
花后人为模拟倒伏对超级稻生长、产量和品质的影响*
2018-07-05 02:00赵新勇邵在胜吴艳珍赵轶鹏王余龙王云霞杨连新
中国生态农业学报(中英文) 2018年7期
赵新勇, 邵在胜, 吴艳珍, 赵轶鹏, 王余龙, 王云霞, 杨连新**
花后人为模拟倒伏对超级稻生长、产量和品质的影响*
赵新勇1,2, 邵在胜1, 吴艳珍1, 赵轶鹏2, 王余龙1, 王云霞3, 杨连新1**
(1. 扬州大学江苏省作物遗传生理国家重点实验室培育点/粮食作物现代产业技术协同创新中心 扬州 225009; 2. 江苏徐淮地区徐州农业科学研究所 徐州 221121; 3. 扬州大学环境科学与工程学院 扬州 225009)
倒伏是水稻实现高产优质的主要限制因子之一。以‘南粳9108’为材料, 研究人为模拟倒伏对超级稻物质生产和分配、产量和产量构成因素以及稻米品质的影响及其与倒伏发生时期的关系,探讨倒伏对水稻生长、产量和品质的影响, 为超级稻抗倒栽培提供依据。试验设3个处理, 分别为正常植株(CK)、抽穗后15 d倒伏(AL15)和抽穗后30 d倒伏(AL30), 成熟期测定水稻物质积累与分配、产量和产量构成以及稻米主要品质性状。结果表明, 与正常生长水稻相比, AL30和AL15处理使水稻地上部各器官生物量下降, 总生物量分别显著降低7.3%和24.3%。从物质分配看, AL15处理使水稻生物量在生殖器官中的分配比例显著下降, 而AL30处理植株响应不显著。AL30和AL15处理使水稻产量分别显著下降8.3%和36.4%。抽穗后倒伏造成的产量损失主要与饱粒率和饱粒千粒重下降有关。AL30处理使饱粒率和饱粒千粒重分别显著下降5.6%和3.3%, AL15处理分别极显著下降30.8%和6.7%。水稻结实期倒伏导致饱粒重下降主要与糙米变小, 稻谷充实程度明显降低有关。从稻米品质看, AL30和AL15处理使水稻整精米率分别下降7.5%和14.7%, 达显著和极显著水平; AL30处理对稻米蛋白质和直链淀粉含量均无显著影响, 但AL15处理使稻米蛋白质含量极显著增加20.2%; RVA谱数据表明, 结实期倒伏水稻崩解值减少而消减值增加, 但均未达显著水平。以上结果表明, 结实中后期发生倒伏对超级稻‘南粳9108’籽粒产量和稻米品质影响较小, 但结实早期倒伏将使物质生产和籽粒灌浆均受到抑制, 最终导致产量大幅下降, 同时稻米加工和食味品质亦呈变劣趋势。
水稻; 倒伏; 物质生产和分配; 产量; 产量构成; 品质
水稻()是我国约65%人群的主食[1], 而倒伏问题是目前限制水稻丰产、优质和高效的主要因素之一[2-3]。水稻倒伏之后不仅给机械化生产带来困难[4], 还因叶片和根系等器官生长受抑, 阻碍了光合产物、水分和养分向籽粒的运输, 限制了产量潜力的发挥[2,5-6]。从稻米品质看, 倒伏群体通常湿度较大, 利于霉菌生长和各种病虫害发生, 进而导致品质变劣[4]。目前, 稻作生产上高产重穗型品种的推广面积逐渐增加, 直播、抛秧以及机插秧等种植方式的应用面积明显增多; 同时, 全球变化特别是台风和冰雹等极端天气亦呈增加趋势[7-9], 上述变化均有可能增加水稻的倒伏风险[10-13]。因此, 进一步增强水稻抗倒机理及其调控途径的研究显得越来越重要。
