
广西大青山西南桦人工林草本优势种群生态位研究
2018-07-07 03:47:34庞圣江杨保国刘士玲邓硕坤贾宏炎
中南林业科技大学学报 2018年6期
庞圣江 ,张 培 ,杨保国 ,刘士玲,邓硕坤 ,贾宏炎
(1.中国林业科学研究院热带林业实验中心,广西 凭祥 532600;2.广西友谊关国家森林生态系统定位观测研究站,广西 凭祥 532600)
生态位(Niche)是植物群落生物多样性、种间竞争与共存的研究基础,属于种群生态学研究的核心领域[1-2]。究其本质而言,生态位是物种在特定生态环境条件下,物种与环境二者之间相互作用的特征表现[3]。它既能有效地反映物种在生态系统的功能和地位,揭示植物种群的种间关系、群落结构及其与环境之间的耦合关系,亦有助于认识种群利用自然资源的地位和作用,对群落的稳定性和生物多样性恢复都具有重要意义[4]。
目前,国内外学者已从植物种群生态位理论[5]、生态位测定方法[6]和应用[7]等方面做了大量论证,在种间关系、群落结构和生物多样性及其形成机制等方面得到了广泛应用。如,张彦雷等[8]研究认为,太岳山油松人工林的林龄、密度、林木树高以及胸径与林下草本植物种类组成和生物量关联性较为密切;王伟伟等[9]对青海大通地区不同树种配置的人工林草本群落生态位研究发现,针阔叶混交林更有利于草本植物群落的生长和发育;同时,刘帅华等[10]利用Levins与Pianka生态位计算公式分析云杉Picea asperata纯林、白桦Betula platyphylla纯林以及云杉-白桦混交林3种人工林的草本植物生态位,结果表明针阔混交林中,草本优势种生态位所代表的群落结构更为合理,混交林的配置模式更适合在当地推广;此外,潘高等[11]对处于植被演替过程中的针叶林和针阔混交林下草本植物的研究表明,草本生态位重叠程度较高时,对资源的利用存在较大的空间异质性,一些群落的结构仍可能处于不稳定的动态变化;彭姣等[12]研究指出,维护油茶Camellia oleifera人工林中具有较多经济开发价值的草本植物多样性,兼顾农林复合生态经营,可获取最大的经济收益。然而,不同造林模式下,林下植物优势种群间的相互关系怎样,各草本优势种群的地位和功能又是如何,是否会对人工林群落结构产生影响,这些问题是否对近自然化改造过程中人工林生态系统的稳定性及生物多样性产生影响,应引起学者们共同关注。
西南桦Betula alnoides为桦木科 Betulaceae 桦木属 Betula 的落叶乔木;具有速生、干形通直、木材纹理美观、适应性广和耐贫瘠干旱等优良特性,天然分布于我国广西、云南和西藏等省区。西南桦人工林为近年来南亚热带地区发展较快、种植面积广、生态和经济效益较好的人工林生态系统之一[13]。草本植物是西南桦林下植被的重要组成部分,在促进人工林养分循环和维护森林立地生产力方面起着不可代替的作用。因此,本研究运用生态位研究理论,对广西大青山林区西南桦人工林草本植物优势种群的生态位宽度、生态位重叠程度和生态响应进行定量研究,将有助于揭示草本优势种在群落中的生态适应性、功能和地位,为解释西南桦人工林群落结构、物种竞争与共存等问题提供理论依据,亦为人工林近自然化改造过程中林下植被的配置和调节、有利于生物多样性的恢复以及建立稳定的生态系统提供参考。
