
松褐天牛垂直分布规律
2018-07-07 01:20涂业苟喻爱林曾吉华王文辉刘亚军刘晓华
生物灾害科学 2018年2期
涂业苟 ,喻爱林 ,曾吉华 ,王文辉 ,刘亚军 ,刘晓华
(1. 江西省林业科学院,江西 南昌 330013;2. 南昌市白蚁防治研究所,江西 南昌 330039)
松褐天牛(Monochamus alternatusHope)是我国南方松林的重要蛀干害虫,在我国分布于西藏以东、辽宁以南,东至台湾、南至广东,国外分布于日本、朝鲜、越南、老挝等国;其不仅能直接危害寄主树造成死亡,也是松材线虫(Bursaphelenchus xylophilusNickle)最主要的传播媒介[1-3]。 目前有关学者集中在松褐天牛的生物学特性、发生规律和防控技术等方面做了大量研究[4-10], 本文通过利用松褐天牛产卵对衰弱木的趋性,研究马尾松上的松褐天牛垂直分布和分布特点,为了解其生物学特性、种群动态和预测预报具有重要意义,同时为探讨生物防治和注干输药防治松褐天牛的方法提供思路和试验证据,从而控制松材线虫病的迁移和扩散。
1 试验地点
试验地点位于江西省万安县沙坪镇,地理坐标为 114°48′31〞E,26°17′41〞N,平均海拔300 m。该试验区域为 15~25 年生马尾松天然次生林,林分郁闭度0.3~0.6,树高 2.5~11 m,胸径 5~15 cm,近年来受松褐天牛为害严重。
2 材料与方法
2.1 材料
2015 年 10 月,在试验地内随机选择胸径5~13 cm [平均胸径(9.09±2.08) cm],树高 2.7~8.3 m [平均树高(5.69±1.72) m]受松褐天牛严重危害的马尾松死树 47 株,离地面 10 cm处将其锯倒,测量树高、地径和胸径等指标。从基部开始每 1 m锯成一木段,一直到梢部直径为 3 cm处止,每株各木段按顺序编号,并记录每株每段树皮上的刻槽数量。刻槽数量为成虫产卵时在树干上刻槽的数量。
2.2 方法
2.2.1 松褐天牛刻槽、侵入孔和羽化孔数量与树干胸径的关系 调查马尾松死树 47 株,并放置于 1.5 m×1.5 m×1.5 m的纱网木制养虫笼内。待翌年8 月成虫完全羽化后,去皮逐株逐段调查统计幼虫侵入孔与成虫羽化孔的数量。侵入孔数量为幼虫进入木质部产生孔的数量,羽化孔数量为成虫羽化出来孔的数量。
