
诱变育种提高嗜盐四联球菌精氨酸和瓜氨酸利用能力
2018-07-18 09:39:30雷庆子王博堵国成方芳
食品与发酵工业 2018年6期
雷庆子,王博,堵国成,方芳*
1(江南大学 生物工程学院,江苏 无锡,214122)2(工业生物技术教育部重点实验室,江苏 无锡,214122 ) 3(糖化学与生物技术教育部重点实验室,江苏 无锡,214122 )
氨基甲酸乙酯(ethyl carbamate,EC)是一种存在于发酵食品中的2A类致癌物,具有遗传毒性,可导致肺肿瘤、淋巴癌、肝癌和皮肤癌[1-2]。EC曾被用作抗肿瘤药物,由于具有致癌性,现已停止临床使用[3]。摄入体内的EC经细胞色素P450的羟基化作用和氧化作用生成可损伤DNA的8-羟基-脱氧鸟苷和乙烯基-氨基-甲酸酯环氧化物,通过破坏DNA结构引发癌变[4-6]。
发酵食品中形成EC的主要前体包括氰化物、氨甲酰磷酸、尿素和瓜氨酸[7]。酱油生产过程中容易积累有害物氨基甲酸乙酯,这种现象在高盐稀态法发酵生产酱油的过程中尤为显著[8-9]。高盐稀态酱油中,EC的主要前体是瓜氨酸,由乳酸菌通过精氨酸脱亚氨酶途径(arginine deiminase pathway,简称ADI途径)代谢精氨酸生成[10-14]。ADI途径包括3个关键酶:精氨酸脱亚氨酶(arginine deiminase, ADI)、鸟氨酸转氨甲酰酶(omithine transcarbamylase, OTC)和氨基甲酰激酶(carbamate kinase, CK),分别由arcA、arcB、arcC基因编码[15-16]。由于酱油发酵是一个混菌发酵的食品体系,生产上不允许使用基因工程菌或用基因工程手段改造生产用菌株。通过内源微生物干预是实现控制和减少酱油中EC及其前体的有效措施。
寻找来源于酱醪微生物体系并可利用精氨酸且不积累瓜氨酸的菌株,通过在发酵过程中强化这一菌株来降低酱油中的EC前体瓜氨酸,可为建立消除食品混菌发酵体系中的有害微生物代谢物的方法提供研究基础和理论支持。嗜盐四联球菌(Tetragenococcushalophilus)是存在于酱油、泰国鱼酱、果汁等高盐或高糖环境中的一类乳酸菌[17-19]。它具有提升食品风味、改善食品品质的功能[20-21]。已有研究表明,来源于酱醪的嗜盐四联球菌R23可利用酱醪中的精氨酸和瓜氨酸,从而减少EC前体,起到控制EC含量的作用[22]。然而,体系中葡萄糖和精氨酸含量对嗜盐四联球菌利用或积累瓜氨酸均有显著影响:当体系中葡萄糖和精氨酸含量增加时,嗜盐四联球菌利用精氨酸和瓜氨酸能力显著下降[22-23]。这说明,嗜盐四联球菌用于酱油发酵时,其精氨酸和瓜氨酸利用能力受环境因素制约。因此,通过诱变育种的方法增强嗜盐四联球菌利用精氨酸和瓜氨酸的能力,对于发展酱油产业、控制或降低酱油中EC含量、提高发酵食品的安全性具有重要意义。