尽管外界条件对水稻倒伏抗性的影响、机理及调控方面已有很多报道[14-18], 但水稻因灾(如台风和暴雨等)发生倒伏后对水稻最终产量的影响及其可能原因研究较少[4,18]。这种影响是否因倒伏发生时期而异, 以及稻米品质是否变劣则更不清楚。陈书强等[18]对两水稻品种的大田观察发现, 灌浆期台风引起的倒伏发生越早, 千粒重降幅越大; 刘利华等[19]报道灾后自然倒伏导致水稻大幅减产, 这主要与结实率明显下降有关, 其次亦与粒重变轻有关。但这两例研究均未涉及稻米品质。与因灾导致的自然倒伏水稻相比, 人工模拟倒伏可人为选用代表植株, 控制倒伏的时间和强度, 因此处理精度较高。目前, 利用人工模拟倒伏开展水稻产量和品质响应的研究较少。郎有忠等[20]报道结实期人为倒伏使水稻籽粒产量和稻米品质均呈下降趋势, 降幅与倒伏时期密切相关。上述几例报道均没有分析水稻物质生产与分配的变化及其与产量损失的关系。
超级稻作为中国自主创新的重大技术, 对实现我国水稻单产第三次飞跃具有重要意义[21-22]。超级稻‘南粳9108’是江苏省农业科学院近年来育成的迟熟中粳新品系, 已在生产上大面积推广。其稻米直链淀粉含量低(<10%), 米质介于一般糯米与黏米之间, 被称为软米稻。软米具有滋润爽口、冷却后不变硬、不回生、富有光泽、食用时冷热皆宜等特点[23]。这类水稻倒伏发生后其产量形成和品质性状发生什么变化?这些变化是否因倒伏发生时期而异等问题均不清楚。本研究以‘南粳9108’为供试材料[24], 在抽穗后不同时期对水稻进行人工茎倒伏处理, 系统观察其对水稻生长、产量和品质的影响, 以期为超级稻抗倒栽培提供依据。
1 材料与方法
1.1 材料培育与试验设计
本试验在2015年扬州大学作物遗传生理重点实验室两个土培池中进行。每个土培池大小一致, 均为: 长3.8 m, 宽1.3 m, 深0.45 m。试验池为稻麦轮作, 其土壤基础理化性质为: 有机碳12.7 mg·kg-1, 全氮0.81 mg·kg-1, 全磷1.70 mg·kg-1, 速效磷58.7 mg·kg-1,速效钾63.9 mg·kg-1。本研究以超级软米稻‘南粳9108’为供试品种。育秧方式为旱育秧, 于2015年5月10日进行播种, 6月11日进行移栽, 株距18 cm, 行距20 cm, 每个池子共移栽18行, 每行7穴, 基本苗为每穴两苗, 模拟大田管理, 于10月20日收获。水稻生育期肥料状况: 总氮量15 g·m-2, 其中6月10日施用基肥, 用量是总氮量的60%, 7月20日施用穗肥, 用量占总氮量的40%; 磷和钾肥均为7 g·m-2, 作基肥一次施用。水分管理为: 6月11日—7月22日保持浅水层(约3 cm), 7月23日后干湿交替, 控水搁田。及时防治病虫害, 保证水稻正常发育。
试验将池中正常生长的水稻作为对照, 设置抽穗后30 d、抽穗后15 d人为模拟倒伏处理。人为模拟倒伏的方法如下: 在抽穗后30 d、15 d, 每个土培池选取4行水稻, 间隔1行水稻进行人工模拟倒伏处理, 即每处理两行, 每行作为1个重复。处理时用一只手将距离水稻基部10 cm处握住, 另一只手将水稻顺势折倒, 使水稻折倒后与地面呈平行状态, 且所有倒伏处理水稻植株的方向一致。2015年供试水稻在灌浆期阴雨天为25 d。
1.2 测定内容与方法
成熟期取样前对‘南粳9108’进行茎蘖数调查, 调查穴数为20穴, 计算平均茎蘖数。根据平均茎蘖数, 每个土培池各处理均在两个位置分别取代表性植株5穴。取样方法: 用铲子从水稻基部将水稻挖出, 用清水洗净, 将所有水稻植株分为叶片、茎鞘、稻穗3个部分, 其中叶片和茎鞘洗净后用烘箱在105 ℃下杀青0.5 h, 之后将温度调至80 ℃后烘干, 稻穗则在常温条件下风干。取上述风干后稻穗进行手工脱粒, 用FX-Ⅱ型风选仪区分饱粒、秕粒和空粒。饱粒数用数粒板进行测定, 空秕粒数用手工测定, 测定完粒数后分别称重。用游标卡尺测定稻谷及糙米的长度、宽度和厚度。