2.2.2 松褐天牛在树干上不同高度的分布规律 选取其中树高 7 m 以上的马尾松死树 12 株,把树干高度按 1 m为 1 个米阶分级,即 0~1 m、1~2 m、2~3 m、3~4 m、4~5 m、5~6 m和 7 m以上,分为 7 级,统计松褐天牛在不同米阶上分布规律。
2.2.3 松褐天牛侵入孔、羽化孔数量与刻槽数量的关系 调查马尾松死树 47 株,根据马尾松上侵入孔、羽化孔的数量,分析其分布与刻槽数量的关系。
2.2.4 松褐天牛幼虫侵入孔和成虫羽化孔在树干上的垂直分布 调查马尾松死树47 株,幼 虫侵入孔和成虫羽化孔的调查方法同 2.2.1,统计幼虫侵入孔和成虫羽化孔在树干上的分布规律。
2.3 数据处理
试验所有数据用 SPSS17.0 软件进行处理[11], 并进行方差分析。通过线性回归、二次抛物线的方法,拟合刻槽、侵入孔和羽化孔与树干胸径的关系以及整株(或某米阶)松褐天牛刻槽与整株侵入孔、羽化孔数量的关系。对获得的数据采用聚集度指标进行侵入孔和羽化孔垂直分布测定及分布型指数分析,数据处理及计算方法主要参照文献[12-13]。
3 结果与分析
3.1 松褐天牛刻槽、侵入孔和羽化孔数量与树干胸径的关系
由图 1 可知,松褐天牛刻槽、侵入孔和羽化孔的数量与树干胸径呈正相关关系(n=47)。在胸径一定范围内,树干胸径越大,刻槽、侵入孔和羽化孔的数量也越多。根据调查数据计算得出松褐天牛刻槽、侵入孔和羽化孔的数量与树干胸径的关系模型分别为y=55.841x-273.11 、 y=35.252x-168.85 和y=26.252x-129.66,并 分别对其模型进行F检验,其 相关性均达到极显著水平(R2= 0.990 8;F=981.566;d f=1,46;P=0.000 1)、(R2= 0.989 9;F=338.335;df=1,46;P=0.000 1)和(R2= 0.978 2;F=418.095;df=1,46;P=0.000 1)。