1 材料和方法
1.1 菌株
嗜盐四联球菌(Tetragenococcushalophilus)R23,分离自高盐稀态酱油的酱醪,保藏于中国典型培养物保藏中心(保藏编号:CCTCCNo:M2013480)。
1.2 培养基
嗜盐四联球菌培养基(g/L):蛋白胨10,MnSO40.05,乙酸钠5,MgSO40.2,蔗糖10,牛肉膏5,柠檬酸三铵2,葡萄糖 10,吐温80 1,酵母粉4,K2HPO42,NaCl 100,pH 7.2。
初筛培养基(g/L):酵母粉5,吐温80 1 ,MgSO40.2 ,蛋白胨5 ,MnSO40.05 ,葡萄糖0.5,FeSO40.4,柠檬酸三胺2,牛肉膏5,CaCO30.1,NaCl 100 ,吡哆醛-5-磷酸0.05,K2HPO42,瓜氨酸0.5,精氨酸3~4,pH 5.5。
复筛培养基(g/L):酵母粉5,葡萄糖0.5,MgSO40.2,牛肉膏5,MnSO40.05, 蛋白胨5,FeSO40.4,柠檬酸三胺2,吐温80 1,CaCO30.1,吡哆醛-5-磷酸0.05,K2HPO42,NaCl 180,瓜氨酸0.5,精氨酸3~5,pH 5.5。
氨基酸利用培养基(g/L):酵母膏5,NaCl 180,葡萄糖0.5,吐温80 1,牛肉膏5,MgSO40.2,蛋白胨5,MnSO40.05,FeSO40.4,柠檬酸三胺 2,CaCO30.1,吡哆醛-5-磷酸0.05,K2HPO42,瓜氨酸 0.5,精氨酸5,pH 5.5。
1.3 主要试剂
色谱纯甲醇、瓜氨酸、色谱纯乙腈、分析纯三氯乙酸、鸟氨酸、氨基酸标准样品、四氢呋喃、精氨酸购自上海生工生物工程股份有限公司。用于二乙酰一肟-氨基硫脲比色法显色反应的试剂如文献所述[24-25]。培养细胞/细菌总RNA提取试剂盒(DP430),反转录试剂盒(FastQuant RT Kit (With gDNase)(KR106))购自天根生化科技(北京)有限公司。氨测定试剂盒(100M6204)购自SIGMA公司。
1.4 诱变方法
将嗜盐四联球菌R23培养至对数生长期,离心收集菌体,用磷酸盐缓冲液(pH 7.0)洗涤3次后悬浮菌体,分别进行紫外诱变和等离子诱变。
(1)紫外诱变
将菌液置于15 W紫外灯下30 cm处进行紫外诱变,诱变时间分别为0、20、30、60、90、100、120 s。照射后菌液置于避光处静置培养30 min,避光稀释涂平板,于30 ℃恒温避光培养3 d。根据诱变致死率绘制诱变致死率曲线,选择致死率在90%左右的诱变条件,确定为:距15 W紫外灯30 cm处、照射2 min。
