧区因其特殊的地理气候环境以及牧民的生活习惯造就了其独特的微生物资源[4-5],在发酵乳品制作过程中,伴随着以乳酸菌为主的微生物生长代谢活动[6]。乳酸菌是传统发酵乳品中微生物菌群的优势菌,且传统发酵乳品的保健功能离不开乳酸菌的作用[7]。目前乳酸菌分离鉴定方法分为常规鉴定、快速鉴定、基因分子鉴定方法等,其中常规鉴定和快速鉴定方法存在费时费力、准确率低于基因分子鉴定方法等缺点,而基因分子鉴定方法因其具有鉴定结果准确、方法成熟以及省时省力等优点而被视为常用的菌株鉴定方法[8]。
在食品行业中,硝酸盐和亚硝酸盐虽然被允许作为食品添加剂使用,在食品工业中起到重要的作用。然而硝酸盐和亚硝酸盐含量超过安全范围将会对人体造成危害,特别是亚硝酸盐危害健康更甚,过量会积累多种疾病,如癌症、克山病、氰紫症、智力下降等[9-12]。在牲畜喂养过程中,如果环境、饲料、饮水中硝酸盐、亚硝酸盐含量过高,可能导致其乳中的亚硝酸盐含量超标[13-14],故亚硝酸盐含量一直作为乳品质量检测的重要指标,受到广泛关注。文献表明,乳酸菌具有降解亚硝酸盐特性[15]。乳酸菌具备亚硝酸盐降解能力主要因其在生长代谢过程中会产生乳酸和一系列酶对亚硝酸盐具有降解作用[16],同时在发酵体系中乳酸菌成为优势菌时,可抑制其他杂菌的生长,从而抑制亚硝酸盐的产生。目前,乳酸菌降解亚硝酸盐的研究多集中于泡菜和酸菜等资源上,而对于从传统发酵乳品中分离出乳酸菌进行亚硝酸盐降解能力分析的相关研究鲜有报道。本研究通过对西部牧区传统发酵乳品中乳酸菌分离鉴定及其优势菌种亚硝酸盐降解能力分析比较,分别从地区、乳品品种及乳酸菌种类等方面探究其能力差异,为乳酸菌新资源发掘及其合理化利用提供参考依据[17]。
1 材料与方法
1.1 材料与试剂
1.1.1 样品
2016年8月分别在青海牧区采集15 份酸牦牛奶和2 份奶渣,甘肃牧区采集5 份酸牦牛奶,内蒙古牧区采集21 份酸马奶和8 份酸驼奶,新疆牧区采集20 份酸马奶和9 份酸驼奶,西藏牧区采集24 份奶渣样品,装入50 mL无菌离心管中,冰盒冷链至实验室,放入4 ℃冰箱储存备用。
1.1.2 培养基与试剂
MRS肉汤培养基、MRS琼脂培养基 北京陆桥技术有限公司。
革兰氏染色试剂盒 北京陆桥技术有限公司;聚合酶链式反应(polymerase chain reaction,PCR) Premix TaqTM日本Takara公司;放线菌酮 美国Inalco公司;亚硝酸钠、对氨基苯磺酸、盐酸萘乙二胺、乙酸锌、亚铁氰化钾、硼砂 美国Sigma-Aldrich公司;细菌基因组DNA提取试剂盒 北京天根生化科技有限公司;引物上海生工生物工程股份有限公司。
1.2 仪器与设备
E100生物显微镜 日本尼康公司;T100TMThermal Cycler PCR仪、E1617-T130 plus凝胶成像系统 美国Bio-Rad公司;DYY-6C电泳仪 君意东方公司;DRP-9162型电热恒温培养箱 上海森信实验仪器有限公司;YXQ-LS-50SII-01-00立式压力蒸汽灭菌器 上海博迅实业有限公司;HC3018型高速离心机 安徽科大中佳有限公司;UV-2600型紫外分光光度计 日本岛津公司。