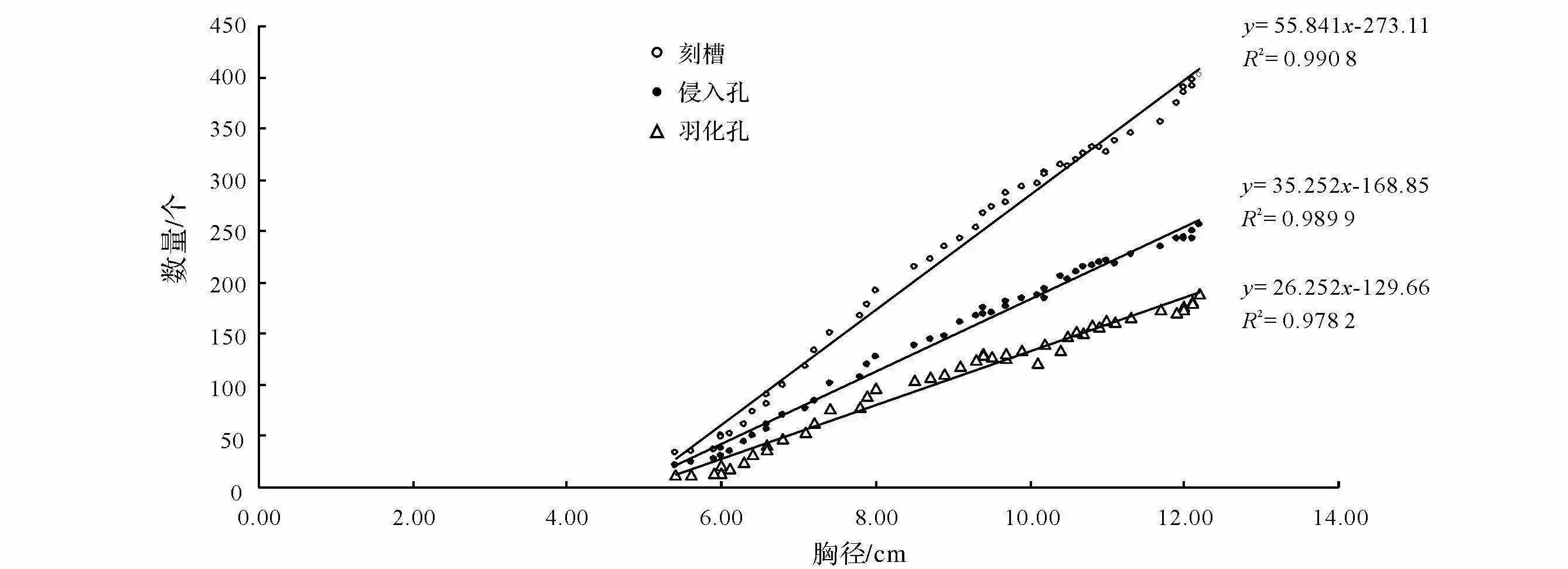
图1 松褐天牛刻槽、侵入孔和羽化孔与树干胸径的关系
3.2 松褐天牛在树干上不同米段的分布规律
由表 1 可知,松褐天牛刻槽、侵入孔和羽化孔数量在马尾松树干上不同米阶的分布有所不同,各米段之间的刻槽(F=419.341;df=7,88;P=0.000 1)、侵入孔(F=498.624;df=7,88;P=0.000 1)和羽化孔(F=225.006;df=7,88;P=0.000 1)数量均存在极显著差异。松褐天牛刻槽最集中的米段在 2~3 m,平均数量为 77.92 个,约占总刻槽数量的 21.38%;其次为3~4 m、1~2 m和 4~5 m,分别约占 18.16%、17.95%和 14.22%;松褐天牛的刻槽数量在马尾松树上主要集中在 1~5 m,占总刻槽数量的 71.71%。同样,松褐天牛侵入孔和羽化孔最集中的米段均在 2~3 m,平均数量分别为 56.08个、43.83 个;松褐天牛侵入孔和羽化孔的数量在马尾松树上主要集中在1~5 m,分别占总数的 75.20%、78.03%。以上结果表明,松褐天牛刻槽、侵入孔和羽化孔的数量在马尾松树上主要集中分布在1~5 m。
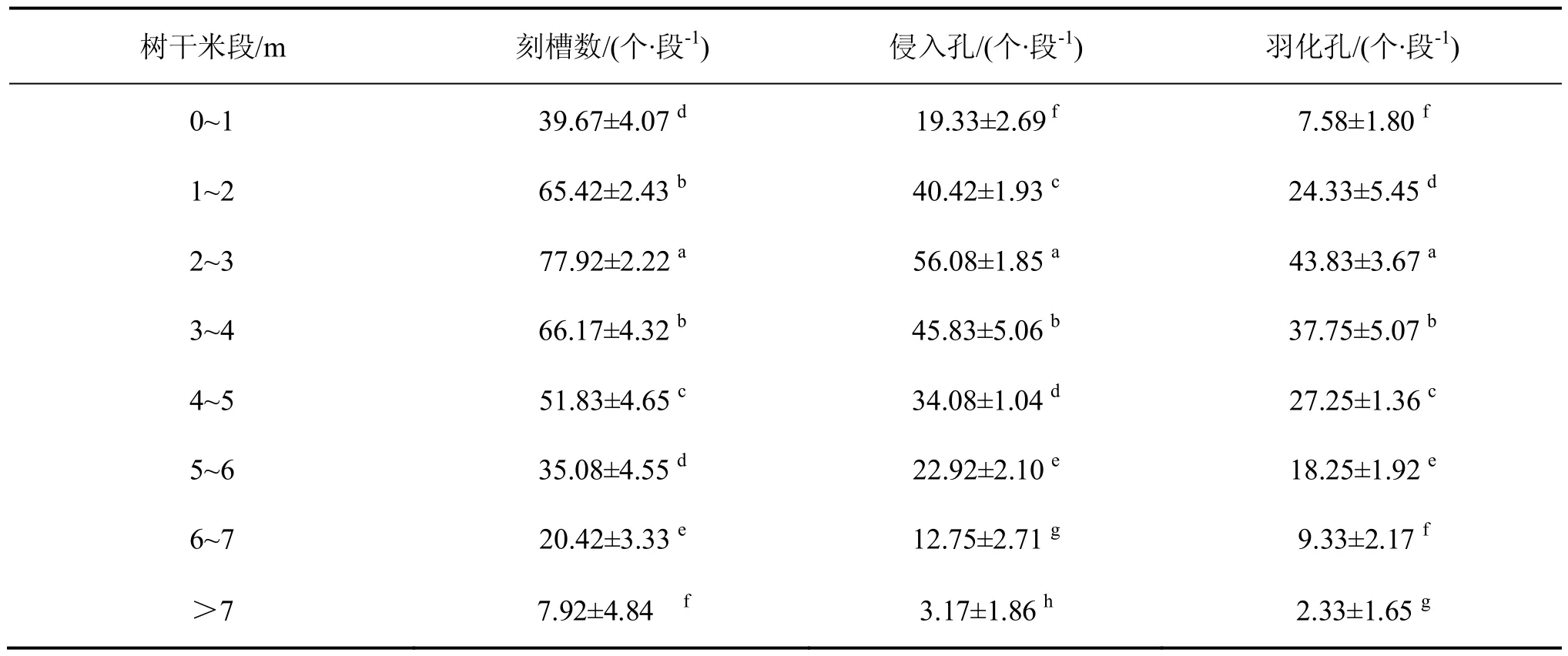
表1 松褐天牛刻槽、侵入孔和羽化孔在马尾松树干上不同米阶的分布规律
3.3 松褐天牛侵入孔、羽化孔数量与刻槽数量的关系
松褐天牛侵入孔、羽化孔的数量均与刻槽数量呈正相关关系,刻槽数量越多,幼虫侵入孔和成虫羽化孔也越多 (图2)。 根据调查数据计算得出松褐天牛侵入孔、 羽化孔的数量与刻槽的关系模型分别为y=0.630 4x+3.767 1、y=0.470 7x-1.412 9,并分别对其模型进行 F 检验,相关性均达到显著水平(R2= 0.996 4;F=1 271.289;df=1,46;P=0.022 3)、(R2= 0.989 9;F=6 421.481;df=1,46;P=0.009 9)。
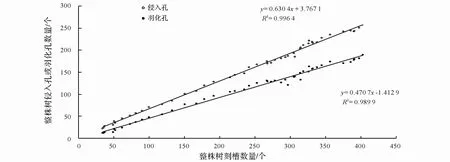
