
毛新杨×银灰杨杂种花粉母细胞减数分裂及花粉形态1)
2018-07-20 12:14:12田菊牛新月铁英林晓飞
东北林业大学学报 2018年7期
田菊 牛新月 铁英 林晓飞
(内蒙古和盛生态科技研究院有限公司,呼和浩特,011517) (内蒙古大学)
杨树是我国华北、东北和西北地区用材林、防护林、碳汇林建设的"当家"树种[1],迄今已栽植约500万hm2,但相关区域杨树良种选育进展缓慢,目前在用材林、防护林建设和城镇绿化中,仍采用20世纪中期选育出的群众杨(Populus‘Popularis’)、北京杨(P.pyramidalis×P.cathayana‘Beijingensis’)等杂交品种,以及尚未品种化的新疆杨(P.bolleana)、河北杨(P.hopeiensis)等树种,严重制约了生态建设中树种多样性的选择。杨树的基因组杂合性非常高,往往在减数分裂过程中存在着较为复杂的染色体行为。目前,杨属白杨派、黑杨派、青杨派中部分树种在减数分裂上研究较多,大多集中在减数分裂进程与染色体行为方面[2-4]。毛新杨×银灰杨二倍体杂种母本是耐寒性较强的毛新杨(P.tomentosa×P.bolleana),父本为更为耐寒的银灰杨(P.canescens),具有速生、耐盐碱、抗寒、抗旱、抗病虫害等特点,适宜于内蒙古、宁夏、辽宁和吉林南部平原地区栽培。本研究为了较好地利用毛新杨×银灰杨杂种的优良基因资源,为后期杂交育种、多倍体育种以及分子育种等提供必要的遗传学信息,主要开展了花芽外部形态、花粉形态的观测,并利用醋酸洋红临时压片法对小孢子母细胞减数分裂进程及其染色体行为进行了系统地研究。
1 材料与方法
2017年2月份,于内蒙古自治区通辽市奈曼旗兴隆沼林场附近(43°4′5″N,120°49′5″E),采集花芽饱满且无病虫害的毛新杨×银灰杨杂种雄株花枝,用厚塑料薄膜包裹严实快递至内蒙古和盛生态科技研究院进行切枝水培,室内温度保持在20~25 ℃,每2~3 d换一次水。为了保证每个减数分裂时期均能被观察并固定,对枝条进行了多批次多时间段水培。
花序形态观察: 从材料水培时起,每隔1 d观察一次,每次固定观察30个花芽。记录花芽的长度和宽度(1/2位置为标准数值),并选择具有代表性的花芽拍照。
减数分裂观察: 水培开始后,每隔2~4 h采集3~5个雄花芽,去掉苞片,置于卡诺固定液(V(无水乙醇)∶V(冰醋酸)=3∶1)中固定2~24 h后,再放在4 ℃条件下保存,一直固定到花药成熟散粉时为止。减数分裂观察时,采用康向阳等[2]的方法进行,即夹取卡诺固定液中花芽的小花,置于洁净载玻片上,除去花盘,滴加2滴醋酸洋红,用尖嘴镊子夹碎花药制作涂片,火焰干燥,晾干后,利用Olympus BX-51光学显微镜观察,Olympus DP70照相系统数码拍照。
花粉母细胞减数分裂特异性统计 : 在减数分裂观察过程中,进行特异性统计,每个时期观察500个以上的花粉母细胞,根据统计结果进行减数分裂特异行为比率计算。
花粉大小及败育率统计: 花序散粉时,利用干燥的毛笔将花粉收集到放有硅胶的西林瓶中,盖上盖子置于冰箱中4 ℃备用。花粉大小观察测定时,滴一滴醋酸洋红在洁净的载玻片上,用镊子夹取少量花粉,均匀涂抹在醋酸洋红中,盖上盖玻片,消除气泡,染色5 min后置于光学显微镜下观察,利用Olympus DP70照相系统进行随机拍照(视野不重复),观察花粉粒总数为1 000个以上,利用Adobe Photoshop CS3对花粉直径、败育花粉率、天然2n花粉率进行统计。以花粉直径大于平均直径的1.28倍作为未进行减数的2n花粉[3]。