(2)等离子诱变
取10 μL的菌悬液滴加在无菌载玻片上,用常压室温等离子体(ARTP)进行等离子诱变,在功率100 W,通气量为10 SLM条件下分别诱变0、1、3、5、7、10 s,然后转移至含1 mL生理盐水EP管中,将样品梯度稀释后涂布于嗜盐四联球菌培养基平板上,30 ℃培养3 d。根据存活率绘制诱变致死率曲线,选择致死率在95%左右的诱变剂量,确定诱变条件为:在功率100 W、通气量为10 SLM条件下照射3 s。
1.5 突变株筛选
(1)高通量转接培养:挑取突变株单菌落,接种至分别含有1 mL,3、3.5、4 g/L精氨酸初筛培养基的96深孔板中,30 ℃培养5 d。
(2)初筛:利用二乙酰一肟-氨基硫脲比色法测定瓜氨酸含量[26],进行诱变菌株初筛,利用酶标仪测定溶液530 nm处吸光值(OD530),以未进行诱变育种的嗜盐四联球菌R23的培养基比色测定吸光值作为对照组,选择吸光值更小的突变株进行复筛。
(3)复筛:将初筛得到的菌株接种至有1 mL复筛培养基的深96孔板中,30 ℃静置培养5 d,离心,利用二乙酰一肟-氨基硫脲比色法分别测得在530 nm处吸光值(OD530),以嗜盐四联球菌R23的培养基比色测定吸光值作为对照,选择吸光值较其更小的突变株作为复筛菌株,并利用高效液相色谱法(HPLC)测定复筛菌株培养体系中氨基酸含量,得到优势突变株。
1.6 氨基酸测定
利用HPLC测定体系中游离氨基酸的含量[26]。测定时取 1 mL菌液,10 000 r/min离心5 min,取上清液。用质量分数为5%的三氯乙酸将其稀释5倍,用滤膜(0.22 μm)过滤稀释液至液相瓶中,用HPLC测定滤液中瓜氨酸、鸟氨酸和精氨酸含量。测定条件为:色谱柱 ODS-2 HYPERSIL(250 mm×4.6 mm,5 μm),流速:1 mL/min。检测器为 VWD 紫外检测器,柱温 40 ℃,检测波长为 338 nm,分离时间 38 min[10]。流动相 A (1 L):无水乙酸钠 5 g,超纯水 1 L,三乙胺 200 μL,四氢呋喃5 mL, pH 7.2。流动相 B (1 L):无水乙酸钠 5 g,超纯水200 mL,甲醇 400 mL,乙腈 400 mL, pH 7.2。
1.7 环境因素对突变株利用精氨酸和瓜氨酸能力的影响