图2 整株树松褐天牛刻槽数量与侵入孔、羽化孔的关系
为了快速地掌握林间松褐天牛的虫口密度,也分析了整株侵入孔、羽化孔数量与树干高度 2~3 m刻槽数量的关系,结果表明整株马尾松上松褐天牛侵入孔、羽化孔数量与树干高度 2~3 m的刻槽数量存在明显相关,回归方程都表现为极显著关系(P<0.01)(图 3)。
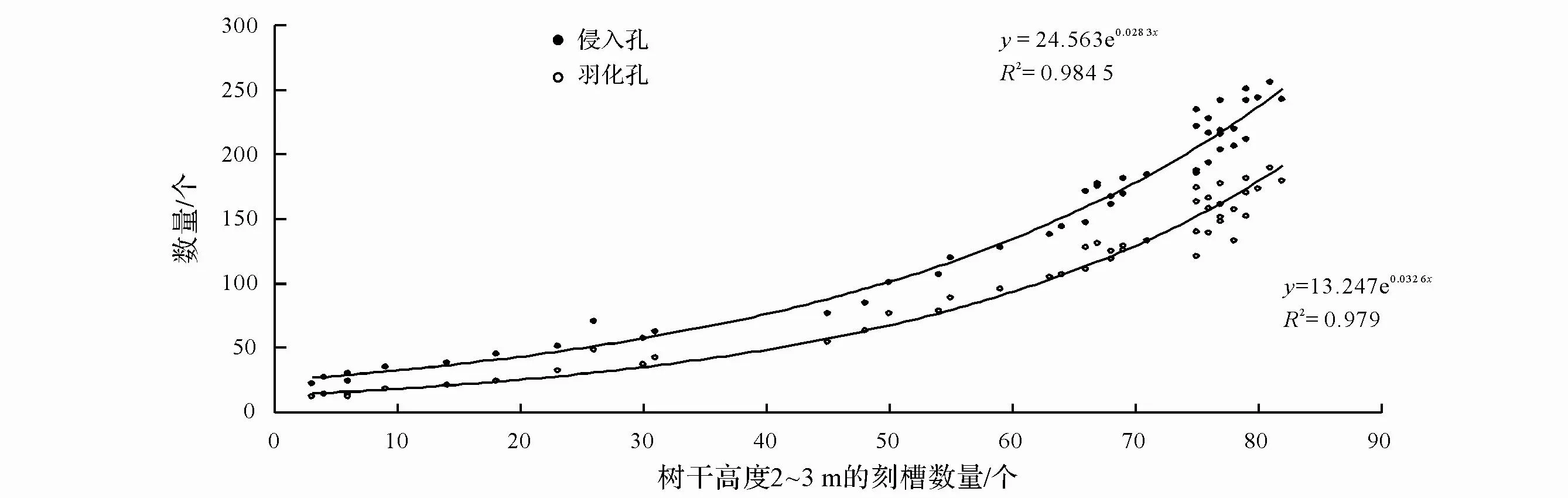
图3 整株树松褐天牛侵入孔、羽化孔数量与树干高度2~3 m刻槽数量的关系
3.4 松褐天牛幼虫侵入孔和成虫羽化孔的垂直分布
松褐天牛幼虫侵入孔的各项聚集指标见表 2。由表2中可知,C>1,I>1,Ca>0,K>0,M*/M>1,各项聚集指标均显示松褐天牛幼虫侵入孔的空间分布属聚集分布。通过M*-M回归分析法(Iwao),M*=1.013 6M+1.415 7;β=1.013 6,α=1.415 7(R2= 0.990 4,相关显著),α>0,分布的基本成分是个体相互吸引,β>1,为聚集负二项分布。另外根据 Talyor 幂法则,lg(ν)=0.002 7+1.259 6 lg(M),b>1,lgα>0(R2= 0.855 6,相关显著),结果说明松褐天牛幼虫种群在一切密度下都是聚集的,且聚集度随密度的升高而增加。同样,松褐天牛成虫羽化孔的各项聚集指标见表 2。由表2中可知,C>1,I>1,Ca>0,K>0,M*/M>1,各项聚集指标也均显示松褐天牛成虫羽化孔的空间分布属聚集分布。通过M*-M回归分析法(Iwao),M*=1.028 0M+1.013 1;β=1.028 0,α=1.013 1(R2= 0.991 0,相关显著),α>0,分布的基本成分是个体相互吸引,β>1,为聚集负二项分布。根据 Talyor 幂法则,lg(ν)=0.134 9+1.189 0lg(M),b>1,lgα>0(R2= 0.862 7,相关显著),结果说明松褐天牛羽化孔在一切密度下都是聚集的,且聚集度随密度的升高而增加。
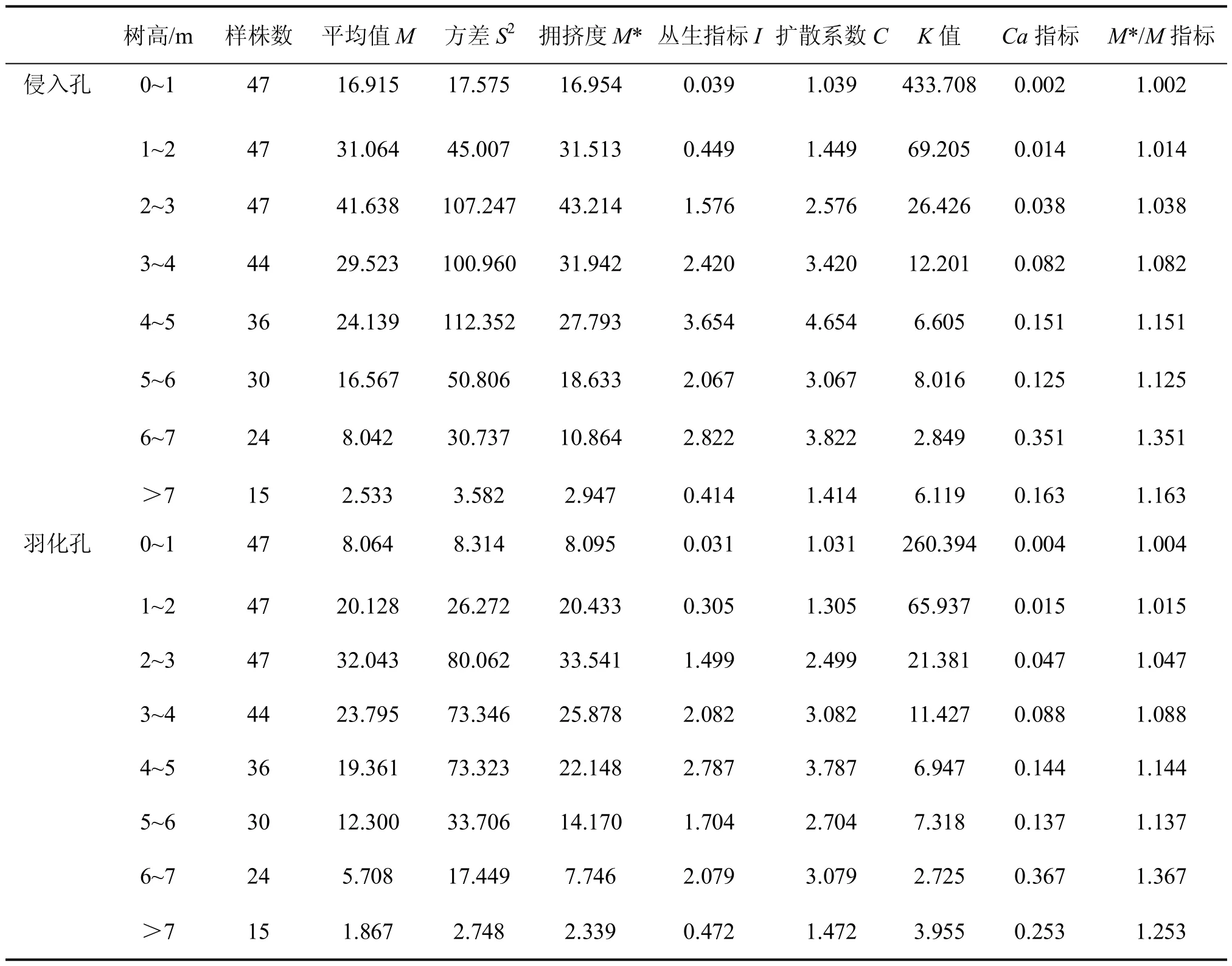
表2 松褐天牛幼虫侵入孔和成虫羽化孔的聚集度指标
4 讨 论