1 研究方法
1.1 研究地概况
大青山林区位于广西凭祥市中国林科院热带林业实验中心(21°57′~ 22°19′N,106°40′~106°59′E)。该林区属于南亚热带季风气候,全年日照时数1 200~1 600 h。年均气温20.5~21.7 ℃,极端高温40.3 ℃,极端低温-1.5 ℃;年均降水量为1 200~1 500 mm,季节分布差异较大,主要集中在4—9月,相对湿度80%~84%。该林区原生植被有季雨林和常绿阔叶林,西南桦主要种植于海拔190~670 m,土壤类型主要为山地红壤,土层深厚、肥沃,土层厚度均在1 m以上。
1.2 样地设置与数据采集
2015年5月以来,在调阅广西大青山林区营林资料和野外全面勘查的基础上,选择该林区具有代表性的西南桦林地,设置面积为20 m×30 m的标准样地,同时,在每个样地内沿对角线的两个角和中心位置,分别设置3个5 m × 5 m的样方,调查林下的植物群落。其中,西南桦纯林调查9个样地27个草本样方,西南桦×红锥Castanopsis hystrix混交林17个样地51个草本样方,西南桦×灰木莲Manglietia glauca混交林 6个样地18个草本样方,西南桦×杉木Cunninghamia lanceolata 混交林15个样地45个草本样方和西南桦×马尾松Pinus massoniana混交林6个样地18个草本样方,共53块标准样地159个草本样方。调查记录样方的草本植物群落中物种数量、株高和盖度等特征,参考植物种重要值选取每种西南桦人工林内前10个草本优势种进行生态位分析,重要值公式=(相对密度+相对频度+相对盖度)/3。
1.3 数据分析
1.3.1 生态位宽度
生态位宽度采用Shannon-Wiener指数[14]和Levins指数[15]计算公式对数据处理分析:
Shannon-Wiener指数:。
Levins指数:。
式中,B(SW)i为物种 i的 Shannon-Wiener生态位宽度指数,B(L)i为物种i的Levins生态位宽指数;Pij为物种i在第j资源位占它利用全部资源位的百分率比例;r为资源位总数(样方数)。Pij=nij/Ni,而 Ni=Σnij,nij为物种 i在第 j样方的数量特征值 (如密度、盖度和重要值等),本研究为物种i在第j个样方内的密度。
1.3.2 生态位重叠
生态位重叠测度公式为:和,其中Lih为物种i与物种h的生态位重叠指数;Lhi为物种h与物种i的生态位重叠指数;B(L)i为物种i 的Levins生态位宽指数;B(L)h为物种h的Levins生态位宽度指数。
1.3.3 生态位响应