数据统计与图片处理:采用Microsoft Excel 2010软件进行数据管理、统计分析与图表绘制,采用Adobe Photoshop CS3进行图片制作。
2 结果与分析
2.1 毛新杨×银灰杨杂种雄株的花序发育进程
通过观察发现,毛新杨×银灰杨杂种雄株花序发育进程分为花蕾期(图1 a),这时花序长度约为1 cm,花芽膨大,花序未露出苞片;伸展期(图1 b、图1c),花序长度快速增加,并露出苞片,每一朵小花逐渐变大,颜色逐渐由浅红变成深红,由极度紧缩慢慢变伸展;盛花期(图1 d),这时花序长度仍在伸长,每一朵小花均逐渐开裂散粉,一般是花序基部最先散粉,然后是中部,顶端最后散粉;开花末期(图1 e),每朵小花均开裂散粉,散粉完毕后,花盘干枯,花序开始逐渐脱落。
在室内20~25 ℃水培条件下,毛新杨×银灰杨杂种花期为7 d,其花序长度变化范围为1.00~5.20 cm,呈“S”型增长,在3~4 d ,生长速度加快,第4 d的长度是第3 d的1.36倍,5 d后长度趋于稳定,7 d之后花序脱落;宽度变化范围为0.74~1.00 cm,3 d后宽度趋于稳定(表1)。通过方差分析表明,每天的花序长度和宽度之间均差异极显著(sig≤0.001),说明在花序发育过程中,其长度和宽度均有一定程度的增加,但长度的增加量更加明显。

a.花蕾期(0~1 d);b、c.伸展期(2~3 d);d.盛花期(4 d);e.开花末期(5~7 d);标尺均为1 cm。

图1 毛新杨×银灰杨杂种花序发育进程
注:表中数据为平均值±标准差;同列不同字母表示差异显著(P<0.05)。
2.2 毛新杨×银灰杨杂种雄株花粉母细胞减数分裂进程及染色体行为
毛新杨×银灰杨杂种减数分裂进程在花蕾期完成。室温20~25 ℃条件下,完成减数分裂大概需要39 h。水培至14 h时,开始进行减数分裂,花粉母细胞进入细线期(图2a),细胞核变大,核仁十分明显,染色质逐渐盘旋折叠,变短变粗,并出现极化现象,向核仁一侧聚拢。经过28 h后,进入粗线期(图2b),染色体明显缩短、变粗,颜色加深,此时出现了同源染色体配对现象。水培至33 h后,减数分裂进入双线期,可明显观察到,核仁颜色变浅,开始消失,联会的两条同源染色体开始分离,但某些区段仍黏结在一起,形成交叉结(图2c)。水培35 h后,进入终变期(图2d),染色体变得非常收缩,更短更粗达到最小体积,此时,核仁完全消失。终变期的结束就标志着减数分裂前期Ⅰ的完成。之后,减数分裂进入一个较快的变化阶段。中期Ⅰ时(图2e),纺锤体出现,在其牵引下,染色体排列在赤道板上,此时,出现了一些形态不同的单价体和二价体。后期Ⅰ(图2f),同源染色体逐渐向两级移动。末期Ⅰ(图2g),在花粉母细胞两极各出现1个核仁,染色体呈染色质状,并开始聚集。末期Ⅰ完成后,标志着第一次减数分裂完成,进入第二次减数分裂。前期Ⅱ(图2h),表现为染色体逐渐变粗短,核仁第2次开始消失。中期Ⅱ(图2i),染色体显著缩短,整齐地排列在赤道板上。后期Ⅱ(图2j),着丝点分裂为2个,染色单体在纺锤丝牵引下分别移向两极。末期Ⅱ(图2k),移向两极的染色体开始聚集,开始解螺旋化,恢复到染色质的形态,同时4个核仁形成。随着两次的细胞质分裂,花粉母细胞形成4个子细胞,即“四分体”(图2l)。

a.细线期;b.粗线期;c.双线期;d.终变期;e.中期Ⅰ;f.后期Ⅰ;g.末期Ⅰ;h.前期Ⅱ;i.中期Ⅱ;j.后期Ⅱ;k.末期Ⅱ;l.四分体;标尺均为20 μm。
