
肥水和清水下塘时泥鳅鱼苗的生长和存活
2018-07-24 12:55张云龙张子涵欧阳霞姚晓丽董昭然鲍传和
水产科学 2018年4期
张云龙,袁 娟,张子涵,欧阳霞,姚晓丽,董昭然,鲍传和
( 1.安徽农业大学 动物科技学院,安徽 合肥 230036; 2.肥东县水产技术推广站,安徽 合肥 231600 )

1 材料与方法
1.1 试验鱼
试验用鱼为同一批繁殖的4日龄仔鱼。
1.2 养殖条件及放养密度
泥鳅苗种培育池为8 m×4 m的水泥池。放苗前7 d以5 kg漂白粉化浆泼洒清塘。清塘之后注入新水,水深50 cm,以适量生石灰调节水体pH至约7.5,放养密度2500尾/m2,即每个水泥池放养泥鳅4日龄仔鱼80 000尾[初始体质量(1.28±0.37) mg,初始体长(3.97±0.41) mm)],养殖水温25~28 ℃。
1.3 试验设计
本试验分两种不同的培育模式,分为A组和B组,每个模式设置3个平行。模式A为肥水下塘,放苗前4 d用发酵菜粕、氨基酸生物肥和磷酸一铵按质量比2∶1∶1组合施肥,即发酵菜粕150 kg/hm2,氨基酸生物肥和磷酸一铵各75 kg/hm2。之后再根据饵料生物量,每隔3~5 d追肥一次,保证浮游动物量不低于10 个/mL。模式B为清水下塘,即放养泥鳅仔鱼的水泥池中不施肥,保持水质清澈。另外配备培养饵料生物的水泥池(饵料池),肥水方式同模式A。每日8:00和17:00从饵料生物池中以40目的自制浮游生物网捞取饵料生物,将捞取的饵料以60目的双层网片过滤3次以去除大个体饵料生物和杂质,过滤后投喂至模式B试验池中,每日早晚2次以浮游动物定量法检测培育池中饵料生物密度,保证放苗池中饵料生物的密度持续不低于10 个/mL。
1.4 养殖管理
试验池水深50 cm,水温25~28 ℃,自然光照,备有增氧泵,在溶解氧不足时增氧,以确保溶解氧不低于5 mg/L。定期监测水温、溶解氧及氨氮含量。A组水泥池中氨氮质量浓度为0.3~0.8 mg/L,施肥之后的1~2 d内氨氮含量较高。模式B的池中氨氮质量浓度低于0.2 mg/L。养殖期间,若中午气温过高,阳光较为强烈,则在水泥池上盖上凉纱,覆盖面积约为水泥池面积的33.3%。B组池如遇水质变差、浑浊时适量换水,保持水质良好、清澈。
1.5 取样
试验开始第2 d(5日龄)、7 d(10日龄)、12 d(15日龄)、17 d(20日龄)、22 d(25日龄)、27 d (30日龄)和32 d(35日龄)的早晨(7:00),随机从每个水泥池中捞出30尾泥鳅仔鱼,用游标卡尺测量全长(L),万分之一分析天平测量体质量(m),测量后未死亡的个体放回原池中。试验结束时,即第32 d,每个水泥池清池计算泥鳅数量,计数偏差不超过10%。同时计算下列参数:
质量增加率/%=(m2-m1)/m1×100%
增长率/%=(L2-L1)/L1×100%
特定生长率/%·d-1=(lnm2-lnm1)/(t2-t1)×100%
相对生长率/%·d-1=(eg-1)×100%,g=(lnm2-lnm1)/(t2-t1)
体质量变异系数变化率/%=(CVm2-CVm1)/CVm1×100%
全长变异系数变化率/%=(CVL2-CVL1)/CVL1×100%
成活率/%=Nt/N×100%
1.6 数据分析
试验数据均用平均值±标准差表示,用Microsoft Excel进行初处理,SPSS 18.0进行独立样本t检验比较两组之间均值的差异,显著性水平为α=0.05,Origin 9.0作图。