生态位响应的测定公式:R=B(L)/ΔLih(i=h),其中R为生态效应速率,B(L)为Levins生态位宽指数[16-17]。
2 结果与分析
2.1 生态位宽度
林下草本植物优势种Shannon-Wiener和Levins生态位宽度指数计算结果,见表1。从表1中可以看出,不同树种配植模式下,草本优势种的生态位宽度各不相同;在同一种植模式下,草本植物优势种的生态位宽度亦是有差异的。在西南桦纯林中,五节芒、弓果黍和铁线蕨等生态位宽度指数较大,对资源的利用能力较强,为该林分草本优势种;在西南桦×红锥混交林中,铁线蕨生态位宽度值最大,半边旗和乌毛蕨生态位宽度值次之,而大多数草本种群生态位宽度值较小,生态幅度较窄;西南桦×灰木莲混交林中,五节芒和蔓生莠竹的生态位宽度值较大,说明这2个草本植物对资源的利用较为充分,在对资源的竞争中处于优势地位;西南桦×杉木混交林中,生态宽度值较大的草本植物分别为五节芒、铁线蕨和半边旗,表明其在林分内处于草本层优势地位;而在西南桦×马尾松混交林中,五节芒、团叶鳞始蕨、铁线蕨和蔓生莠竹等,在资源的竞争中较为强势。

表1 西南桦人工林草本优势种生态位宽度Table 1 Niche breadth of dominant herbal species under Betula alnoides plantation
由表1可知,相同的草本植物,可出现在不同的种植模式,如弓果黍、五节芒、铁芒萁、半边旗和荩草等5种草本植物在西南桦人工林内均有分布,说明这些草本植物种对当地环境的适应性较强。而在不同林分内,同一植物的生态位宽度指数变化较大,如在西南桦纯林内,弓果黍的生态位宽度值最大,在西南桦×红锥混交林和西南桦×杉木混交林,其生态位宽度则较小;在西南桦×红锥混交林内,铁芒萁的生态位宽度较小,在其他4个林分生态位宽度较为居中;在5种林分内,荩草的生态位宽度均处于较低地位。
综上所述,5种西南桦林分草本优势种Shannon-Wiener和Levins生态位宽度指数有所差异,但两者的大小变化趋势较为一致。此外,五节芒、铁线蕨等的生态位宽度值较大,说明它们在西南桦林分内分布广、资源利用具有较大优势,对环境的适应能力较强。
2.2 生态位重叠
草本优势种生态位重叠值分析结果显示(表2~表6):生态位宽度指数较高的物种,与其他种群存在较高的生态重叠,如五节芒的生态位宽度指数较高,且分布广泛、利用资源的能力较强,与其他草本优势种有着较大生态位重叠。一些种植模式中,生态位宽度较窄的草本优势种亦可能出现较高的重叠度,如在纯林和西南桦×红锥混交林,露籽草的生态位宽度较窄(见表1),却与其他草本优势种有着较大生态重叠值;西南桦纯林内,露籽草-弓果黍的生态位重叠值为0.030 7,露籽草-铁线蕨的生态重叠值为0.043 4;而在后者林分内,露籽草-半边旗的生态重叠度达到0.046 9。

表2 西南桦纯林草本优势种生态位重叠值Table 2 Niche overlaps of dominant herbal species under Betula alnoides forest