将收获的稻谷在室温下放置3个月后测定稻米米质。加工品质按照中华人民共和国农业部部颁标准《NY/T83—1988米质测定方法》, 取每个样品100 g, 对其进行出糙米、出精、挑选整精米, 并且每个步骤之后均称重, 依此数据算得出糙率、出精率和整精米率; 取一定量糙米样品, 用Foss Tecator近红外谷物快速品质分析仪(瑞典)按孙成效等[25]的方法测定直链淀粉及粗蛋白含量; 将精米磨粉后取3 g, 用澳大利亚Newport Scientific仪器公司的Super 3型RVA (Rapid Viscosity Analyzer)测定淀粉的黏滞特性。
根据以上原始测定数据计算以下2级参数:
地上部干重(g·m-2)=叶片干重+茎鞘干重+稻穗干重(1)
叶片占地上部干重比例(%)=叶片干重/地上部干重 ×100 (2)
茎鞘占地上部干重比例(%)=茎鞘干重/地上部干重×100 (3)
稻穗占地上部干重比例(%)=稻穗干重/地上比干重×100 (4)
籽粒产量(g·m-2)=单位面积穗数×每穗颖花数×饱粒千粒重×饱粒率 (5)
饱粒率(%)=样本饱粒数/总粒数×100 (6)
秕粒率(%)=样本秕粒数/总粒数×100 (7)
空粒率(%)=样本空粒数/总粒数×100 (8)
饱粒千粒重(mg)=样本总粒重(g)/总粒数×1 000(9)
所有籽粒平均粒重(mg)=(空粒重+秕粒重+饱粒重)/(空粒重+秕粒数+饱粒数)×1 000 (10)
稻谷(糙米)体积(mm3)=3π×长×宽×厚/4 (11)
籽粒充实度(%)=糙米体积/稻谷体积×100 (12)
出糙率(%)=糙米重/稻谷重×100 (13)
出精率(%)=精米重/稻谷重×100 (14)
整精米率(%)=整精米重/稻谷重×100 (15)
1.3 统计与分析方法
相关数据用Microsoft Excel 2013录入和整理。用SPSS 19.0进行裂区方差分析, 采用一般线性模型, 以倒伏处理为固定因子, 以小区重复为随机因子。各处理采用最小显著差数(LSD)法进行比较。超过LSD0.05或LSD0.01水平的视为显著或极显著。
2 结果与分析
2.1 结实期人为倒伏对‘南粳9108’物质生产与分配的影响
结实期人为倒伏处理对‘南粳9108’地上部及各器官最终干重的影响示于图1。结果表明, 结实期人为倒伏处理使‘南粳9108’地上部总干重及各器官干重均下降, 降幅因处理时期和不同器官而异。与自然生长水稻(对照)相比, 抽穗后30 d倒伏(AL30)处理和15 d倒伏(AL15)处理使水稻地上部干重平均分别下降116 g·m-2、385 g·m-2, 降幅分别为7.3%和24.2%, 分别达显著和极显著水平。从不同器官看, AL30处理使水稻叶片、茎秆和稻穗干重均呈下降趋势, 但只有叶片降幅达显著水平; AL15处理使叶片、茎秆和稻穗干重分别下降19.4%、17.5%和28.4%, 均达极显著水平。多重比较结果表明, AL15处理水稻地上部、叶片和稻穗的干重均显著或极显著小于AL30处理, 但茎鞘干重在两处理间无显著差异。
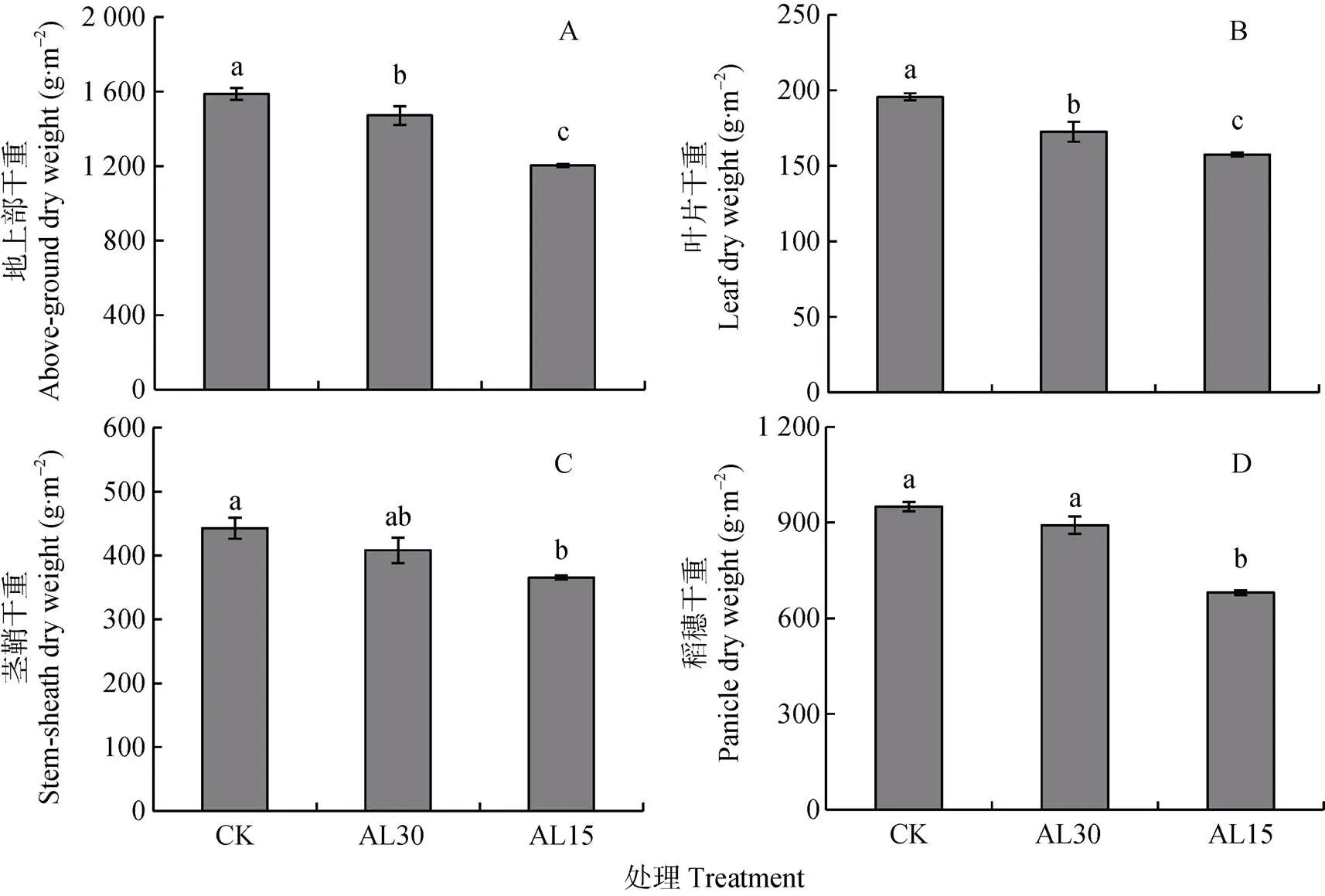
图1 结实期人为倒伏处理对‘南粳9108’成熟期地上部总干重及其各器官干重的影响
