
江西官山云锦杜鹃生长规律及群落特征
2018-07-07 01:20余泽平王国兵方平福姚小华杨清培
生物灾害科学 2018年2期
余泽平 ,王国兵 ,方平福 ,姚小华 ,杨清培
(1. 江西官山国家级自然保护区管理局,江西 宜丰 336300;2. 江西农业大学 林学院,江西 南昌 330045)
云锦杜鹃(Rhododendron fortuneiLindl.)为杜鹃花科的常绿灌木或小乔木,观赏价值极高[1-2],江西省 III级重点保护植物。主产于长江流域湖南、江西、安徽、浙江等省区,在海拔 1 000 m以上中山地带组成混交林或纯矮林,是当地植被重要组成成分和生态服务功能的供给者。但同其他山顶群落一样,对气候的变化特别敏感[3], 全球气侯变化可能对这些群落的动态平衡产生不利影响[4], 因此加强对山顶矮林的研究和保护利用就显得尤为重要[5]。
另外,随着自然保护区基础建设的相对成熟。从国家和群众的需求看,探索生态资源维持及其动态监测预报机制成为必须。保护区工作不仅是要保护好各种生境和生物资源,还要进一步提升生态保护与建设的科技含量:让生态资源与服务功能监测动态化、及时化和预测化。
江西官山自然保护区的石花尖、麻菇尖等局部有成片分布的云锦杜鹃矮林,它是官山森林群落中的重要成分,它与周边其他森林或草甸生态系统一起,对维持官山的生态平衡有着十分重要的作用。2014年在“全国生物多样性野外监测示范基地修缮项目”、“ 中央财政林业国家级自然保护区补助资金项目”的支持下,官山保护区积极开展生物资源与生态环境监测与研究[6]。 因此,云锦杜鹃林成为山顶生态环境监测的重要对象。
监测预报需要各项数据的积累。群落结构与物种组成是功能发挥的保证,是监测研究的重要内容,它也决定群落中树木的生长状况,从而影响到种群与群落变化。本文对官山云锦杜鹃群落、生长规律和种群动态开展监测与研究,以期为山顶这一特殊的自然资源、景观资源的保护与利用及可持续发展提供基础数据和理论参考。
1 材料与方法
1.1 研究区概况
江西官山自然保护区位于江西省西北部九岭山脉西段(114°29′~114°45′E,28°30′~28°40′N),海拔 200~1 480 m。气候属中亚热带温暖湿润区,年均气温 16.2 ℃,年降雨量 1 950~2 100 mm(东河站)。常绿阔叶林是这里的典型地带性植被,同时还有大量常绿落叶阔叶混交林、落叶阔叶林、针叶林、针阔混交林、灌丛等植被类型,大量古老、珍稀的植物也呈群落分布[7]。云锦杜鹃灌丛群落位于东河保护站石花尖,地理位置28°34′33.63″N,114°36′5.5″E,海拔 1 200 m,现生长状况良好,保存完整,是保护区面积最好的云锦杜鹃保护地和监测研究基地。
1.2 样地设置与解析木
2016 年,在全面踏查的基础上,选择最具有典型的云锦杜鹃群落,设置长期监测固定样地(GD-33),样地大小 200 m2。 采用相邻格子法把每块样地分成 2 个10 m×10 m的样方,对样地内所有胸径≥1.0 cm的木本植物进行挂牌,每木检尺,测定其树高、胸径、冠幅生长。同时,记录样地内云锦杜鹃的幼苗。同时对林下灌木、草本进行描述性调查。
在样地附近选择 1 株云锦杜鹃标准木,采 用 monsi 分层切割法,主要以1 m为区段,截取圆盘带回实验室,以5 a生为龄阶对解析木圆盘进行分析(图 1),分别测量并计算每个龄阶的直径和树高,建立树高、胸径与年龄的关系[8]。
1.3 数据处理
群落组成群落中各物种重要值(IV)、物种多样性 Simpson指数、Shannon-Wiener 指数、Pielou均匀度指数、生态优势度指数等指标参考文献[9-10],种群静态生命表绘制及各项指标意义与计算参考文献[11]。
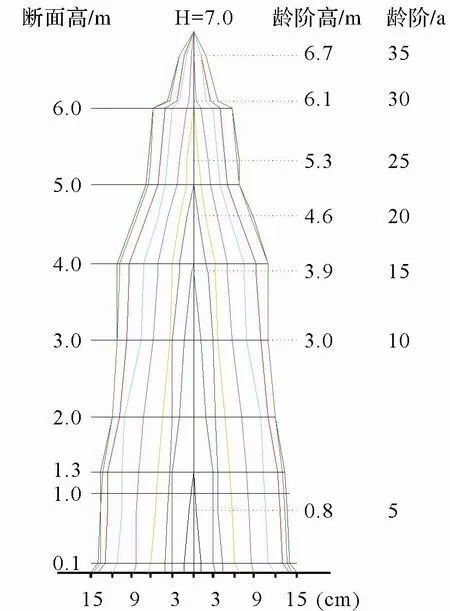
图1 云锦杜鹃树干纵剖面
2 结果与分析
2.1 群落特征
2.1.1 群落组成 官山保护区云锦杜鹃群落是一种典型的山顶矮林。 群落结构与物种组成相对简单 (表 1)。云锦杜鹃高7.0 m左右,为群落优势种,重要值为59.72。伴生种主要有杜鹃(Rhododendron simsii)、圆锥绣球(Hydrangea paniculata)、山樱花(Cerasus serrulata)等,群落密度3 950株/hm2, 主林层郁闭度约0.7。