表3 西南桦×红锥混交林主要草本优势种生态位重叠值Table 3 Niche overlaps of dominant herbal species under Betula alnoides×Castanopsis hystrix forest
总体上来看,西南桦林分内草本优势种生态位重叠程度尚处于较低水平,纯林Lih重叠值变化范围在0~0.030 7之间,Lhi值变化范围为0~0.043 4;西南桦×红锥混交林Lih值变化范围为0~0.008 4,Lhi值变化范围为0~0.046 9;西南桦×灰木莲混交林Lih值变化范围为0~0.053 8,Lhi值变化范围为0~0.079 2;西南桦×杉木混交林Lih值变化范围为0~0.023 7,Lhi值变化范围为0~0.012 6;西南桦×马尾松混交林Lih值变化范围为0~0.064 5,Lhi值变化范围为0~0.059 4;表明5种西南桦林分的草本优势种生态相似性相对较低,生态位重叠范围高度集中在0~0.050 0,占草本植物总对数98%以上。

表4 西南桦×灰木莲混交林草本优势种生态位重叠值Table 4 Niche overlaps of dominant herbal species under Betula alnoides×Manglietia glauca forest
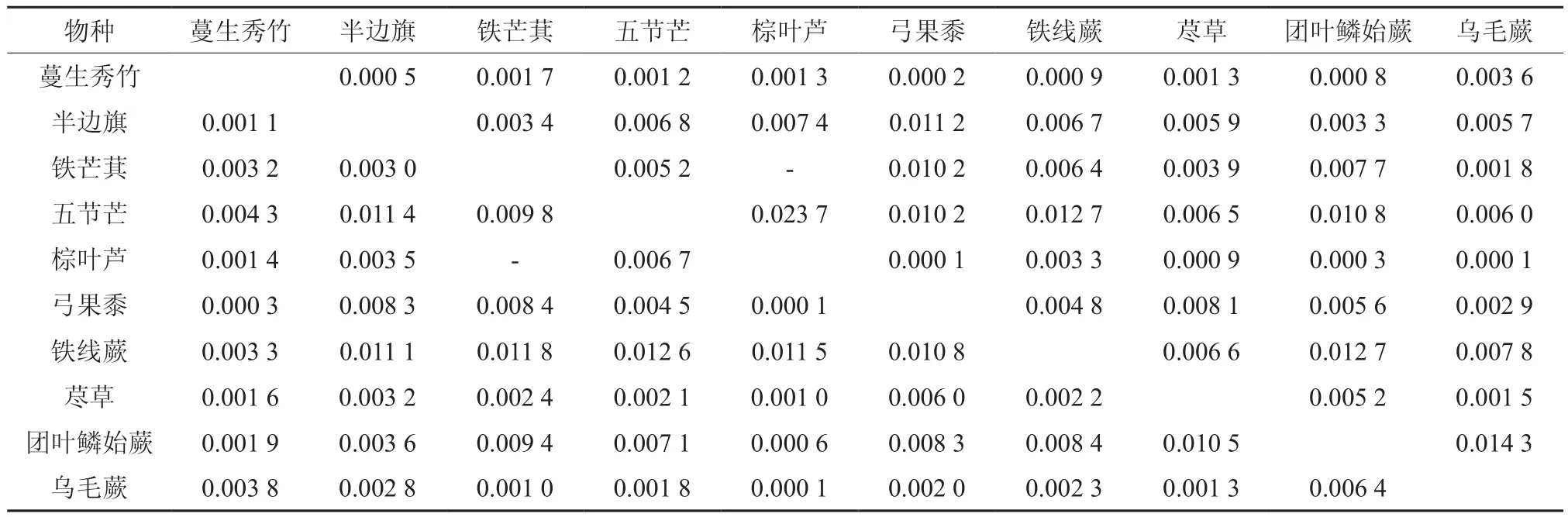
表5 西南桦×杉木混交林草本优势种生态位重叠值Table 5 Niche overlaps of dominant herbal species under Betula alnoides×Cunninghamia lanceolata forest
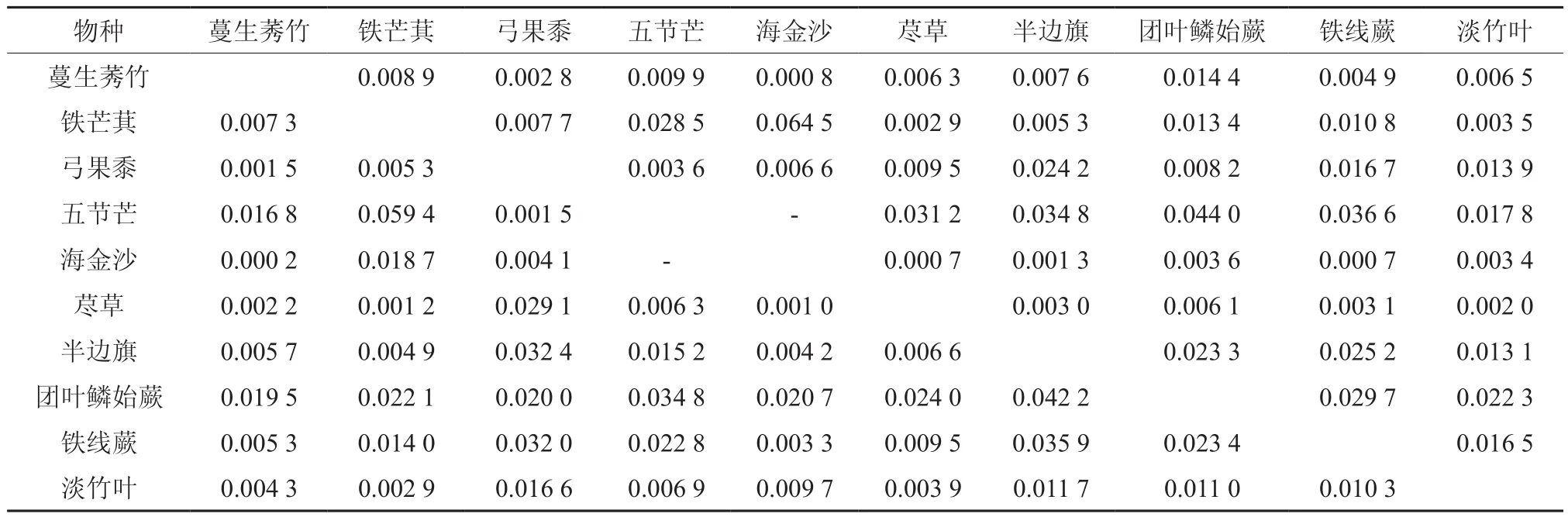
表6 西南桦×马尾松混交林草本优势种生态位重叠值Table 6 Niche overlaps of dominant herbal species under Betula alnoides×Manglietia glauca forest
2.3 生态位响应
从表7可以看出,西南桦纯林中,仅有3个草本植物优势种群为发展型,分别为弓果黍(6.210 3)、五节芒(8.223 7)和铁线蕨(16.895 2);蔓生莠竹、露籽草、铁芒萁等其他7个属于衰退型种群,随着对资源利用竞争加剧,淡竹叶(-117.649 9)和半边旗(-69.149 3)生态空间衰退的趋势更为明显。西南桦×红锥混交林内,发展性最强的是五节芒(20.215 8),其次是铁线蕨(11.385 1)和乌毛蕨(9.870 5),半边旗(3.379 9)的发展性最弱;其他6种属于衰退性种群,生态空间均呈不断缩小的趋势。西南桦×灰木莲混交林中,铁芒萁(22.236 3)的发展性最强,整体上,草本植物群落对资源利用的竞争强度尚处于较低水平。