CK: 不进行人为倒伏处理; AL30: 抽穗后30 d人为倒伏处理; AL15: 抽穗后15 d人为倒伏处理。不同小写字母表示在0.05水平上差异显著。CK: no lodging; AL30: artificial lodging at 30 days after heading; AL15: artificial lodging at 15 days after heading. Different lowercase letters indicate significant differences at< 0.05.
水稻物质分配对结实期倒伏处理的响应见图2。与对照相比, AL30处理使稻穗占地上部总干重比例平均增加1.3%, 但叶片和茎鞘占地上部干重的比例平均分别下降4.9%和0.5%, 其中叶片比例达显著水平。与AL30处理趋势相反, AL15处理使稻穗比例平均下降5.5%, 叶片和茎鞘比例平均分别增加6.2%和9.1%, 且均达极显著水平。多重比较表明, 结实期两个处理时期之间各器官分配比例均存在极显著差异。
2.2 结实期人为倒伏对‘南粳9108’籽粒产量和产量构成因子的影响
结实期人为倒伏处理对‘南粳9108’籽粒产量的影响示于图3A。人为倒伏处理水稻产量的降幅因处理时期而异。与正常生长水稻相比, AL30和AL15处理使水稻产量平均分别下降74 g·m-2和324 g·m-2, 降幅为8.3%和36.4%, 分别达显著和极显著水平。多重比较表明, AL15处理水稻的产量较AL30处理平均下降30.6%, 差异达极显著水平。可见, 水稻结实期倒伏发生越早, 产量损失越大。
人为倒伏处理对水稻单位面积穗数和每穗颖花数的影响示于图3B-C。由于倒伏处理在抽穗之后实施, 因此对单位面积穗数和每穗颖花数均没有显著影响, 这也说明本试验处理植株具有较好的代表性。单位面积总颖花数为单位面积穗数和每穗颖花数的乘积, 故两个倒伏处理对这一参数也没有显著影响。
水稻饱粒率、秕粒率和空粒率的变化示于图3D-F。与对照相比, AL30、AL15处理使水稻饱粒率平均分别下降5.6%和30.8%, 达显著和极显著水平(图3D)。水稻饱粒率高低与空粒和秕粒的比例有关, 与饱粒率相反, 人为倒伏处理使水稻秕粒率和空粒率均呈增加趋势, 以秕粒率的增幅更为明显(图3E-F)。与对照相比, AL30处理使秕粒数和空粒数分别增加287和32, 其相应秕粒率和空粒率平均分别增加296.0%和9.0%, AL15处理使对应参数分别增加925和852, 其对应比例分别增加941.0%和237.0%, 除AL30处理的空粒率外, 其余均达极显著水平。多重比较表明, AL15理水稻饱粒率较AL30处理下降26.7%, 达显著水平。秕粒率和空粒率则分别增加163.0%和209.0%, 均达极显著水平。

图2 结实期人为倒伏处理对‘南粳9108’叶片、茎鞘和稻穗干重占地上部总重比例的影响
CK: 不进行人为倒伏处理; AL30: 抽穗后30 d人为倒伏处理; AL15: 抽穗后15 d人为倒伏处理。不同小写字母表示在0.05水平上差异显著。CK: no lodging as control; AL30: artificial lodging at 30 days after heading; AL15: artificial lodging at 15 days after heading. Different lowercase letters indicate significant differences at< 0.05.