2 结 果
2.1 生长特点
本试验共取样1260尾泥鳅,测量体质量和全长,得到泥鳅体质量与全长的幂函数关系式:m=0.0674L2.2644(r2=0.9888),符合鱼类体质量、体长的一般关系式m=aLb,拟合曲线见图1。全长、体质量与日龄之间均呈指数函数关系,分别为L=1.8208 e0.0878t(r2=0.9471)和m=0.1257 e0.2219t(r2=0.8962),拟合曲线见图2。

图1 泥鳅全长与体质量的关系
2.2 生长和生长离散
两种培育模式下泥鳅全长和体质量的变化见图3。试验期间,A组与B组的全长和体质量之间差异不显著 (P>0.05),A组的终末全长和终末体质量均大于B组。
A组和B组的质量增加率、增长率、特定生长率及相对生长率均在25~30日龄之间达到最大,总体趋势均为先升后降,再升后再降,表现出一定的波动性,另一个极大值出现在10~15日龄之间(表1)。两组的各生长指标之间无显著性差异(P>0.05),说明两种培育模式对其生长并无显著的影响。
两种培育模式下,全长变异系数和体质量变异系数总体上呈缓慢下降趋势,略有波动。A组和B组各指标变化无明显的规律,全长变异系数和体质量变异系数仅在5日龄和20日龄差异显著(P<0.05),随着日龄的增加,泥鳅的变异系数并未发生显著变化(表2)。
试验期间,两组泥鳅的变异系数变化率均有较大的波动,其中负值占了大多数,说明其离散程度略有降低。A组的全长变异系数变化率和体质量变异系数变化率最大值分别出现在30~35日龄和20~25日龄。B组全长变异系数变化率和体质量变异系数变化率最大值均出现在25~30日龄(表3)。
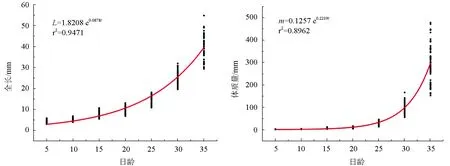
图2 泥鳅全长和体质量与日龄的关系

图3 两种模式下泥鳅全长和体质量的变化(平均值±标准差,n=90)

项目组别孵出日龄5~1010~1515~2020~2525~3030~35质量增加率%A88.87±35.28172.37±24.0474.05±29.29157.80±33.51298.07±50.73199.43±28.45B56.25±22.53167.16±13.5674.66±10.24180.30±5.95315.57±64.98178.93±22.20增长率%A27.65±5.4054.29±5.1025.79±6.1047.80±3.4069.01±5.9561.21±8.22B19.54±7.0148.37±2.3125.67±4.5650.84±1.0276.37±13.9252.00±7.05特定生长率%/dA12.49±3.6219.99±1.7310.88±3.5718.83±2.5827.52±2.6121.87±1.94B8.79±2.9919.64±1.0111.13±1.1520.61±0.4328.33±3.0720.47±1.61相对生长率%/dA13.36±4.1322.14±2.1211.54±3.9420.75±3.1231.71±3.4224.46±2.40B9.21±3.2421.70±1.2411.78±1.2922.89±0.5232.79±4.1022.73±1.91
注:表中结果为平均值±标准差,n=3.
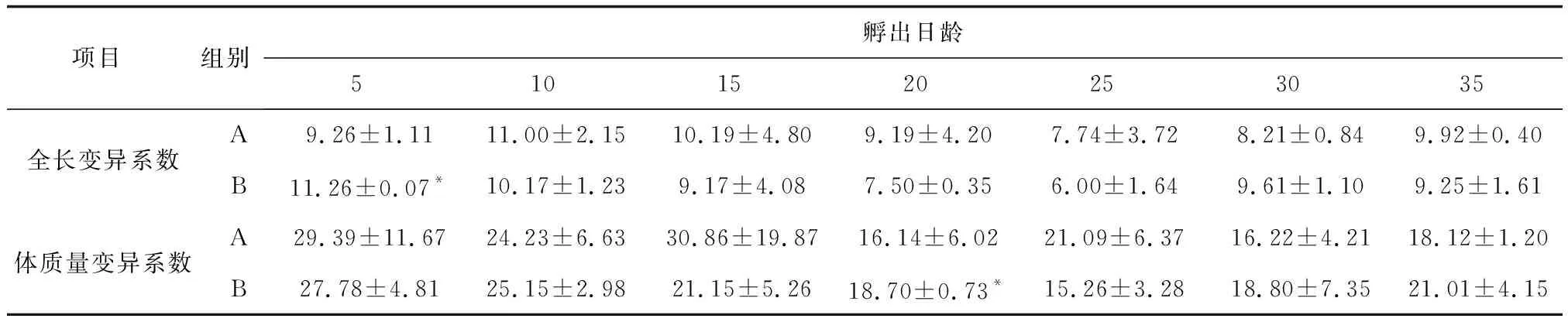
表2 不同培育模式下泥鳅的变异系数 %
注:表中结果为平均值±标准差,n=3,*为组间差异显著(P<0.05),下同.