将突变株和野生菌株R23接种至嗜盐四联球菌培养基,30 ℃静置培养3 d,离心收集菌体并用磷酸盐缓冲液(pH7.0)洗涤。考察葡萄糖浓度、温度以及酱油乙醇发酵时期环境因素对突变株利用精氨酸和瓜氨酸能力的影响时,菌体再用相应氨基酸利用培养基垂悬,在30 ℃(考察温度影响在设定温度下培养)静置培养5 d,测定精氨酸和瓜氨酸的消耗量。
1.8 酶活的测定方法
粗酶液的制备:取30 mL对数生长后期嗜盐四联球菌菌液离心收集菌体,用磷酸盐缓冲液(pH 7.0)洗涤后加入等体积的氨基酸利用培养基,30 ℃培养5 h,4 500 r/min离心 5 min 收集菌体。用磷酸盐缓冲液(pH 7.0)洗涤并垂悬菌体,破壁取上清即得粗酶液。
(1)精氨酸脱亚胺酶(ADI)酶活的测定
在5 mL的EP管中加入150 μL的50 mmol/L精氨酸、2.3 mL的50 mmol/L磷酸盐缓冲液(pH7.0)和50 μL粗酶液,37 ℃水浴30 min后用5%的三氯乙酸终止反应,对照用50 mmol/L磷酸盐缓冲液(pH7.0)代替粗酶液。用高效液相色谱法测定反应液中瓜氨酸含量,计算ADI酶活(ADI 酶活为稀释倍数乘以单位时间内转化生成的瓜氨酸含量)。
(2)鸟氨酸转氨酶(OTC)酶活的测定
在5 mL的EP管中加入160 μL的 50 mmol/L磷酸盐缓冲液(pH 7.0)、40 μL的25 mmol/L鸟氨酸溶液、150 μL的133 mmol/L 氨甲酰磷酸溶液、240 μL的粗酶液和600 μL的超纯水,37 ℃水浴15 min后用5%的三氯乙酸终止反应,对照用50 mmol/L磷酸盐缓冲液(pH7.0)代替粗酶液。用高效液相色谱法测定反应液中瓜氨酸含量,计算 OTC 酶活(OTC 酶活为稀释倍数乘以单位时间内转化生成的瓜氨酸含量)。
(3)氨基甲酸酯激酶(CK)酶活的测定
在1.5 mL的EP管中加入160 μL 的50 mmol/L磷酸盐缓冲液(pH 7.0)、200 μL的73 mmol/L的MgCl2溶液以及200 μL的50 mmol/L ADP,混匀后放置 10 min,加入200 μL的133 mmol/L氨甲酰磷酸溶液,混匀后继续放置 10 min,向混合液中加入100 μL粗酶液,37 ℃水浴15 min后用5%的三氯乙酸终止反应,对照用50 mmol/L磷酸盐缓冲液(pH7.0)代替粗酶液。将反应液稀释一定倍数后,按照氨测定试剂盒(SIGMA公司)中的测定方法测定CK酶活(CK酶活为稀释倍数乘以单位时间内转化生成的NH3含量)。
1.9 实时定量PCR法分析基因转录水平