人为倒伏对水稻饱粒千粒重及所有籽粒平均粒重的影响见图3G-H。AL30处理使饱粒千粒重和所有籽粒平均千粒重分别下降0.9 g和0.6 g, 降幅为3.3%和4.4%, 前者达显著水平; AL15处理使对应指标分别下降1.8 g和5.9 g, 降幅为6.7%和24.4%, 均达极显著水平。多重比较表明, AL15处理水稻饱粒千粒重、所有籽粒平均粒重较AL30处理分别下降3.5%和21.2%, 差异达显著和极显著水平。
2.3 结实期人为倒伏对‘南粳9108’稻谷和糙米形态性状的影响
结实期人为倒伏处理对‘南粳9108’稻谷形态的影响列于表1。与对照相比, AL30处理对稻谷形态性状均没有显著影响, 但AL15处理使稻谷长度、宽度、厚度和体积一致下降, 其中稻谷厚度(-8.3%)和体积(-3.7%)的降幅较大, 前者达显著水平。多重比较表明, AL15处理稻谷的长度、宽度、厚度和体积均小于AL30处理, 其中稻谷厚度差异达显著水平。
从糙米形态看, 结实期倒伏处理使糙米长度均呈增加趋势, 但使宽度、厚度和体积一致下降(表1)。AL30处理使糙米宽度、厚度和体积平均分别下降16.9%、2.1%和15.4%, 其中稻谷宽度和体积达显著和极显著水平。AL15处理使对应指标分别下降17.7%、10.0%和25.4%, 宽度和体积差异达极显著水平。多重比较表明, AL15处理糙米长、宽、厚和体积均小于AL30, 其中长度和体积之间的差异达显著和极显著水平。
水稻籽粒充实度为糙米体积占稻谷体积的百分比。图4表明, AL30和AL15处理使籽粒充实度平均分别下降13.0%和16.0%, 达显著和极显著水平, 但这两个处理之间无显著差异。
2.4 结实期人为倒伏对‘南粳9108’稻米品质性状的影响
稻米加工品质主要指标包括糙米率、精米率和整精米率, 这些指标对倒伏处理的响应示于图5。与对照相比, AL30处理对糙米率、精米率均无显著影响, 使整精米率显著下降7.5%, 达显著水平。AL15处理使糙米率、精米率和整精米率分别下降0.8%、2.8%和14.7%, 其中精米率和整精米率差异分别达显著和极显著水平。多重比较表明, AL15处理水稻的糙米率、精米率和整精米率均低于AL30处理, 其中糙米率(-1.1%)和整精米率(-7.8%)两者间差异达显著水平。