1.3 方法
1.3.1 乳酸菌分离纯化
将样品进行梯度稀释,吸取0.5 mL样品于含4.5 mL无菌水的离心管中,涡旋混匀,将样品稀释至10-5,选取10-3、10-4、10-5三个梯度的稀释液在MRS固体培养基(含40 mg/L放线菌酮)上进行平板涂布,于37 ℃厌氧培养48 h。观察并记录菌落的特征,经肉眼及显微镜下菌落形态的观察分出不同菌落,对其进行革兰氏染色证实为革兰氏阳性的菌落,在MRS琼脂培养基上连续划线2 次,进行纯化。将培养好的菌种编号并接种于30%甘油管中,置于-80 ℃冰箱中保存。
1.3.2 乳酸菌鉴定
基因组DNA提取:按照细菌基因组DNA提取试剂盒明示的方法提取乳酸菌DNA。
PCR扩增:分别对待测菌的16S rDNA序列27F(5’-AGAGTTTGATCCTGGCTCAG-3’)和1492R(5’-GGTTACCTTGTTACGACTT-3’)进行扩增。PCR体系:DNA模板1 μL,正反向引物各1 μL(20 pmol/μL),超纯水22 μL,PCR Premix TaqTM25 μL。扩增程序:94 ℃预变性5 min;94 ℃变性30 s,55 ℃退火30 s,72 ℃延伸2 min,循环25次;72 ℃延伸10 min。
PCR扩增产物经质量分数为1.5%的琼脂糖凝胶电泳检测后,将出现条带的PCR产物交由上海生工生物工程股份有限公司进行序列测定。测序结果根据GenBank数据库比对后,以相似度在99%以上为标准确定分离菌株的属种。
1.3.3 乳酸菌亚硝酸盐降解能力分析
1.3.3.1 菌株培养
将待测菌株接种于MRS肉汤培养基中于37 ℃活化2 代后,以体积分数1%的量接种到8 mL含有150 mg/L亚硝酸钠的MRS肉汤培养基中,于37 ℃静置培养24 h[18-19]。
1.3.3.2 亚硝酸盐测定
采用比色法对乳酸菌降解亚硝酸盐能力进行测定。菌株培养24 h后,于8 000×g离心10 min,取0.15 mL上清液依次加入0.75 mL饱和硼砂溶液、0.3 mL 106 g/L亚铁氰化钾溶液、0.3 mL 220 g/L乙酸锌溶液,然后加入28.5 mL超纯水,混匀,室温放置30 min,于6 500×g离心5 min。取20 mL上清液加入比色管中,加入1 mL4 g/L对氨基苯磺酸溶液,摇匀,静置5 min,再加入0.5 mL2 g/L盐酸萘乙二胺溶液,超纯水定容至25 mL,摇匀,静置15 min。零管调零,1 cm比色杯于538 nm波长处测吸光度,同时以不接菌的无亚硝酸钠MRS肉汤、不接菌的含150 mg/L亚硝酸钠MRS肉汤为空白对照[18]。通过亚硝酸钠标准曲线得到空白对照和样品中亚硝酸钠浓度。