将嗜盐四联球菌培养至对数生长期,离心收集菌体,用磷酸盐缓冲液(pH 7.0)洗涤3遍后分别垂悬于含4 g/L和5 g/L精氨酸的氨基酸利用培养基中,30 ℃培养4 h。低速离心获得菌体,与2 mL RNA 稳定剂混合,-80 ℃保藏。采用液氮研磨的方式对上述获得的菌体进行破壁。用培养细胞/细菌总RNA提取试剂盒(天根生化科技(北京)有限公司)提取RNA并用反转录试剂盒(FastQuant RT Kit (With gDNase) (天根生化科技(北京)有限公司)制备cDNA。qRT-PCR使用的引物如表1所示,选择16S rRNA作为内参基因。使用2-ΔΔCt的方法进行转录水平分析[27]。
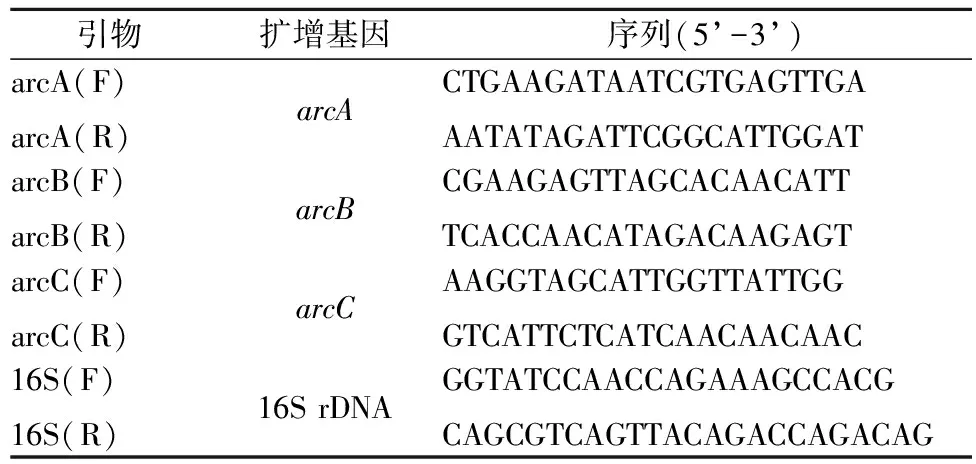
表1 用于实时定量PCR的引物Table 1 Primers for qRT-PCR
2 结果和分析
2.1 嗜盐四联球菌诱变育种及高效利用瓜氨酸突变株筛选
嗜盐四联球菌R23在精氨酸质量浓度低于3 g/L时可利用酱醪中的精氨酸且不积累EC前体瓜氨酸,但是当精氨酸含量高于3 g/L时,体系中瓜氨酸开始积累[22]。为获得精氨酸和瓜氨酸利用能力提高的菌株,通过诱变嗜盐四联球菌R23,共得到紫外诱变突变株386株,等离子诱变突变株237株。采用二乙酰-肟-氨基硫脲比色法确定瓜氨酸相对含量的筛选方法,用于突变株的初筛(图1)[25]。
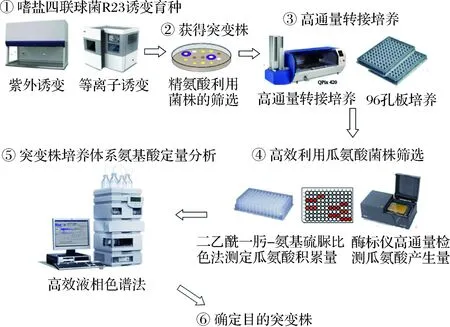
图1 嗜盐四联球菌诱变及突变株筛选Fig.1 Mutagenesis of Tetragenococcus halophilus and screening of mutants
通过初筛和复筛,获得了4株在精氨酸质量浓度高于3 g/L时,瓜氨酸积累少于出发菌株R23的突变株,分别命名为1-A2,1-A10,5-H1和3-H9(图2)。
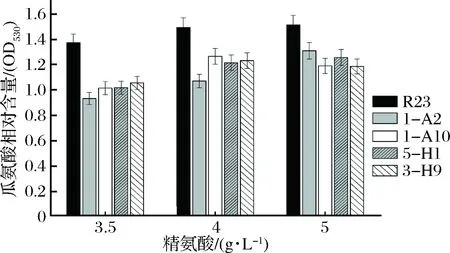
图2 嗜盐四联球菌利用精氨酸和积累瓜氨酸能力分析Fig.2 Conversion of citrulline from arginine by T. halophilus 1-A2,1-A10,5-H1 and 3-H9,mutants of T. halophilus R23
2.2 嗜盐四联球菌突变株利用精氨酸和瓜氨酸的能力
为进一步确定突变株利用精氨酸和瓜氨酸的能力,以及寻找具有有效降低酱油中EC含量潜力的菌株,分别对4株嗜盐四联球菌突变株利用精氨酸和瓜氨酸的能力进行了分析和比较(图3)。
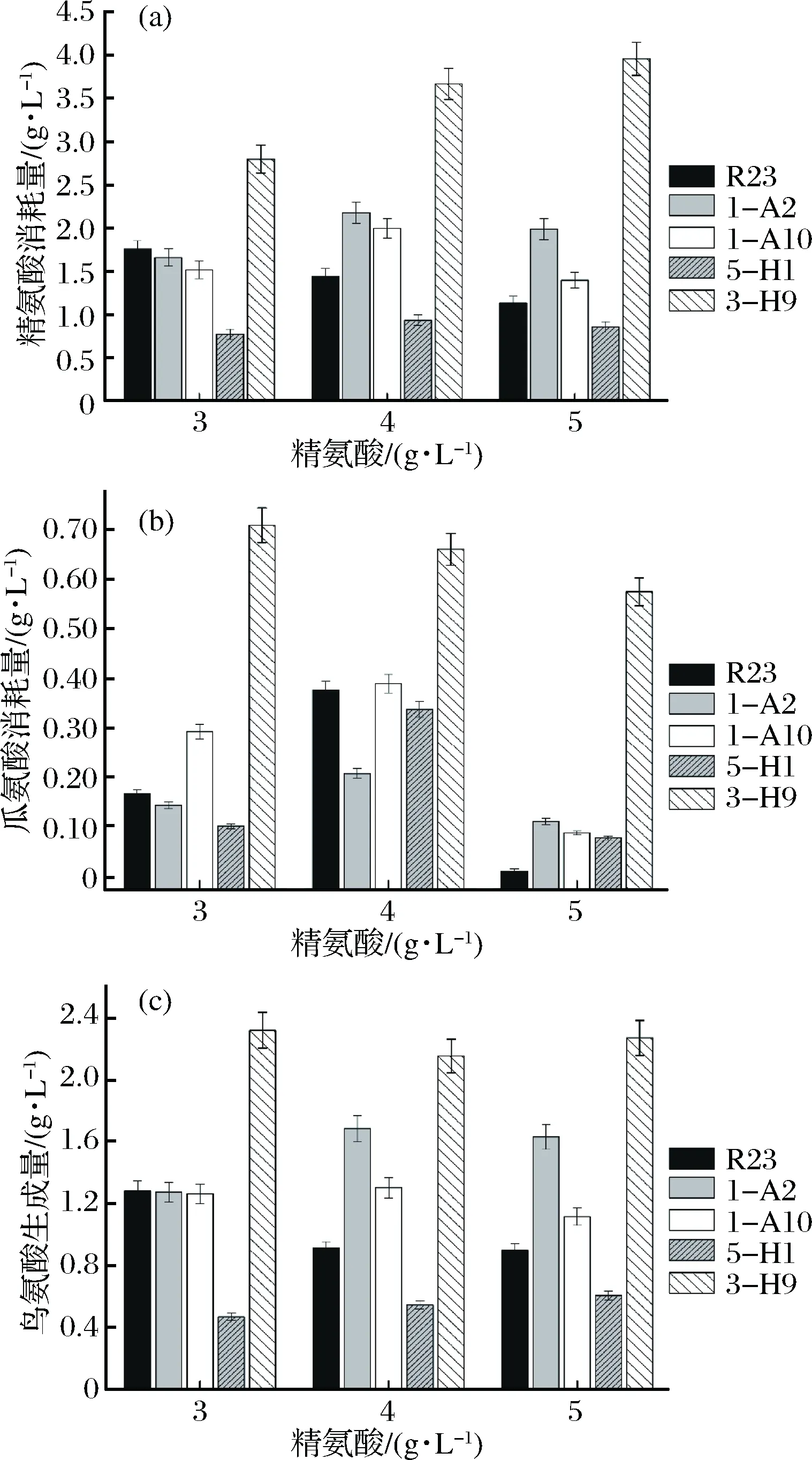