表3 不同培育模式下泥鳅的变异系数变化率 %
2.3 成活率
由表4可见,两种培育模式显著影响泥鳅仔鱼的成活率(P<0.05),B组显著高于A组(P<0.05)。

表4 培育模式对泥鳅成活率的影响
3 讨 论
3.1 泥鳅的生长特点
泥鳅仔鱼体质量与全长之间呈显著的幂函数关系,这在鱼类中较为普遍,如斑鳍白姑鱼(Pennahiapawak)[7]、大鳞副泥鳅(Paramisgurnusdabryanus)[8]及双须叶须鱼(Ptychobarbusdipogon)[9]等。泥鳅的b值为2.2644,与多数鱼类为2.5~4.0不同[10],可能与泥鳅的体型有关。雷逢玉等[11]得到不同年龄段泥鳅体质量与体长关系式的b值为2.4561,金燮理等[12]得到隔年泥鳅体质量与体长关系式的b值为1.026,说明不同种群和不同年龄都影响b值。
泥鳅全长和体质量与日龄之间呈指数关系,并非匀速生长。泥鳅全长一直有较快的增长速度,而体质量在25日龄之前增加较慢,25日龄后则增加很快。这说明泥鳅仔鱼在25日龄前全长的增长较快,25日龄体质量增加较快。全长与日龄呈指数关系在菊黄东方鲀(Takifuguflavidus)仔稚鱼[13]中也得到了证明,但也有较多的研究中全长与日龄呈现直线关系,如刀鲚(Coilianasus)[14]仔稚鱼等。可见不同鱼类及同种鱼类不同时期的生长模式不尽相同。
3.2 培育模式对泥鳅仔、稚鱼生长和生长离散的影响
仔、稚鱼类一般生长较快,但其生长又易受外界条件的影响。放养密度[15]、驯食饵料[16-19]、分级[20]、再次分级[21]、延迟投喂[22]、联合投喂[23]等因素均可影响鱼类生长。本研究中,肥水下塘和清水下塘两组泥鳅仔稚鱼的生长未表现出显著差异,主要原因应该是两种模式的初始放养密度和饵料密度、种类差异不大。因此,单就生长而言,这两种模式之间并不存在优劣之分。虽然15日龄之后肥水下塘组的全长和体质量要高于清水下塘组,但其质量增加率、增长率、特定生长率和相对生长率4项指标与清水下塘组差异不显著,说明这两组泥鳅仔稚鱼的生长速度无差异。肥水下塘组死亡率较高,即随日龄的增加,肥水下塘组的密度低于清水下塘组,这应是肥水下塘组全长和体质量较高的主要原因。高密度养殖会降低鱼类对能量的摄入[24-25],造成鳍和鳃等组织损伤[26]。值得注意的是,肥水下塘、清水下塘两组的质量增加率、增长率、特定生长率及相对生长率都在15~20日龄这段时间出现了明显的低值,说明泥鳅在15~20日龄之间存在一个生长的低谷期。与此相对应,这一阶段泥鳅仔稚鱼的胰蛋白酶活性也较低[27]。这是否为导致泥鳅15~20日龄的生长低谷期的因素还需进一步研究。
生长离散广泛存在于鱼类种群中,这是鱼类种群对不利摄食条件的一种应答[28]。内在条件、饵料及环境因素等[29-31]均可引起生长离散,培育方法如驯食[32]等也影响生长离散。两种培育模式下,泥鳅仔、稚鱼均具有较高的体长变异系数和体质量变异系数,说明泥鳅的生长离散并非培育模式所致。饵料对生长离散的影响很大,仔稚鱼摄食差异将直接导致生长离散。研究表明,饵料密度过高或者过低均会影响鱼类的生长和生长离散[33-37]。本研究中,饵料密度与多数研究中的仔稚鱼最适饵料密度相差不大,如大西洋鲱(Clupeaharengus)[38]仔鱼在饵料密度为3.0个/mL时生长较好;花尾胡椒鲷(Plectorhinchuscinctus)[39]仔鱼最适饵料密度为30 个/mL。由此可推断,本研究中泥鳅仔稚鱼生长离散的原因并不是饵料密度。试验期间,肥水下塘组和清水下塘组的养殖密度随着泥鳅仔稚鱼死亡率的增加而降低。