西南桦×杉木混交林中,半边旗(30.376 5) 的发展性最强,而蔓生莠竹、铁芒萁、粽叶芦等6个衰退型种群,在资源的竞争中逐渐处于劣势地位。西南桦×马尾松混交林中,蔓生莠竹(-853.915 7)的衰退最为明显,在资源竞争中处于被动地位,其生存的生态空间有严重退缩的趋势;铁芒萁(53.658 8)的发展性最强,在未来的演替过程中,可能形成优势种群占据草本层的格局。
研究结果表明,草本优势种群表现出的发展型或衰退型,是对该林区西南桦林分内生态环境因子的综合响应。
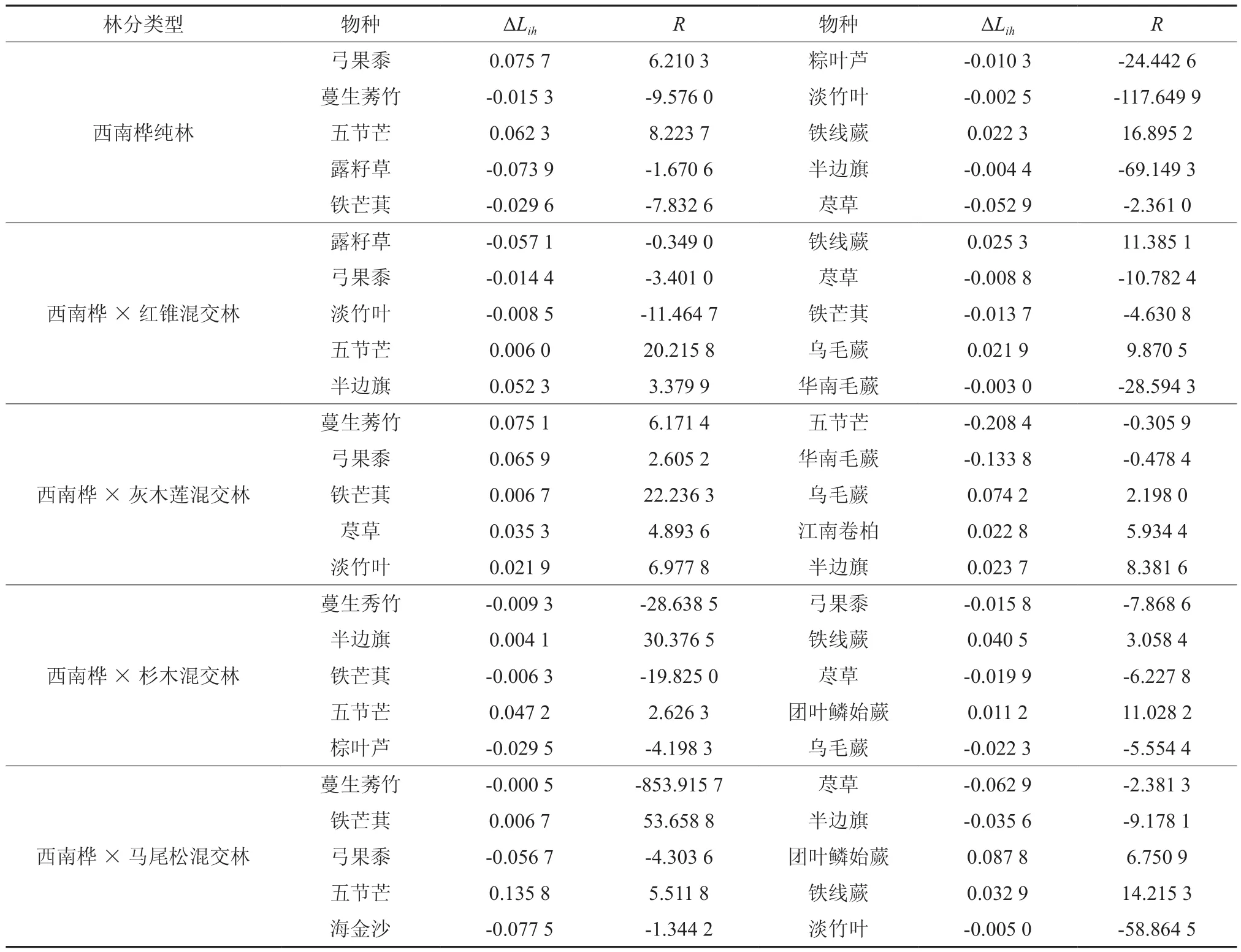
表7 西南桦人工林草本优势种发展或衰退状况Table 7 Aggression and declining situation of dominant herbal species under Betula alnoides plantation
3 结论与讨论
一般情况下,植物种群生态位宽度值越大,资源利用率越高,其对环境适应能力越强[14];同时,说明该物种作为广泛分布种而存在,对资源竞争较为强势;反之,物种则需要在特定环境生存,对资源竞争则处于弱势[18]。本研究发现,5种西南桦林分中五节芒、铁线蕨分布较广,资源利用效率高,适应能力较强;弓果黍、半边旗和铁芒萁在部分林分内数量也比较多。这些生态位宽度值较大的草本植物,生态适应性较强,对生境条件要求较低,故而成为上述研究区域的草本优势种[19-20]。华南毛蕨、江南卷柏等草本植物,仅出现于少数西南桦林分,生态位较窄,适应性较差。虽然露籽草和团叶鳞始蕨2个种群数量较多,但分布较为集中,致使其生态位宽度值也不高。总体而言,5种西南桦林分的草本优势种群生态位宽度值偏低,介于0.019 9~0.748 5;且最大值和最小值两极值间差别大,致使西南桦林分内出现常见种与偶见种共存的现象。
研究发现,5种西南桦林分内草本优势种生态位宽度指数Shannon-Wiener 和 Levins指数亦有所不同,但两者的变化趋势基本一致,在一定程度上能够反映草本植物优势的种生态位现状,这与李菁等[21]对于兴安落叶松人工林林下草本植物种群生态位的研究结论基本一致,他们研究认为这一现象可能与草本优势种对资源的利用效率有关。5种西南桦林分的草本植物群落,在内外因素的综合影响下,草本优势种生态位差异较大。西南桦作为落叶树种,纯林种植或者与马尾松、杉木配植时,林内透光性较好,为五节芒、铁芒萁等喜光植物提供适宜的生长环境;与灰木莲、红锥等阔叶树种混交林中,林内光照相对较弱,但却是蔓生莠竹、弓果黍和铁线蕨等耐阴植物生长的理想环境。因而,在不同西南桦林分,这些草本优势种生态位宽度值均较大,成为草本优势种群。有学者研究指出,植物的生物学及生态特征等内在因素对种群发展变化起着决定性的作用;其次,立地条件等外在因素通过影响光照、土壤养分和水分含量及微生物种类及数量,进而改变植物种群生态位[22]。露籽草、荩草和华南毛蕨等草本优势种群,其生态宽度指数值相对较小(见表1),说明适合其生存的生境较少,仅在少数林地内出现,容易遭受人为因素的干扰,在一定程度上限制了种群的发展。从人工林生物多样性恢复与保护的角度来考虑,生态位宽度值较小的草本植物,则是森林生物多样性保护的重点对象。