图3 嗜盐四联球菌利用精氨酸和瓜氨酸的能力比较Fig.3 Utilization of arginine and citrulline by T. halophilus 1-A2,1-A10,5-H1 and 3-H9,mutants of T. halophilus R23
由图3可以看出,当体系中含有0.5 g/L瓜氨酸和3、4、5 g/L精氨酸时,突变株3-H9利用精氨酸能力最高,比R23提高了0.6~2.49倍。突变株3-H9利用瓜氨酸能力也比R23提高了0.74~56倍。突变株5-H1精氨酸利用能力下降,瓜氨酸利用能力没有显著提高。突变株1-H10和1-A2利用精氨酸的能力比R23略有提高,但对瓜氨酸的利用能力远低于突变株3-H9。因此,选择突变株3-H9进行后续研究。
2.3 环境因素对突变株利用精氨酸和瓜氨酸能力的影响
ADI途径是细菌获得能量的一条辅助代谢途径。当培养体系中还原糖充足时菌株优先利用还原糖供能,因此ADI途径代谢通量会受到还原糖的抑制[10]。由于乙醇和游离脂肪酸的协同作用,细菌通过ADI途径代谢精氨酸时,会积累瓜氨酸,对EC前体含量的有效控制时期为酱油发酵前期,即乳酸发酵时期[28]。在酱油发酵前期(1~7 d),酱醪中葡萄糖的质量浓度为0~7 g/L[29]。通过考察葡萄糖浓度对突变株利用精氨酸、瓜氨酸能力的影响发现,葡萄糖质量浓度大于2 g/L时野生菌株R23已不能消耗精氨酸,而突变株3-H9在葡萄糖质量浓度为7 g/L时仍能利用精氨酸。突变株3-H9利用精氨酸的能力在葡萄糖质量浓度为2、3、5、7 g/L时,分别比R23提高 5.05、5.62、3.41、1.22 g/L(表2)。这说明,突变株3-H9精氨酸代谢受葡萄糖抑制作用被减弱,具有进一步减控酱油中EC前体的潜力。
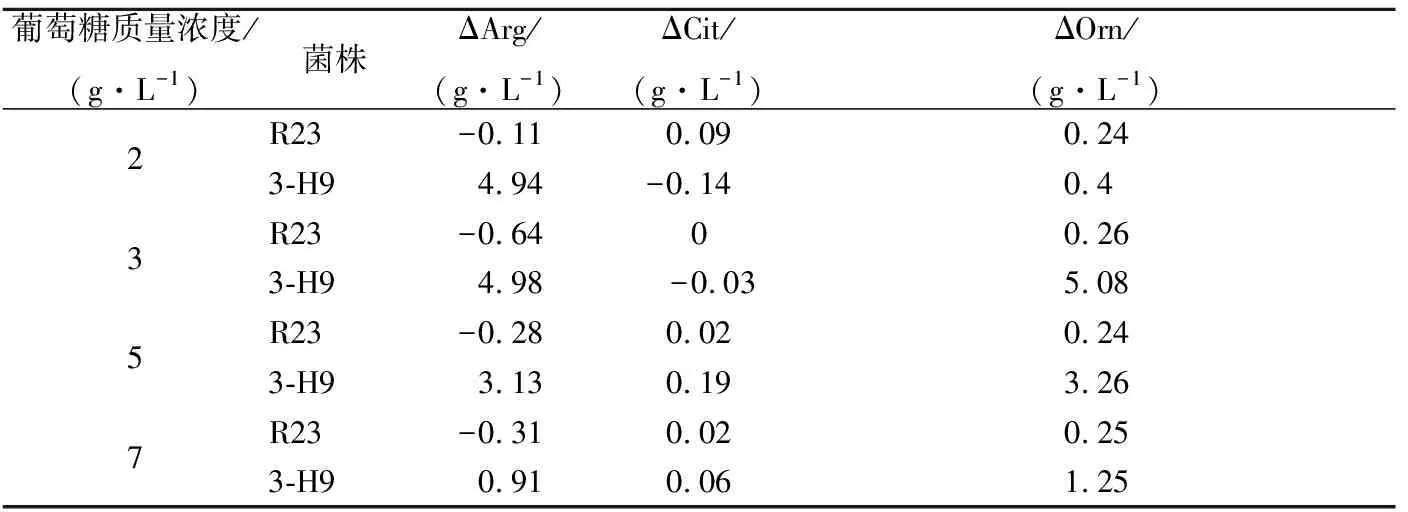
表2 葡萄糖对嗜盐四联球菌利用精氨酸和瓜氨酸能力的影响Table 2 Effects of glucose concentration on arginine and citrulline utilization of T. halophilus mutants
注: ΔArg为精氨酸消耗量;ΔCit为瓜氨酸消耗量; ΔOrn为鸟氨酸生成量。
高盐稀态酱油发酵过程前期温度较低,为了研究温度对嗜盐四联球菌突变株利用精氨酸能力的影响,考察了突变株3-H9在20、25、30 ℃条件下利用精氨酸能力。由表3可知,温度低于30 ℃时,突变株3-H9和野生菌株R23利用瓜氨酸和生成鸟氨酸的能力均有所减弱。较低温培养条件下,突变株3-H9利用精氨酸的能力高于野生菌株R23。在20、25 ℃培养时,3-H9利用精氨酸能力分别比R23提高57.7%和47.8%(图4)。