稻米蛋白质和直链淀粉含量对倒伏处理的响应示于图6。AL30处理对稻米蛋白质和直链淀粉含量没有影响, 但AL15处理使稻米蛋白质含量平均增加20.2%, 达显著水平。多重比较表明, AL15处理稻米蛋白质含量较AL30处理高19.2%, 达极显著差异, 但直链淀粉含量在两处理间无显著差异。
稻米淀粉黏滞特性谱的测定结果示于表2。倒伏处理使稻米最高黏度、崩解值、最终黏度和峰值时间均呈下降趋势, 而消减值相反。与对照相比, AL30处理使最高黏度、崩解值、最终黏度和峰值时间平均分别下降10.6%、2.4%、10.1%和0.5%, 其中最终黏度达显著水平。AL15处理使对应参数分别下降14.2%、5.6%、12.9%和1.1%, 其中最高黏度和最终黏度达显著水平。对消减值而言, AL30和AL15处理稻米分别增加1.6%和2.3%, 但未达显著水平。多重比较表明, 结实期两处理间只有最终黏度表现出显著差异。
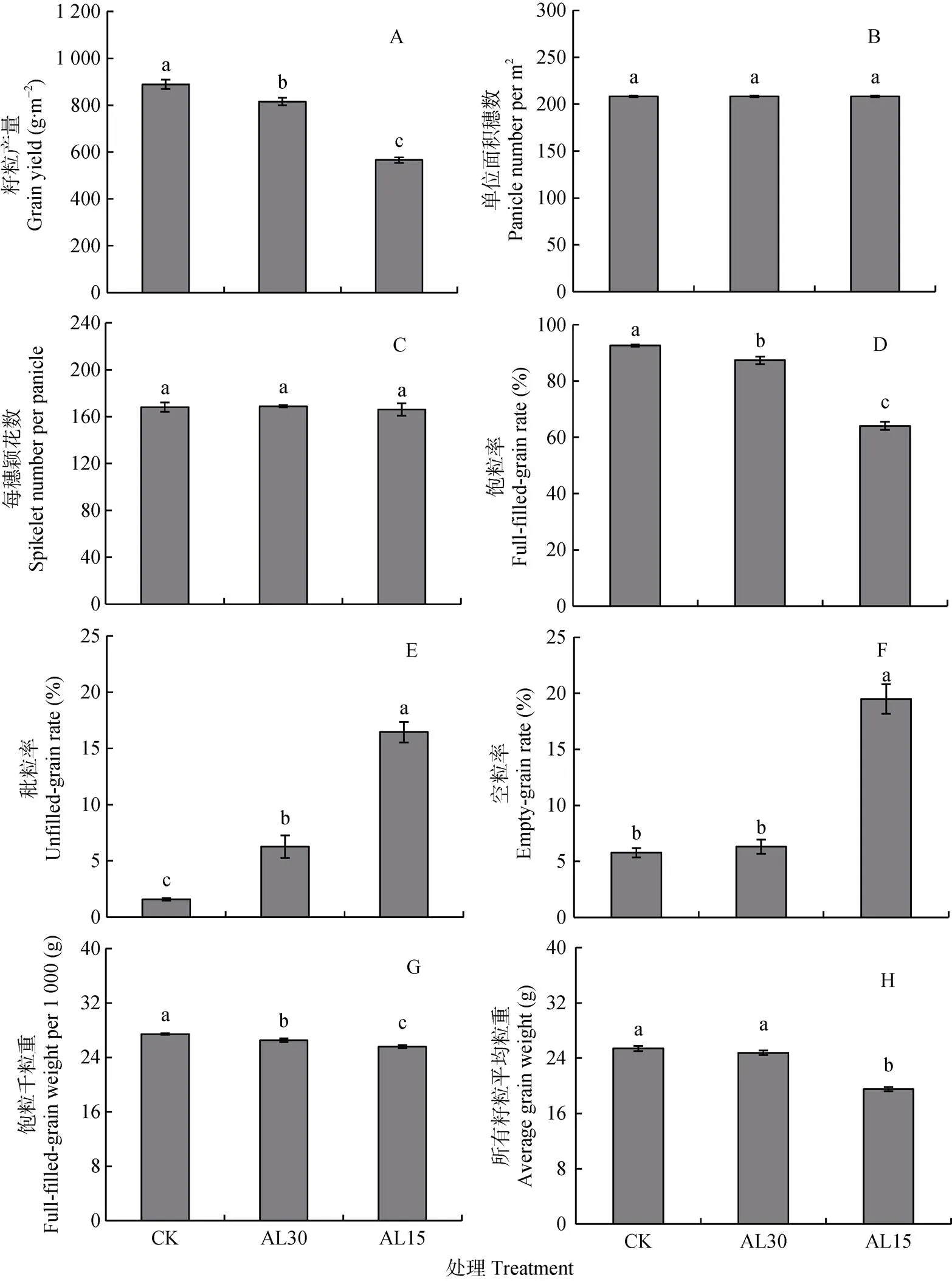
图3 结实期人为倒伏处理对‘南粳9108’籽粒产量(A)、单位面积穗数(B)、每穗颖花数(C)、饱粒率(D)、秕粒率(E)、空粒率(F)、饱粒千粒重(G)及所有籽粒平均粒重(H)的影响
CK: 不进行人为倒伏处理; AL30: 抽穗后30 d人为倒伏处理; AL15: 抽穗后15 d人为倒伏处理。不同小写字母表示在0.05水平上差异显著。CK: no lodging as control; AL30: artificial lodging at 30 days after heading; AL15: artificial lodging at 15 days after heading. Different lowercase letters indicate significant differences at< 0.05.