研究证实,5种西南桦林下草本植物优势种的生态位宽度虽然存在一定差异,但在不同林分间大多数优势种均有出现,且分布相对均匀,能够较好地反映出草本植物种群的竞争状况。五节芒、铁芒萁以及铁线蕨等优势种群有着较大生态位宽度值,均为广泛分布种群,资源利用效率高,与其他草本优势种群生态位重叠较大(表2和表4),以往研究者对兴安落叶松人工林林下草本植物生态位研究亦得出相类似观点[21];一些诸如淡竹叶、华南毛蕨等生态位宽度值较小的种群,却有着较大的生态位重叠(见表4),这与冶民生等[23]对岷江干旱河谷主要林下植物种群生态位的研究比较相似。究其原因:其一,西南桦人工林频繁的抚育措施,生态系统遭受严重的影响,致使林内资源的空间异质性较大,草本优势种在适宜生境内,呈现出斑块形式的分布;其二,生态位宽度较小的草本优势种群,其生物生态学特性各有差异,对资源获取或者环境适宜能力存在较大的差别。这些因素的综合作用下,导致某些草本优势种群的生态位宽度值较小,但其却可能存在较高的生态位重叠。整体上看来,草本优势种群间的生态重叠程度较低,这与该林区西南桦人工林人为干扰强度大、草本优势种群较少、群落发展受到一定限制有关。
从草本优势种群△Lih来看:5种西南桦林分内,较大生态位宽度的草本优势种群,往往具有较广的资源利用谱,且大多数属于发展型种群,如五节芒、铁线蕨和铁芒萁等;而具有较小生态位宽度的草本优势种群,对林分环境的适应性较差,大多数属于衰退型种群,如淡竹叶、蔓生莠竹等。从草本优势种的生态响应速率来看:铁线蕨(纯林)、五节芒(西南桦×红锥林)、铁芒萁(西南桦×灰木莲、西南桦×马尾松林)和半边旗(西南桦×杉木林)发展性是最强的,明显超过其他草本植物种群,可以预见,未来演替过程中,这些草本植物种群将会逐渐代替其他衰退型的草本植物,成为相对应的西南桦人工林草本优势种群。而生态位较窄的草本植物,如淡竹叶、蔓生莠竹等在相应林分内将会逐渐消亡。从人工林生物多样性恢复的角度来考虑,如何使生态位宽度窄的草本植物保存下来而不被取代,最有效的办法就是提高其生态位宽度:首先,采取合理的人为干扰措施,释放一些草本优势种群的部分资源空间,增加生态位宽度窄的草本植物可利用资源;其次,减少生态位宽度较大的物种数量,达到降低其对资源的竞争能力。