本研究发现,在一定胸径范围内,松褐天牛刻槽、侵入孔和羽化孔的数量与树干胸径的关系均呈正相关关系。胸径越大,刻槽、侵入孔和羽化孔的数量也越多。就单株而言,松褐天牛刻槽、侵入孔和羽化孔的数量在马尾松树上最集中的高度在 2~3 m;但主要集中分布在 1~5 m的地方,这部分高度的数量均占总数的 70%以上,这与孟俊国等[14]的 研究结论相一致。
根据垂直分布研究得出,马 尾松上松褐天牛的频次分布为聚集负二项分布,空 间分布型为聚集型分布,且密度越高,呈现出聚集度越大的趋势,分析松褐天牛的聚集原因是其自身和环境共同的作用结果[15]。
本文为生产实践提供了 1 种虫口密度的调查方法,与侵入孔、羽化孔的调查相比,刻槽的调查不需要剥树皮,方便快捷。根据整株(或树干高度 2~3 m)刻槽的数量,可以预测整株马尾松上的松褐天牛侵入孔和羽化孔总数量,更直观可靠。同时,在松褐天牛的防治上根据其刻槽数量进行生物防治(释放天敌)或注干输药防治,这样既节省人力、物力,又减少化学农药的环境污染,对有效防治松褐天牛具有重要的意义。
[1]萧刚柔. 中国森林昆虫[M]. 2版.北京:中国林业出版社,1992:483-485.
[2]Yoh A, Katsumi T. Newly discovered transmission pathway ofBursaphelenchus xylophilusfrom males of the beetleMonochamus alternatustoPinus densifloratrees via oviposition wounds[J]. Journal of Nematology, 2002,34(4): 396-404.
[3]张建军, 张润志, 陈京元. 松材线虫媒介昆虫种类及其扩散能力[J]. 浙江林学院学报,2007,24(3): 350-356.
[4]王柏泉, 徐明飞. 鄂西南地区松褐天牛生物学特性及发生规律研究[J]. 湖北民族学院学报(自然科学版), 2002, 20(2):21-24.
[5]杨忠武, 黄吉平, 杨春生, 等. 桂林市松褐天牛生物学特性研究[J]. 广西林业科学,2010,39(1):5-7.