标准曲线:吸取0.00、1.00、2.00、3.00 mL亚硝酸钠标准使用液(5.0 μg/mL),分别置于25 mL比色管中,加入1 mL4 g/L对氨基苯磺酸溶液,充分混匀,室温静置5 min,再加入0.5 mL2 g/L盐酸萘乙二胺溶液,用超纯水定容至25 mL,充分混匀,室温静置15 min。空白调零,用1 cm比色杯于538 nm波长处测吸光度,绘制标准曲线,R2应大于0.999。
菌株亚硝酸盐降解能力基于培养基中亚硝酸盐的降解率计算[19]。公式如下:
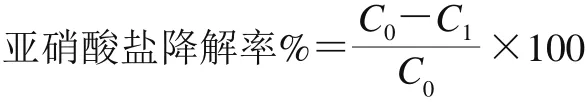
式中:C0为不接菌的含150 mg/L亚硝酸钠MRS肉汤培养基中亚硝酸盐浓度/(mmol/L);C1为测试样品中亚硝酸盐浓度/(mmol/L)。
1.4 数据处理
实验重复3 次,使用SPSS 19.0和Excel软件对实验数据进行处理分析,结果采用 ±s表示,显著性水平设定为P小于0.05。通过频数统计对分离出的乳酸菌亚硝酸盐降解率的不同区间进行归纳分类。
2 结果与分析
2.1 乳酸菌分离鉴定
从青海、甘肃、新疆、内蒙古、西藏5地104 份传统发酵乳品中共分离得到275 株乳酸菌,鉴定出6 个属23 个种:分别为Lactobacillus属12 种,Leuconostoc属4 种,Enterococcus属3 种,Lactococcus属2 种,Streptococcus属1 种,Pediococcus属1 种。其中,地区间比较,青海、甘肃、内蒙古、新疆、西藏样品中分别分离出44、11、76、73、71 株菌;乳品品种间比较,酸牦牛奶、酸马奶、酸驼奶、奶渣中分别分离出52、112、37、74 株菌。菌株信息如表1所示。
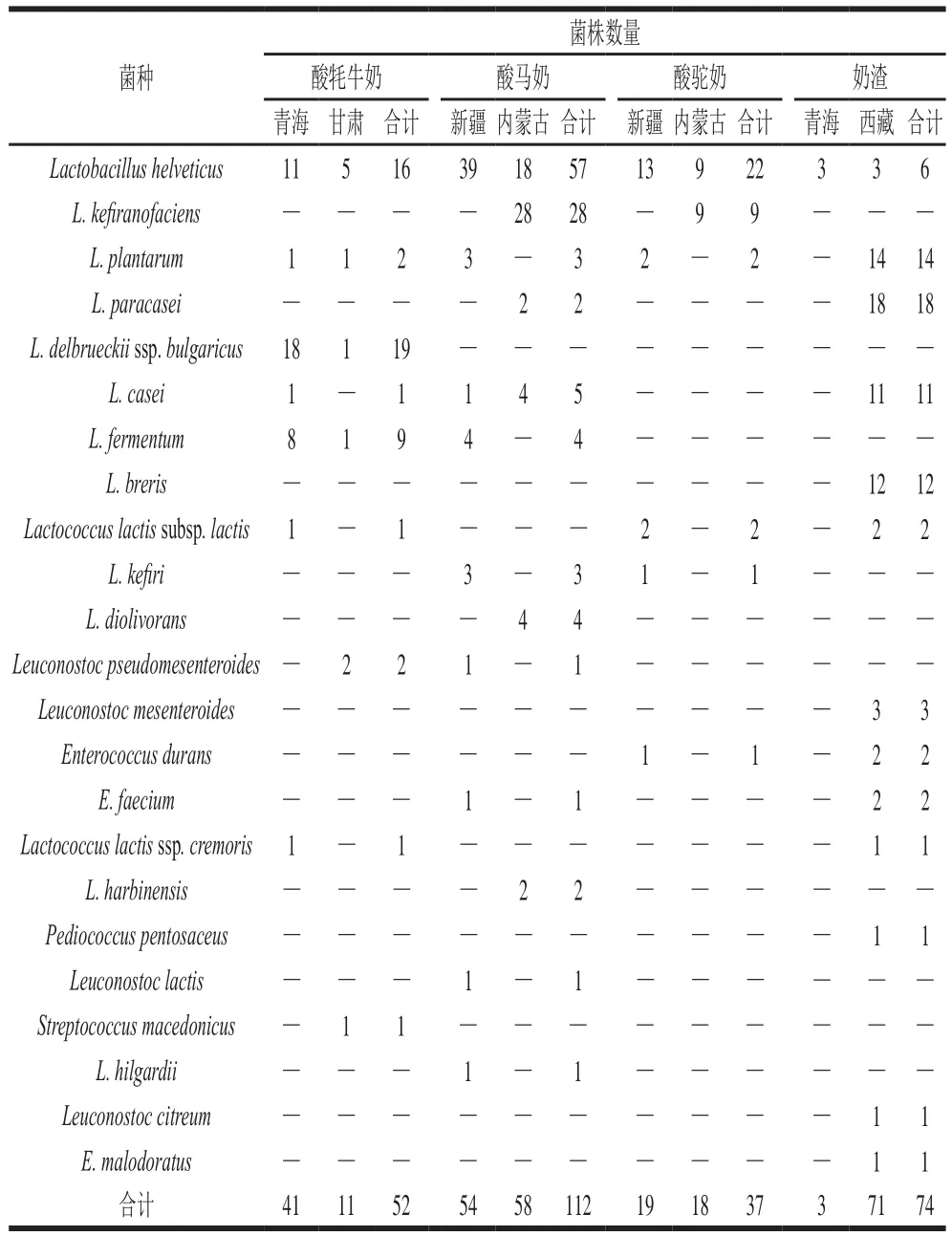
表1 西部地区分离乳酸菌菌株信息Table1 Information about LAB isolated from fermented milk products from western China
不同地区因其海拔、地貌、温度、气候、季节以及牧民生活习惯等因素导致不同发酵乳品中存在着复杂多样的菌群微生态结构,具有特定的优势乳酸菌类群,在分离出的23 种乳酸菌中,突出的表现在于:L. helveticus为青海、甘肃、新疆、内蒙古、西藏5 个地区发酵乳品中共有菌株;L. helveticus和L. plantarum为酸牦牛奶、酸马奶、酸驼奶、奶渣4 种传统发酵乳品中共有菌株。不同地区、不同发酵乳品中乳酸菌组成存在明显差异,以菌株筛分率为基准,青海地区发酵乳品中优势菌为L.helveticus和德式乳杆菌保加利亚亚种(L. delbrueckii ssp.bulgaricus);新疆和甘肃地区发酵乳品中优势菌均为L. helveticus;内蒙古地区发酵乳品中优势菌为马乳酒样乳杆菌(L. kefiranofaciens)和L. helveticus;西藏地区发酵乳品中优势菌为L. breris、L. plantarum、干酪乳杆菌(L. casei)和L. paracasei。