3 讨论
倒伏水稻叶片呈叠压状态, 光合速度下降, 物质积累明显受抑[2,5-6]。水稻发生倒伏对物质积累与分配的影响报道甚少。本研究表明, AL30处理对水稻地上部各器官干重及总重的影响较少, 但AL15处理下水稻这些参数的降幅均达显著或极显著水平,最大降幅接近30%。受逆境胁迫的作物通常物质分配亦会发生变化[26]。本试验发现结实期人工倒伏对‘南粳9108’物质分配的影响因处理时期而异。AL15处理使生物量在稻穗中的分配比例显著减少, 相应地在茎鞘和叶片中的分配比例明显增加; 而AL30处理的趋势刚好相反。这一现象说明, 水稻结实前期倒伏由于胁迫程度较重会抑制光合物质向稻穗的运转; 与此相反, 灌浆中后期倒伏由于胁迫程度较轻反而有利于物质向稻穗的转移。在干旱研究中亦有类似现象的报道, 即适度干旱会促进茎鞘贮藏性物质向稻穗的转移[27-28], 这种资源分配策略的改变可能是逆境作物的一种自我适应。
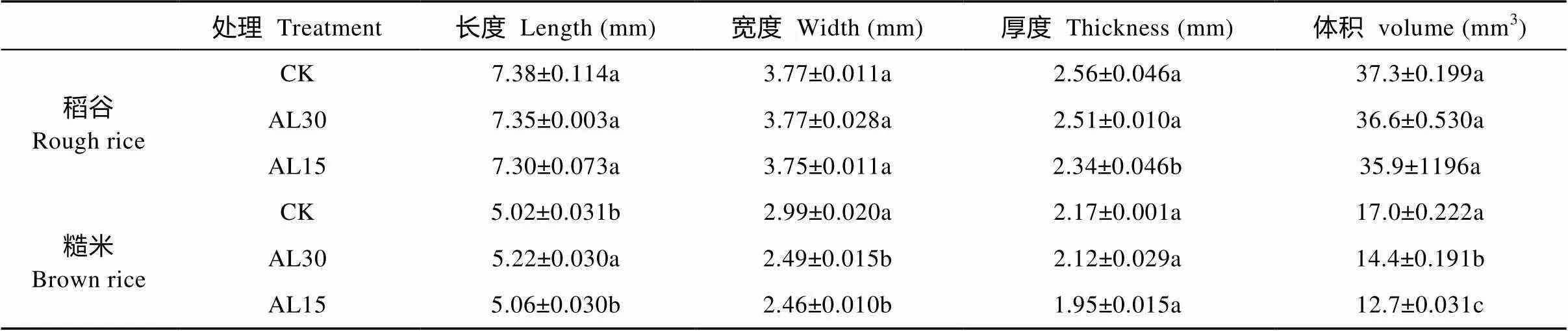
表1 结实期人为倒伏处理对‘南粳9108’稻谷和糙米长度、宽度、厚度和体积的影响
CK: 不进行人为倒伏处理; AL30: 抽穗后30 d人为倒伏处理; AL15: 抽穗后15 d人为倒伏处理。不同小写字母表示在0.05水平上差异显著。CK: no lodging as control; AL30: artificial lodging at 30 days after heading; AL15: artificial lodging at 15 days after heading. Different lowercase letters indicate significant differences at< 0.05.
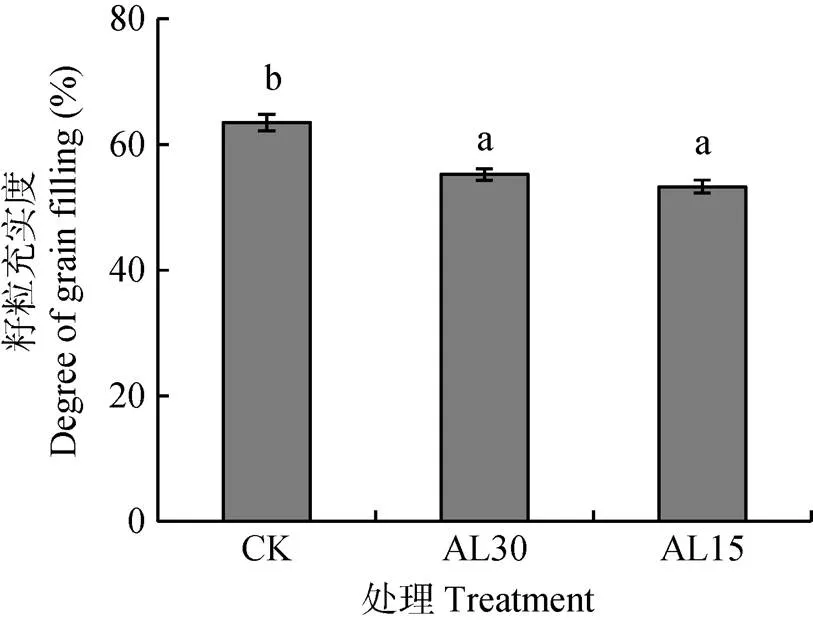
图4 结实期人为倒伏处理对‘南粳9108’籽粒充实度的影响
CK: 不进行人为倒伏处理; AL30: 抽穗后30 d人为倒伏处理; AL15: 抽穗后15 d人为倒伏处理。不同小写字母表示在0.05水平上差异显著。CK: no lodging as control; AL30: artificial lodging at 30 days after heading; AL15: artificial lodging at 15 days after heading. Different lowercase letters indicate significant differences at< 0.05.