与之相对应,两组鱼的生长离散程度均有所下降,但差异并不显著。因此,本研究中养殖密度也不是造成生长离散的主要原因。泥鳅存在天然多倍体,本研究中不排除有多倍体的存在,多倍体与正常二倍体的生长速度差异的研究还未见报道。多数鱼类存在两性生长差异,如尼罗罗非鱼(Oreochromisniloticus)、鳜鱼(Sinipercachautsi)及蓝鳃太阳鱼(Lepomismacrochirus)等[40]。泥鳅存在两性异形[41],但其是否存在两性生长差异尚未见报道。在实践中观察到雌性泥鳅个体较大,推测泥鳅存在两性生长差异则能充分解释泥鳅仔稚鱼期间较高程度的生长离散。
生长离散一定程度上提升了残食的可能性,如斜纹犬牙石首鱼(C.regalis)[42]、鳡鱼(Elopichthysbambusa)[21]等肉食性鱼类。有关泥鳅残食的研究未见报道,与泥鳅同科的大鳞副泥鳅存在残食现象[43]。本试验中也观测到一例泥鳅残食的现象。对同类相残现象应深入研究,以探讨其是否对泥鳅苗种的成活率造成一定的影响。
3.3 泥鳅早期死亡原因
鱼类普遍存在早期死亡率较高的现象,如美洲鳀鱼(Engraulismordax)[44]、五条(Seriolaquinqueradiata)[45]等。本研究中,两种养殖模式显著影响了泥鳅苗种的成活率,清水下塘组[(54.85±6.09)%]显著高于肥水下塘组[(15.68±3.36)%]。仔稚鱼的免疫系统尚未完善,病原微生物对其生存的威胁较大,如黄颡鱼(Pelteobagrusfulvidraco)[46]。本研究中也未发现,肥水下塘组的泥鳅仔稚鱼存在病理特征。饵料、饥饿及消化系统发育也是影响仔稚鱼存活率的重要因素[47-49],但本研究中肥水下塘组和清水下塘组均保持足量的饵料供应。过量的氨氮虽然对鱼类有明显的毒性作用[50-52],但泥鳅具有极高的氨耐受性[53-56]。由此可说明并非上述因素导致肥水下塘组泥鳅苗种的高死亡率。
肥水下塘组和清水下塘组的水源相同,唯一的区别在于肥水下塘组进行了施肥,水质较差,水体富含氮、磷等有机物。鱼鳃易受到水质变化的影响[57],如果这种变化影响了鳃的功能对鱼类来说是致命的[58]。Evans[59]认为,鱼鳃与人类的肠道、肾脏相似,是鱼类处理水环境污染物的主要场所,说明水质对泥鳅的鳃组织有一定的影响。泥鳅孵化后尚不具备内鳃,只具有与内鳃同源的外鳃[60],通常在15日龄后才完全消失[61]。外鳃由若干长条状的鳃丝组成,实践中观察到外鳃丝十分脆弱,容易折损。肥水下塘组水质较差,生物量大、氮磷含量高,对泥鳅仔鱼的鳃丝造成一定的损害,导致了肥水下塘组的成活率显著低于清水下塘组的成活率。
4 结 论
本研究设计了两种泥鳅苗种培育模式,传统的肥水下塘和清水下塘。经过32 d的养殖后,肥水下塘和清水下塘两组泥鳅仔稚鱼均具有较高的生长速度和较大的生长离散程度,两者差异不显著;清水下塘组的成活率显著高于肥水下塘组,肥水下塘组泥鳅仔、稚鱼死亡率高的可能原因是水质恶化引起的泥鳅仔鱼外鳃折损。
猜你喜欢
作文周刊·小学一年级版(2022年16期)2022-05-07
水产科学(2022年1期)2022-01-26
渔业科学进展(2021年6期)2022-01-04
当代水产(2021年4期)2021-07-20
浙江海洋大学学报(自然科学版)(2021年1期)2021-07-06
南方水产科学(2021年3期)2021-06-23
当代水产(2020年4期)2020-06-16
当代水产(2020年4期)2020-06-16
当代水产(2018年8期)2018-11-02
小学生导刊(2018年22期)2018-08-21