酸牦牛奶中优势菌为L.helveticus和L. delbrueckii ssp. bulgaricus;酸马奶和酸驼奶中优势菌均为L. kefiranofaciens和L. helveticus;奶渣中优势菌为L. breris、L. plantarum、L. casei和L. paracasei。本研究的优势菌筛分结果与先前研究相一致,L. helveticus在大多数传统发酵乳品中都具有较高的筛分频率,李远等[20]对新疆不同地区酸驼奶中乳酸菌进行分离鉴定,结果表明L. helveticus为新疆酸驼奶中优势菌株。Sun Zhihong等[21]对新疆、内蒙古、青海地区酸马奶中乳酸菌多样性进行了系统分析发现L. helveticus为新疆酸马奶中优势菌株,而L. casei、L. helveticus和L. plantarum在内蒙古酸马奶中分离出的频率较高。Watanabe等[22]对蒙古国中央省等6 个地区传统发酵乳品中乳酸菌多样性进行研究,结果表明L. helveticus和L. kefiranofaciens为蒙古国中央省酸马奶中的优势菌株。Airidengcaicike等[23]对西藏酸牛奶中乳酸菌进行筛分,发现藏北地区的优势菌株为发酵乳杆菌(L. fermentum)和L. casei,藏南地区的优势菌株则为L. casei和L. plantarum。
目前,有关传统发酵乳品中乳酸菌多样性报道较多,相同地域可能存在不同的优势乳酸菌类群。Sun Zhihong等[24]从青海地区43 份自然发酵牦牛奶中分离出148 株乳酸菌,发现嗜热链球菌(S. thermophilus)、L. delbrueckii ssp. bulgaricus、L. plantarum和乳酸乳球菌乳酸亚种(Lactococcus lactis ssp. lactis)为酸牦牛奶中的优势菌群。Bao Qiuhua等[25]对甘肃省甘南地区酸牦牛奶样品中乳酸菌多样性进行分析,指出S. thermophilus、肠膜明串珠菌(Leuconostoc mesenteroides ssp. mesenteroides)和L. helveticus为甘南地区酸牦牛奶的优势菌群。这一结果与本研究存在不同,本研究从青海、甘肃采集的样品中未分离出S. thermophilus,可能是由于酸牦牛奶发酵时间以及采样季节和采样点不同造成的。
2.2 乳酸菌亚硝酸盐降解能力分析
2.2.1 不同来源乳酸菌亚硝酸盐降解率频数统计
对275 株乳酸菌亚硝酸盐降解率进行频数统计,结果如表2所示,从总体菌株来看,亚硝酸盐降解率范围为4.8%~99.9%,几何均值为77.3%,其中仅25%的菌株亚硝酸盐降解率在74%以下,50%的菌株亚硝酸盐降解率在91.3%以下,8 株菌亚硝酸盐降解率达到99.9%,分离菌株表现出较好的亚硝酸盐降解能力。不同地区、不同发酵乳品中的乳酸菌在降解率范围和几何均值上均具有显著差异,其中西藏地区和奶渣中的乳酸菌显示了最高的亚硝酸盐降解率。