2.3 毛新杨×银灰杨杂种雄株花粉母细胞减数分裂异常行为
毛新杨×银灰杨杂种在减数分裂过程中除了正常的分裂行为外,还会出现一些异常分裂现象(表2)。中期Ⅰ时,一些染色体不排列在赤道板上,而分散在赤道板外,这些异常类型比率达17.27%。在后期Ⅰ时,由于一些染色体分离速度较慢,导致了落后染色体和染色体桥出现,概率分别为7.99%和1.86%。末期Ⅰ时,占总细胞17.17%比率的微核出现。中期Ⅱ,纺锤体一般情况下是互呈60°或120°(图3a),而在毛新杨×银灰杨杂种中出现了其他4种纺锤体,它们分别是融合纺锤体(图3b)、平行纺锤体(图3c)、垂直纺锤体(图3d)以及多个纺锤体,出现概率分别为6.10%、8.01%、8.89%、0.35%,其中以平行纺锤体和垂直纺锤体比率较高。后期Ⅱ时,同样有落后染色体和染色体桥出现,但比后期Ⅰ概率低,分别为3.55%和0.39%。末期Ⅱ,微核的出现率也较末期Ⅰ低,为7.84%。在四分体时期,除了正常形态的四分体外(77.96%)(图3 e),还形成了其他较多种类的减数分裂产物,总变异率可达22.04%,其中单分体(图3 f)、二分体(图3 g)、三分体(图3 h)、多分体的概率分别为2.89%、9.19%、2.13%、0.26%,含微核的单分体、二分体(图3 i)、三分体(图3 j)、四分体(图3 k)的概率分别为0.94%、4.77%、0.94%、0.94%,在这几种特殊"分体"中,属二分体出现概率高,三分体和多分体出现的概率较低。此外,在单分体中,还存在着细胞内只有1个细胞核(图3 f)、2个细胞核(图3 l)以及细胞质断裂(图3 m)的异常情况;在二分体中,出现了细胞核分裂成2个(图3 g)、3个(即4个细胞核中有2个融合未分离)(图3 n)的特异情况;在二分体和四分体中,还出现了多个微核(图3 i和图3o)。
2.4 毛新杨×银灰杨杂种雄株花粉特性
在植物减数分裂过程中,由于减数分裂异常情况的出现,会使花粉大小不一。对毛新杨×银灰杨杂种花粉直径测量后(共测1 278粒),发现其花粉直径变化明显,呈高斯分布,变化范围为15.07~54.61 μm,平均为30.20 μm。花粉直径大小集中分布在28~34 μm ,该直径范围内花粉数占所有被测量花粉数的86.56%。

表2 毛新杨×银灰杨杂种减数分裂异常情况比率
通过对毛新杨×银灰杨杂种的花粉特性研究表明,其存在花粉败育情况(图3 p,箭头1所示),败育率为9.47%,正常花粉率为90.53%(表2),具有较高的可育性。根据前人的研究表明,花粉直径在平均直径1.28倍(38.67 μm)以上为未进行减数的2n花粉,因此,本研究中毛新杨×银灰杨杂种也存在大花粉(图3 p,箭头2所示),但出现概率较低,为1.41%。
3 结论与讨论
温度是影响植物生长发育的主要环境因子。温度越高,植物的生长速度越快。毛白杨在自然状态下,完成减数分裂时间为7~8 d,而在温度为10~25 ℃的环境下,其完成减数分裂所需时间为1~3 d[2]。王君等[4]研究表明,通辽杨在10~20 ℃水培条件下,约7 d完成减数分裂。本研究中,毛新杨×银灰杨杂种在室温20~25 ℃情况下,只需39 h即可完成减数分裂。由此表明,减数分裂进程主要是受植物自身特性和外部温度进行调控的。