林下草本优势种群生态位的研究结果,可用于该地区人工新造林植被恢复,亦为针叶人工林的近自然化改造提供了重要的解决途径[19]。植被恢复就是植物群落的建立与演替的进行,核心即是植物种群的更替过程[24]。新造林地进行植被恢复时,首先考虑恢复生态位宽度较大的草本植物种群(如五节芒和铁芒萁),可将这类物种作为先锋草本植物。具有较大生态位重叠值的草本植物,其资源需要和生境条件相似,故在先锋草本植物种群建立,并发展为草本优势种群,发展与上述草本优势种群生态位重叠较大的伴生植物(如铁线蕨、弓果黍和半边旗);之后,进一步引入与草本优势种群生态位相似程度较低的物种,达到充分利用生境资源。同时,采取合理的人为抚育措施,调节种群间相互关系,促使植被进行演替更新,提高新造林地生态系统的生物多样性水平,最终达到人工林下草本植物连续覆盖的目的。人工林近自然改造中,不仅要充分考虑树种的配置,并结合人为抚育措施调整群落结构、改善林内光照条件和可供林下植物生长的环境[25-26];同时,利用生态位理论与生产实践相结合,将林下植物群落的生态位宽度、相似性和重叠程度作为参考指标,种群存在竞争关系时,至少某一些资源需求不重叠[27],确保森林种群关系的平衡,高效利用环境资源,提高人工林群落的生物多样性和生产潜力,最终达到森林健康水平的提升,实现人工林可持续经营目标。
本研究基于生态位理论,对广西大青山5种西南桦人工林的草本植物群落进行研究,为该地区新造林地的人工恢复和人工林近自然改造中植物多样性的保护提供参考依据。对林区在森林经营方面也提出了符合实际的建议,为科学合理地经营西南桦人工林以及通过植被管理提升森林健康水平和生产力具有重要的意义。然而,植物群落的自然分布与气候因子、立地条件和调查时间关系密切,在不同林分的各发育时期,对植物群落生态位的影响也甚大;因此,需进一步深入研究气候、地形和各林分不同发育阶段等生态位的其它影响因子,这也是下一步研究工作的重点。
[1]Whittaker R H, Levin S A, Root R B.Niche, habitat and ecotope[J].American Naturalist, 1973, 107: 321-338.
[2]杨利民,周广胜,王国宏.草地群落物种多样性维持机制的研究Ⅱ物种实现生态位[J].植物生态学报,2001,25(5):634-638.
[3]Leibold M A.The niche concept revisited mechanistic model and community contex[J].Ecology,1995, 76(5):1371-1382.
[4]Mustshinda C M, O’Hara R B.Integrating the niche and neutral perspectives on community structure and dynamics [J].Oecologia, 2011, 166: 241-251.
[5]Westman W E.Measuring realized niche spaces climatic response of chaparral and coastal sage scrub [J].Ecology, 1991,72:1678-1684.