表2 不同来源乳酸菌亚硝酸盐降解率频数统计Table2 Frequency statistics for nitrite degradation rate of LAB isolated from different sources
2.2.2 不同种类优势乳酸菌亚硝酸盐降解率频数统计
本研究中菌株数量大于6的乳酸菌种类共有8 种,对8 种优势乳酸菌亚硝酸盐降解率进行频数统计,如表3所示,降解能力因乳酸菌种类不同存在差异性,不同优势菌种降解率范围和几何均值差异明显,其中L. breris、L. plantarum、L. paracasei显示了稳定高效的降解能力,且L. plantarum降解能力最强,L. fermentum降解能力最差。

表3 不同种类优势乳酸菌降解亚硝酸盐频数统计Table3 Frequency statistics for nitrite degradation rate of different dominant LAB
2.2.3 不同地区发酵乳品中乳酸菌亚硝酸盐降解能力比较
将青海、甘肃、新疆、内蒙古、西藏5 地发酵乳品中乳酸菌亚硝酸盐降解率进行方差分析,比较不同地区菌株在降解能力上的差异。如图1所示,不同地区菌株降解能力存在显著性差异(P<0.05),来源于西藏的菌株降解能力显著高于青海、甘肃、新疆的菌株(P<0.05);来源于内蒙古的菌株与青海、甘肃、新疆、西藏的菌株降解能力差异不显著(P>0.05)。不同地区发酵乳品由于海拔、温度、气候、季节以及牧民的生活习惯等因素使其存在着复杂多样的菌群微生态结构[26],具有特定的优势乳酸菌类群,优势菌群之间互相影响,使得不同发酵乳品中乳酸菌的丰度存在差异,从而可能导致乳酸菌降解亚硝酸盐特性之间的差异。乳酸菌的遗传多样性与其分离地之间也存在直接关系,遗传多样性使得乳酸菌表达出不同的性状,具体表现在其降解亚硝酸盐特性的差异。Song Yuqin等[27]利用多位点序列分型(multilocus sequence typing,MLST)技术对中国、俄罗斯及蒙古国等地的传统发酵乳品中L. delbrueckii ssp.bulgaricus进行微进化分析,研究发现每个地区乳酸菌都有属于其独特的基因型,证实了乳酸菌会因为适应环境而发生某种特定进化的假说,从而解释了乳酸菌降解亚硝酸盐特性因地区不同而存在差异的推论。
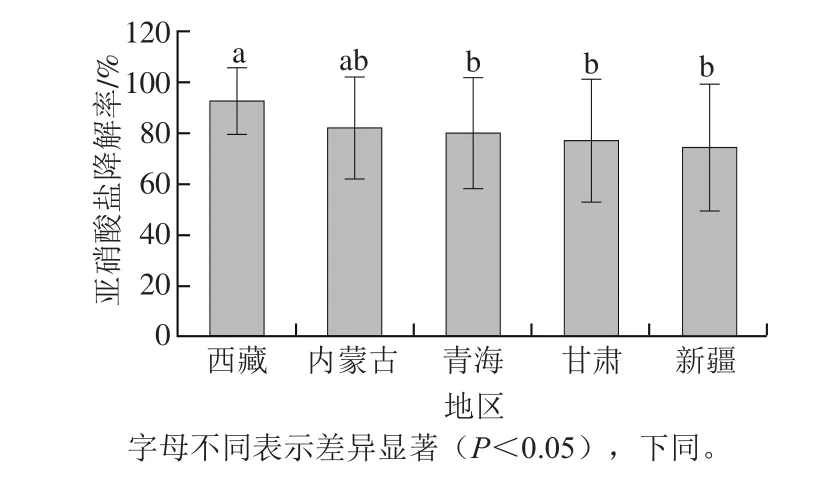
图1 不同地区乳酸菌亚硝酸盐降解率比较Fig.1 Comparison of nitrite degradation rate among LAB from different regions
2.2.4 不同发酵乳品中乳酸菌亚硝酸盐降解能力比较