表3 温度对突变株利用瓜氨酸能力的影响Table 3 Effects of temperature on the use of citrulline in mutant strains
注:ΔCit为瓜氨酸消耗量(g/L); ΔOrn为鸟氨酸生成量(g/L)。
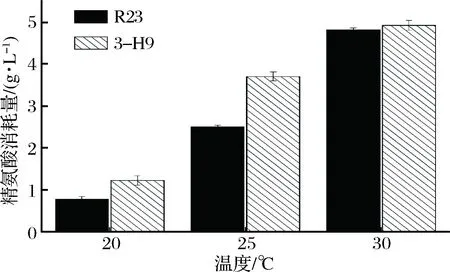
图4 温度对嗜盐四联球菌利用精氨酸的影响Fig.4 Effect of temperature on arginine utilization of T. halophilus
酱油发酵过程中,瓜氨酸的积累有2个时期:一是乳酸发酵时期,占主导地位的微生物是乳酸菌(主要为魏斯氏菌属和足球菌属的菌株),二是乙醇发酵时期,葡萄球菌属、微球菌属和芽孢杆菌属是优势菌群[28,30-31]。乙醇发酵时期酱醪中含有的乙醇和游离脂肪酸是导致细菌利用精氨酸积累瓜氨酸的主要因素。因此,我们考察了突变株3-H9在含有2%的乙醇与0.25%游离脂肪酸(模拟酱油乙醇发酵时期)的培养体系中利用精氨酸的情况。由表4可知,在此条件下,野生菌株R23不利用精氨酸,而突变株3-H9仍可利用精氨酸。由于突变株3-H9在这一培养条件下利用精氨酸积累了瓜氨酸,因此它更适用于在酱油乳酸发酵时期发挥作用。

表4 乙醇和游离脂肪酸对嗜盐四联球菌利用精氨酸和瓜氨酸能力的影响Table 4 Effect of ethanol and fatty acids on arginine and citrulline utilization of T. halophilus
注: ΔArg为精氨酸消耗量;ΔCit为瓜氨酸消耗量; ΔOrn为鸟氨酸生成量。
2.4 突变株瓜氨酸代谢相关基因转录水平及酶活分析
细菌对精氨酸的利用以及瓜氨酸的生成和消耗与ADI途径中3种关键酶:精氨酸脱亚胺酶(ADI)、鸟氨酸转氨酶(OTC)和鸟氨酸氨基甲酸激酶(CK)密切相关。由表5结果可知,突变株3-H9较R23相比,ADI酶、OTC酶和CK酶活性分别提高了27%、38%和0.4%,说明突变株3-H9利用精氨酸和瓜氨酸的能力都比出发菌株高。

表5 突变株及野生菌株中 ADI、OTC、CK酶酶活分析Table 5 Activity of ADI,OTC,CK in mutant and wild strains