表1 官山云锦杜鹃监测样地群落物种组成及重要值
2.1.2 物种多样性 官山云锦杜鹃群落物种组成简单, 物种多样性指数较低 (表 2)。 样地中胸径大于 1.0 cm的物种(DBH>1.0 cm)仅 9 种。云锦杜鹃个体数量在样地中占绝对优势,所以群落 Simpson 指数、Shannon-Wiener 指数、Pielou均匀度指数都较低,而生态优势度较高。

表2 官山云锦杜鹃监测样地群落物种多样性
2.2 生长特征
2.2.1 树高、胸径和材积总生长 云锦杜鹃树高、胸径和材积累积生长都符合舒马切尔(Schumacher)模型D=a×e-b/t(图 2),树高长至 6.0~7.0 m以后,高生长基本停滞,接着主要是径向生长和冠幅生长。
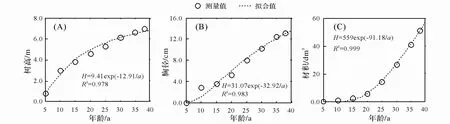
图2 官山云锦杜鹃树高、胸径和材积总生长
2.2.2 树高、胸径与材积生长速率 根据总生长模型,分别求得树高、胸径与材积生长速率(图3)。由图3 可知,云锦杜鹃前期高生长较快,到 6~7 a 生时达到最大(0.4 m/a),然后逐渐减缓,到 40 a 左右每年仅生长 0.05~0.06 m。胸径快速生长期比树高要迟,16~17 a 生才达到最大生长速率(0.5 cm/a),然而材积生长一直在增加,40 a 时还未出现减速。
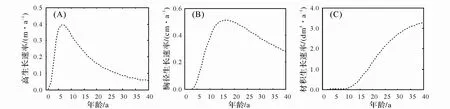
图3 云锦杜鹃树高、胸径和材积生长速率
2.3 种群特征
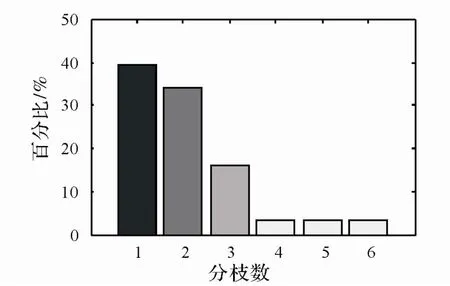
图4 云锦杜鹃不同分枝个体数占种群大小的比例
图5 云锦杜鹃胸径与分枝数的关系
2.3.1 形态特征 群落中云锦杜鹃大多主干明显,在 1.3 m 以下分枝个体中,没有分枝的个体占 39.3%,有 4~6个分枝的个体分别只占 3.6% (图 4)。 另外, 低龄个体分枝数量相对较少, 大龄个体分枝较多 (图 5),可能由于山顶风大,云锦杜鹃前期干性较明显,壮年之后中心干优势减弱或失去优势,促进树木侧枝生长。
2.3.2 静态生命表 采用平均滑技术处理后[12], 编制出云锦杜鹃静态生命表(表 3)。静态生命表中死亡率(qx)、致死力(Kx)反映了种群存活死亡的动态变化。由表 3 可知,20~30 a 时样地中云锦杜鹃的死亡率为25.0%,致死力为 0.29。说明此时种内和种间竞争激烈,无法满足个体对生存空间的强烈需求,导致大量死亡。平均生命期望值(ex)反映的是x龄期个体的平均期望生存能力。云锦杜鹃种群在 0~10 a 时ex值最高(2.83),表明云锦杜鹃幼年的生存质量最高,是生理活动旺盛期。20~30 a生后云锦杜鹃生命力开始减弱,平均期望生存能力较低。

表3 官山云锦杜鹃监测样地种群静态生命表
3 讨论与结论
云锦杜鹃作为园林观赏植物,它是重要的生物资源,作为山顶群落重要的组成成分或自成单优群落,它是生态功能的维持者。因此开展云锦杜鹃长期定位监测研究,对山顶自然资源、景观资源的保护与利用及可持续发展提供基础数据和理论参考,具有重要意义。
群落组成与物种多样性的测定可反映群落及其环境的保护状况,这对指示物种尤其是珍稀濒危物种的丧失是很有意义[10]。 本文研究发现,官山云锦杜鹃监测样地群落结构简单,只有一层 7.0 m左右的冠层,但林分密度大(3 950 株/hm2) , 郁闭度较大(>0.7),但物种组成单一,生物多样性低,远不及广东天井山和湖南幕阜山[13], 而且林下灌木、草木极不发达。样地边缘正在受箬竹(Indocalamussp.)扩张干扰,这可能会导致云锦杜鹃的衰退[14], 应引起关注。