将酸牦牛奶、酸马奶、酸驼奶、奶渣4 种发酵乳品中乳酸菌亚硝酸盐降解率进行方差分析,分析不同发酵乳品中乳酸菌在降解能力上的差异。如图2所示,不同发酵乳品中乳酸菌的降解能力存在显著性差异(P<0.05),来源于奶渣中的菌株降解能力显著高于酸牦牛奶和酸马奶中的菌株(P<0.05),而来源于酸驼奶中的菌株降解能力与酸牦牛奶、酸马奶、奶渣中的菌株差异不显著(P>0.05)。不同发酵乳品中乳酸菌降解能力存在差异,可能是由于牦牛、马、骆驼各物种之间以及各自生存条件的不同,导致其乳中干物质等营养成分存在差异,使得乳中乳酸菌群落结构和所表达出的降解亚硝酸盐特性不同。Sun Zhihong等[28]采用MLST技术对分离自我国和蒙古国的L. helveticus遗传多样性进行了系统分析,指出菌株的聚类与乳源有关,即酸马奶、酸牛奶和酸牦牛奶中分离出的乳酸菌形成单独的类群,且存在各自对应的祖先群体,显示出分离菌株为适应特定的生存环境而发生某种进化的历程。除此之外,乳的发酵前处理以及发酵乳的制作方式不同也会导致发酵乳中乳酸菌群落结构和特性的差异。

图2 不同发酵乳品中乳酸菌亚硝酸盐降解率比较Fig.2 Comparison of nitrite degradation rate among LAB from different fermented milk products
2.2.5 不同种类乳酸菌亚硝酸盐降解能力比较

图3 不同种类优势乳酸菌亚硝酸盐降解率比较Fig.3 Comparison of nitrite degradation rate among different dominant LAB
对8 种优势乳酸菌亚硝酸盐降解率进行方差分析,分析不同优势菌种在降解能力上的差异,如图3所示,8 种不同优势菌株之间降解能力存在显著性差异(P<0.05)。优势菌种中L. plantarum降解率最高,而L. fermentum降解率最低。通过分析可以发现L. breris、L. plantarum、L. paracasei、L. kef i ranofaciens、L. casei与L. fermentum、L. helveticus的降解能力相比有显著性差异(P<0.05);L. delbrueckii ssp. bulgaricus与L. breris、L. casei、L. plantarum、L. paracasei、L. kef i ranofaciens、L. fermentum的降解能力相比有显著性差异(P<0.05)。不同菌种在降解能力上的差异可能是由于不同种乳酸菌在降解亚硝酸盐基因水平上的不稳定,导致其表达亚硝酸盐降解能力的差异。菌种自身所具有的特性间可能存在相互影响,张庆芳等[29]认为,乳酸菌降解亚硝酸盐主要分为酶降解和酸降解两个阶段。乳酸菌发酵前期,pH值大于4.5时以酶降解为主,到了发酵后期pH值小于4.0时,以酸降解为主,故不同种乳酸菌产酸能力、产亚硝酸还原酶能力的不同也是导致其亚硝酸盐降解能力不同的原因之一。乳在发酵过程中硝酸还原菌可将硝酸盐转化成亚硝酸盐,导致发酵乳品中的亚硝酸盐含量超标,而乳酸菌具有抑制硝酸还原菌生长的作用,且不同种乳酸菌所具备抑制硝酸还原菌生长作用的差异较大,也可造成发酵乳中亚硝酸盐含量的不同[30]。