[6]杨洪, 王进军, 赵志模, 等. 重庆松墨天牛发生规律研究[J]. 中国森林病虫,2010,29(2):15-17.
[7]王陈. 松墨天牛的生物学特性及综合防治技术[J]. 安徽农学通报,2015,21(9):101-102.
[8]温小遂, 廖三腊, 孙计拓, 等. 林间释放花绒寄早对松褐天牛防效的研究[J]. 江西农业大学学报, 2016,38(3): 505-510.
[9]喻爱林, 涂业苟, 金明霞, 等. 甲维盐混配剂诱木注药防治松褐天牛研究[J]. 南方林业科学, 2016,44(4):34-36.
[10]韩水兴. 4种药剂林间注干防治松墨天牛幼虫试验[J]. 生物灾害科学,2017,40(2):93-96.
[11]余建英, 何旭宏. 数据统计分析与SPSS应用[M]. 北京: 人民邮电出版社,2003.
[12]李建庆, 杨忠岐, 张雅林, 等. 杨树上云斑天牛种群的空间格局及抽样技术[J]. 昆虫学报,2009,52(8):860-866.
[13]李广伟, 陈秀琳, 尚天翠, 等. 黄斑星天牛产卵刻槽的空间分布格局及抽样技术[J]. 河南师范大学学报, 2012, 40(6):130-133.
[14]孟俊国, 刘博, 徐华潮. 松墨天牛幼虫在马尾松上的分布规律研究[J]. 浙江林业科技,2012,32(2):41-44.
[15]柴希民, 何志华, 李春才, 等. 松墨天牛成虫产卵特性研究[J]. 北京林业大学学报,1997,19(2):69-72.
猜你喜欢
南方林业科学(2021年3期)2021-07-28
绿色科技(2019年5期)2019-11-29
吉林农业(2019年7期)2019-04-01
中国森林病虫(2017年2期)2017-12-18
湖北植保(2017年4期)2017-08-31
小溪流(画刊)(2017年6期)2017-07-10
广西林业科学(2016年1期)2016-03-20
广西林业科学(2016年1期)2016-03-20
广西林业科学(2016年1期)2016-03-20
广西蚕业(2015年3期)2015-11-30