有关研究表明,杂种起源的植物种,因为其染色体来源于不同的物种,会使染色体配对不规则及染色体分离不平衡,往往会表现出减数分裂异质性[5-6]。杨树品种在减数分裂过程中存在着广泛的染色体异常现象,往往出现落后染色体、染色体桥、融合纺锤体、单分体、三分体、带微核的分体等[2,4-6]。本研究中,毛新杨×银灰杨杂种是由杂种三倍体毛新杨和杂种二倍体银灰杨进行双杂交获得的,融合了毛白杨、新疆杨、银白杨、山杨4个亲本的染色体,杂合程度非常高,因此在其小孢子母细胞减数分裂过程中存在着较为复杂的染色体配对现象,从而导致了上速染色体异常现象的发生。中期Ⅰ和末期Ⅰ时异常细胞比率分别为17.27%和17.17%,两个时期变异率十分接近,说明末期Ⅰ微核的形成可能与中期Ⅰ染色体不在一条直线上有较大关联。
减数分裂过程中,一般认为,融合纺锤体和平行纺锤体可使二分体产生,而三分体的产生可能与三极纺锤体有关[5]。毛新杨×银灰杨杂种平行纺锤体和融合纺锤体出现的概率之和为14.11%,而二分体和含微核的二分体概率之和为13.96%,与前述结论一致。另外,本研究中还出现了较少的多个纺锤体,概率为0.35%,其与多分体(概率0.26%)的形成息息相关。此外,互呈一定角度的纺锤体与四分体形成直接相关,在各自分裂项中出现概率比较接近,分别为76.66%和78.90%。
花粉败育往往伴随着减数分裂异常现象的出现而出现。毛新杨×银灰杨杂种败育花粉率为9.47%,微小孢子出现率总和为7.59%,两者差异较小。常童洁等[7]指出,BM-1毛白杨不育雄株在花药发育过程中,由于脂类营养物质匮乏和代谢的紊乱引发绒毡层非正常死亡,使花粉无法及时得到营养供给,最终导致花粉败育。而笔者却认为植物花粉败育是因为减数分裂过程中微小孢子体内由于没有整套完整的遗传物质。
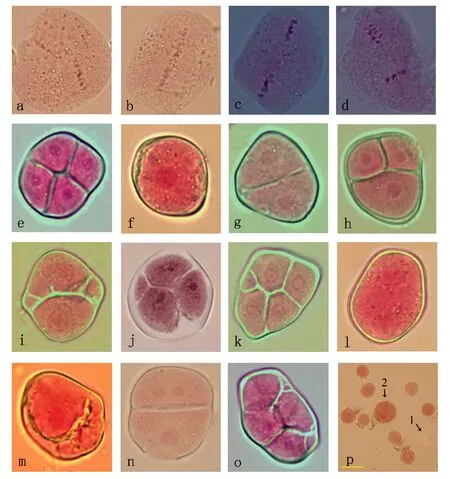
A.60°或120°纺锤体;b.融合纺锤体;c.平行纺锤体;d.垂直纺锤体;e.四分体;f.含1个细胞核的单分体;g.含2个细胞核的二分体;h.三分体;i.含有多个微核的二分体;j.含1个微核的三分体;k.含1个微核的四分体;l.含2个细胞核的单分体;m.细胞质分裂的单分体;n.含3个细胞核的二分体;o.含多个微核的四分体;p.花粉形态;箭头1示败育花粉粒;箭头2示大花粉粒;其他为正常花粉粒;标尺为50 μm。
图3毛新杨×银灰杨杂种中期Ⅱ和四分体时期特异情况及花粉形态
植物界中普遍存在2n花粉,通常单倍性花粉比2n花粉要少一套染色体[3,8]。本研究中天然2n花粉率为1.41%,概率较低。有关研究推测天然2n花粉的发生可能与后期Ⅱ纺锤体定位异常有关[9-10],而Zhang et al.[11]研究指出,第二次减数分裂过程中平行纺锤体的极点融合是毛白杨天然2n花粉发生的主要原因。本研究结果揭示了毛新杨×银灰杨双杂交种减数分裂的遗传变异规律,可为后期倍性育种、诱变育种以及分子育种等研究提供较好的技术资料。
猜你喜欢
美文(2024年6期)2024-03-08 02:56:33
中国医学科学院学报(2022年1期)2022-12-30 13:30:21
鸭绿江(2021年29期)2021-11-11 14:48:26
诊断学(理论与实践)(2020年2期)2020-06-28 12:41:12
生物学教学(2017年9期)2017-08-20 13:22:32
癌变·畸变·突变(2016年5期)2016-08-22 05:55:18
天津农业科学(2015年12期)2015-12-03 23:52:47
癌变·畸变·突变(2015年4期)2015-02-27 06:15:22
作文评点报·作文素材小学版(2014年46期)2014-11-25 13:03:47
作文评点报·作文素材小学版(2014年17期)2014-06-24 11:55:47