ADI酶、OTC酶和CK酶是分别由arcA、arcB和arcC基因编码。为了探明突变株3-H9利用精氨酸和瓜氨酸能力及调控水平变化的原因,对突变株3-H9和野生菌株R23arc基因簇转录水平进行了比较分析。由图5可知,突变株3-H9中arcA、arcB和arcC转录水平分别比R23提高了7.9倍、9.5倍和7.8倍。在ADI途径中,arcA编码的ADI酶催化精氨酸向瓜氨酸转化,arcB基因编码的OTC酶催化瓜氨酸向鸟氨酸转化。通过比较arcA与arcB基因表达水平可以得知该条件下ADI途径产物是终产物鸟氨酸,还是含有中间产物瓜氨酸。此途径中,若arcA/arcB大于1,表现为瓜氨酸积累,arcA/arcB小于1则表现为瓜氨酸利用。通过对突变株3-H9和野生菌株R23的arcA/arcB转录水平之比分析发现:在4 g/L精氨酸浓度下,两株菌arcA/arcB比值均小于1,均表现为利用瓜氨酸;当精氨酸浓度为5 g/L时,野生菌arcA/arcB>1,表现为积累瓜氨酸,而突变株3-H9arcA/arcB<1,表现为利用瓜氨酸(表6)。

图5 ADI途径相关基因转录水平分析Fig.5 Transcriptional analysis of arc genes in T. halophilus
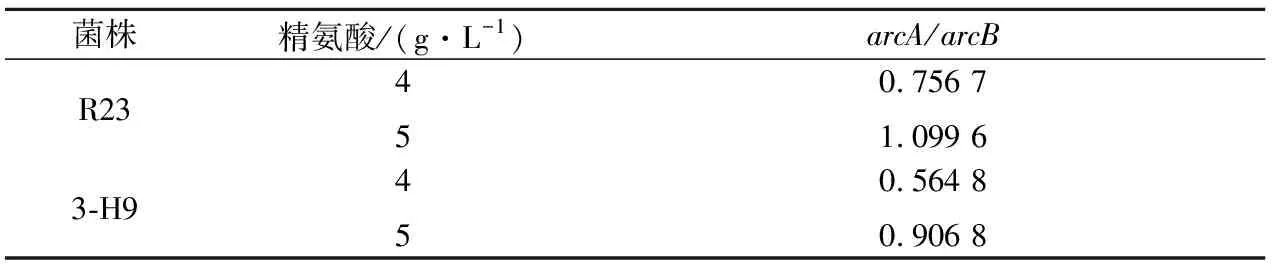
菌株精氨酸/(g·L-1)arcA/arcBR23450.756 71.099 63-H9450.564 80.906 8
3 结论
本研究以对酱油风味有强化作用的内源微生物嗜盐四联球菌为出发菌株,通过等离子和紫外诱变的方法获得了1株利用精氨酸和瓜氨酸能力显著提高的嗜盐四联球菌突变株3-H9。该突变株具有减少酱油中氨基甲酸乙酯前体瓜氨酸的应用潜力。研究结果提供了一种通过改造发酵食品体系中某一有益菌的氮源代谢方式来减少发酵食品中有害物的策略,可为控制或降低酱油中氨基甲酸乙酯及其前体提供理论依据,对于提高嗜盐四联球菌的工业应用性能以及发展酱油产业,提高食品的安全性具有有利意义。
猜你喜欢
临床肝胆病杂志(2017年3期)2017-03-07 06:44:51
现代检验医学杂志(2016年1期)2016-11-12 13:19:34
中国免疫学杂志(2016年2期)2016-01-30 21:18:21
西南军医(2016年3期)2016-01-23 02:17:56
中外医疗(2015年11期)2016-01-04 03:58:50
药学与临床研究(2015年4期)2015-06-05 11:35:53
中国当代医药(2015年30期)2015-03-01 02:08:13
疑难病杂志(2014年12期)2014-04-16 05:19:33
湖泊科学(2014年2期)2014-02-27 10:30:05
河南医学研究(2012年3期)2012-08-15 00:48:04