群落生境和群落密度、郁 闭度等内外因子都影响到群落中树木的生长。山 顶风大限制整个群落的高度,同时也限制了群落内树木的生长高度。云锦杜鹃作为群落优势种,其高度被限制在 7.0 m左右,但前期种内种间竞争其树高生长较快(图 3A),而后期主要是径向生长(图 3B)和侧枝生长(图 5),这可能是这种植物的一种表型可塑性策略[15]。 正因侧枝横向生长,冠幅增大,引起了种间种内竞争的加剧,从而导致此时云锦杜鹃死亡率、致死力升高(表 3)。
年龄判定是植物发育生物学、种群生态学基础。本文通过标准木解析的方法,初步建立了云锦杜鹃树高、胸径、材积与年龄间的相关关系,其胸径与年龄关系为D=31.07e-32.62/t。通过以上模型,可较为准确的根据胸径判定植株年龄。初步判断种群年龄结构(表3),发现样地中云锦杜鹃种群年龄结构呈倒三角形,幼苗幼树和大龄个体都较少,这与广东天井山相似[13]。但样地中云锦杜鹃最大个体不到 50 a,远远低于浙江华顶山云锦杜鹃林的 400~600 a[14], 那么如何与其他群落类型相结合,气候与物候相结合,进行长期监测预报,这些问题还需要进一步研究。
致谢:感谢江西农业大学林学 1401 班文仁权、赖媛、刘家福参与样地调查;感谢天津大学理学院杨晓周同学参与解析木分析。
[1]陈艳华, 彭重华. 湖南省阳明山云锦杜鹃的园林观赏特性[J]. 亚热带农业研究, 2009,5(1): 9-12.
[2]郑万钧. 中国树木志(第三卷)[M].北京:中国林业出版社, 1997.
[3]Walther G R, Post E, Convey P, et al. Ecological responses to recent climate change[J]. Nature, 2002, 416(6879): 389-395.
[4]Foster P. The potential negative impacts of global climate change on tropical montane cloud forests[J]. Earth-Science Reviews,2001, 55(1/2): 73-106.
[5]李晓红, 肖宜安, 龙婉婉, 等. 井冈山自然保护区云锦杜鹃群落物种多样性研究[J]. 浙江林业科技, 2005, 25(1): 17-20.
[6]王国兵, 余泽平, 吴钦树,等. 江西官山大型森林样地中珍稀植物及空间分布格局[J]. 南方林业科学, 2017, 45(4): 4-7.
[7]刘信中. 江西官山自然保护区科学考察与研究[M].北京: 中国林业出版社, 2005.
[8]孟宪宇. 测树学[M].北京:中国林业出版社,2006.
[9]钱迎倩. 生物多样性研究的原理与方法[M].北京:中国科学技术出版社, 1994.
[10]王伯荪, 余世孝, 彭少麟,等. 植物群落学实验手册[M].广州: 广东高等教育出版社, 1996.
[11]封磊, 洪伟, 吴承祯, 等. 珍稀濒危植物南方铁杉种群动态研究[J]. 植物科学学报, 2003, 21(5): 401-405.
[12]程红梅. 大蜀山短毛椴种群生命表与生存分析[J]. 浙江大学学报(农业与生命科学版), 2010, 36(3): 341-347.
[13]黄川腾, 唐光大, 刘乐, 等.广东天井山云锦杜鹃种群及其所处群落特征[J]. 西南林业大学学报, 2010, 30(6): 15-19.
[14]管康林, 吴家森, 范义荣, 等. 华顶山云锦杜鹃林衰退原因及对策[J]. 浙江农林大学学报, 2001, 18(2): 195-197.
[15]吕志伟, 边才苗. 天台山云锦杜鹃幼树的构型分析[J]. 浙江农林大学学报, 2007, 24(6): 696-701.
猜你喜欢
华人时刊(2022年15期)2022-10-27
心声歌刊(2021年3期)2021-08-05
广西植物(2021年3期)2021-06-10
健康之家(2021年19期)2021-05-23
华人时刊(2020年13期)2020-09-25
安徽文学(2020年4期)2020-04-15
小聪仔(幼儿版)(2020年12期)2020-02-01
民族音乐(2018年5期)2018-11-17
乡村地理(2018年3期)2018-11-06
夜郎文学(2018年3期)2018-09-12