3 结 论
从青海、甘肃、新疆、内蒙古、西藏5地104 份传统发酵乳品中共分离得到275 株乳酸菌,鉴定出6 个属23 个种。通过比较分析发现不同地区、不同发酵乳品中存在着复杂多样的乳酸菌菌群微生态结构。其中L. helveticus为青海、甘肃、新疆、内蒙古、西藏5 个地区发酵乳品中共有菌株。L. helveticus和L. plantarum为酸牦牛奶、酸马奶、酸驼奶、奶渣4 种传统发酵乳品中共有菌株。以菌株筛分率为基准,青海发酵乳品中优势菌为L. helveticus和L. delbrueckii ssp. bulgaricus;新疆和甘肃发酵乳品中优势菌均为L. helveticus;内蒙古发酵乳品中优势菌为L. kefiranofaciens和L. helveticus;西藏发酵乳品中优势菌为L. breris、L. plantarum、L. casei和L. paracasei。酸牦牛奶中优势菌为L. helveticus和L. delbrueckii ssp. bulgaricus;酸马奶和酸驼奶中优势菌均为L. kefiranofaciens和L. helveticus;奶渣中优势菌为L. breris、L. plantarum、L. casei和L. paracasei。
对分离出的275 株乳酸菌亚硝酸盐降解能力进行分析,菌株降解率在4.8%~99.9%之间波动,50%的菌株表现出较好的降解能力,降解率平均在91.3%以上,其中8 株菌的降解率达到99.9%。
降解能力因菌株来源和乳酸菌种类不同存在差异性,来源于西藏的菌株降解能力显著高于青海、甘肃、新疆的菌株(P<0.05),来源于奶渣中的菌株降解能力显著高于酸牦牛奶和酸马奶中的菌株(P<0.05);优势菌种中L. plantarum、L. paracasei、L. breris显示了稳定高效的降解能力,且L. plantarum降解能力最强,其中L. breris、L. casei、L. plantarum、L. paracasei、L. kefiranofaciens与L. fermentum、L. helveticus、L.delbrueckii subsp. bulgaricus之间相互比较,部分菌株间存在显著性差异(P<0.05)。
猜你喜欢
食品安全导刊(2021年21期)2021-08-30
现代畜牧科技(2021年6期)2021-07-16
食品与发酵工业(2020年15期)2020-08-20
当代水产(2019年6期)2019-07-25
当代水产(2018年12期)2018-05-16
食品安全导刊(2018年27期)2018-01-17
作文周刊·八年级版(2017年21期)2017-07-10
恋爱婚姻家庭·青春(2016年7期)2016-07-05
中国民族医药杂志(2016年7期)2016-05-09
管理现代化(2016年6期)2016-01-23