[6]Sylvain D, Daniel C, Clementine G C.Niche separation in community analysis: a new method [J].Ecology, 2000, 81: 2914-2927.
[7]Thuiller W, Gasso’ N, Pina J, et al.Ecological niche and species traits: key drivers of regional plant invader assemblages [J].Biol Invasions, 2012, 14:1963-1980
[8]张彦雷,康峰峰,韩海荣,等.太岳山油松人工林林下植被生物量影响因子分析[J].中南林业科技大学学报, 2015, 35(1):104-108.
[9]王伟伟,杨海龙,贺康宁,等.青海高寒区不同人工林配置下草本群落生态位研究[J].水土保持研究, 2012,19(3):156-165.
[10]刘帅华,贺康宁,董 梅,等.青海大通不同人工林林下草本群落生态位特征[J].四川农业大学学报,2012, 30(2):167-173.
[11]潘 高,张合平,潘 登.南方红壤丘陵区3种森林群落内主要草本植物种群生态位特征[J].草业科学, 2015,32(12):2094-2106.
[12]彭 姣,赵运林,徐正刚,等.常德油茶次生林林下植被物种多样性调查[J].中南林业科技大学学报,2016, 36(12):68-73.
[13]曾 杰,郭文福,赵志刚,等.我国西南桦研究的回顾与展望[J].林业科学研究, 2006, 19(3): 379-384.
[14]张金屯.数量生态学[M].北京:科学出版社, 2004.
[15]魏志琴,李旭光,郝云庆.珍稀濒危植物群落主要种群生态位特征研究[J].西南农业大学学报(自然科学版),2004,26(1):1-4.
[16]王翠红,张金屯.汾河水库及河道中优势硅藻生态位的研究[J].生态学杂志,2004,23(3):58-62.
[17]程占红,牛莉芹.五台山南台山地草甸种群对旅游干扰的生态响应[J].水土保持研究,2008,15(6):222-224.
[18]张德魁,王继和,马全林.古浪县北部荒漠植被主要植物种的生态位特征[J].生态学杂志,2007,26(4):471- 475.
[19]蔡道雄,卢立华,贾宏炎,等.封山育林对杉木人工林林下植被物种多样性恢复的影响[J].林业科学研究,2007,20(3):319-327.
[20]农 友,卢立华,贾宏炎,等.桂西南岩溶区不同恢复模式群落生物量及林下植物多样性[J].林业科学研究,2017, 30(2):200-205.
[21]李 菁,骆有庆,石 娟,等.阿尔山地区兴安落叶松林下植物种群生态位[J].林业科学研究, 2011, 24(5):651-658.
[22]胡相明,程积民,万惠娥.黄土丘陵区不同立地条件下植物种群生态位研究[J].草业学报,2006,15(1):29-35.
[23]冶民生,关文彬,吴 斌,等.岷江干旱河谷主要灌木种群生态位研究[J].北京林业大学学报,2006,28(1):7- 13.
[24]张继义,赵哈林,张铜会,等.科尔沁沙地植物群落恢复演替系列种群生态位动态特征[J].生态学报, 2003, 23(12): 2741-2746.
[25]Hautier Y, Niklaus P A, Hector A.Competition for light causes plant biodiversity loss after eutrophication[J].Science, 2009,324(5927):636-638.
[26]刘延滨,王庆成,王承义,等.退化落叶松人工林近自然化改造对土壤微生物及养分的影响[J].生态学杂志,2012,31(11):2716-2722.
[27]丛沛桐,颜延芬,周福军,等.东北羊草群落种群生态位重叠关系研究[J].植物研究,1999,19(2):212-219.
猜你喜欢
中华建设(2024年3期)2024-03-16 08:04:52
中华建设(2023年10期)2023-10-07 05:23:04
考试与评价·高二版(2021年2期)2021-09-10 07:22:44
今日中国·西班牙文版(2021年4期)2021-04-09 11:22:46
少儿美术(2020年8期)2020-11-06 05:21:38
汉语世界(The World of Chinese)(2019年2期)2019-04-19 01:38:10
现代园艺(2018年2期)2018-03-15 08:01:03
启蒙(3-7岁)(2017年4期)2017-06-15 20:28:55
广西林业科学(2016年1期)2016-03-20 05:33:02
广西林业科学(2016年4期)2016-03-16 05:44:58