水稻结实期自然倒伏或人工模拟倒伏会影响光合作用和生长发育, 进而导致籽粒产量下降[18,20]。郎有忠等[20]报道人工模拟倒伏处理使两水稻品种(‘两优培九’和‘CY-6’)产量平均下降4.9%~83.9%, 倒伏发生时间越早, 减产越多。本研究表明, 与对照水稻相比, AL30处理和AL15处理使产量平均分别下降约8%(>0.1)和36%(<0.01), 后者是前者的4倍多, 说明产量损失程度与倒伏发生时期关系密切。本试验倒伏处理的产量损失明显小于郞有忠等[20]报道的结果, 该研究观察发现AL30处理使两品种产量分别下降41%和64%。笔者认为上述差异的可能原因有以下两个: 首先是两者人工倒伏的方式不同, 本试验是一次性折断稻株, 而郎有忠等[20]试验处理为连续3 d对稻株进行持续压迫, 受胁迫的程度更为强烈, 因此产量降幅更大。其次可能与品种差异有关, 本试验采用的高产优质新品种‘南粳9108’茎秆较粗, 倒伏后恢复能力更强[20]。当然, 上述假设还需选用更多品种进行比较验证。
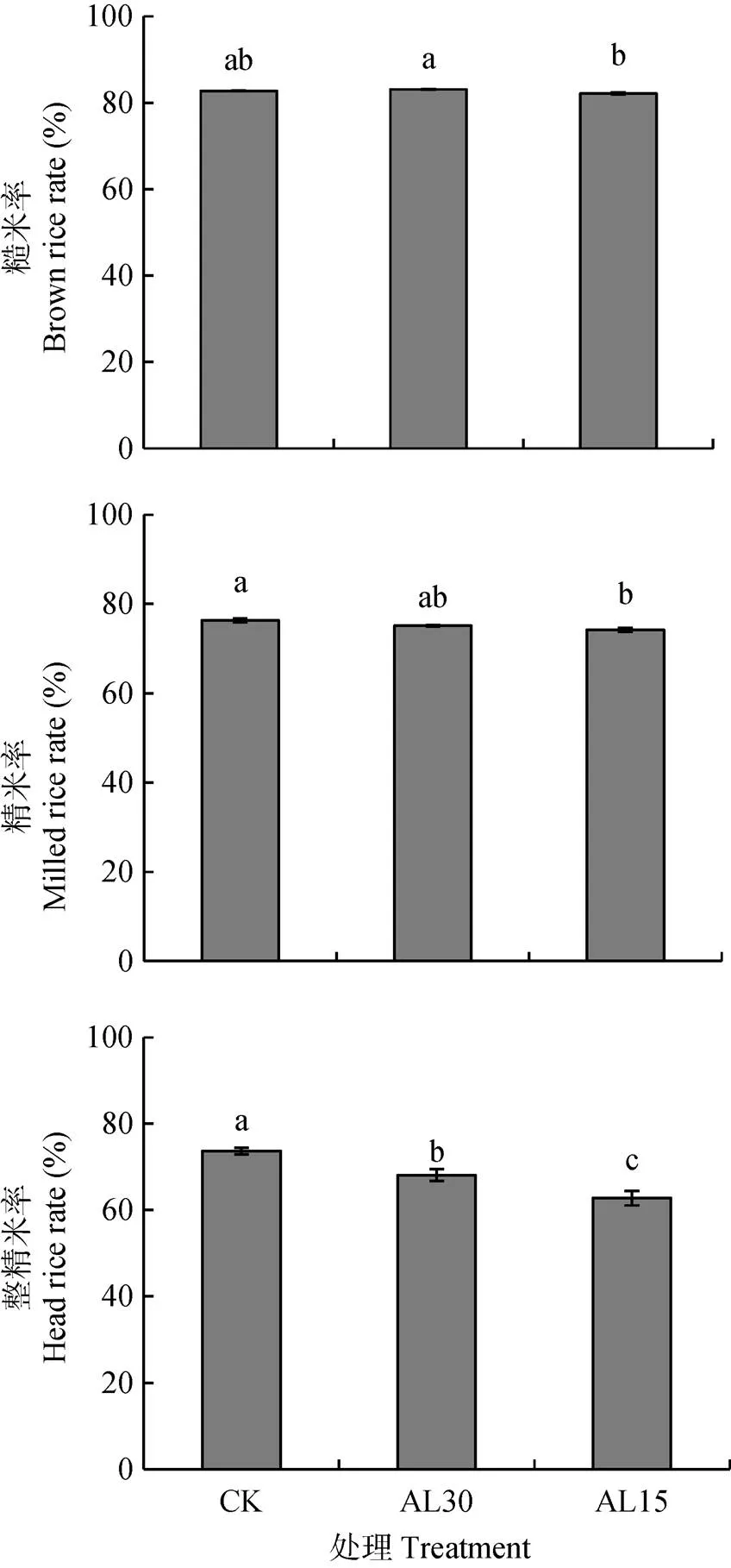
图5 结实期人为倒伏处理对‘南粳9108’稻米糙米率、精米率和整精米率的影响
CK: 不进行人为倒伏处理; AL30: 抽穗后30 d人为倒伏处理; AL15: 抽穗后15 d人为倒伏处理。不同小写字母表示在0.05水平上差异显著。CK: no lodging as control; AL30: artificial lodging at 30 days after heading; AL15: artificial lodging at 15 days after heading. Different lowercase letters indicate significant differences at< 0.05.
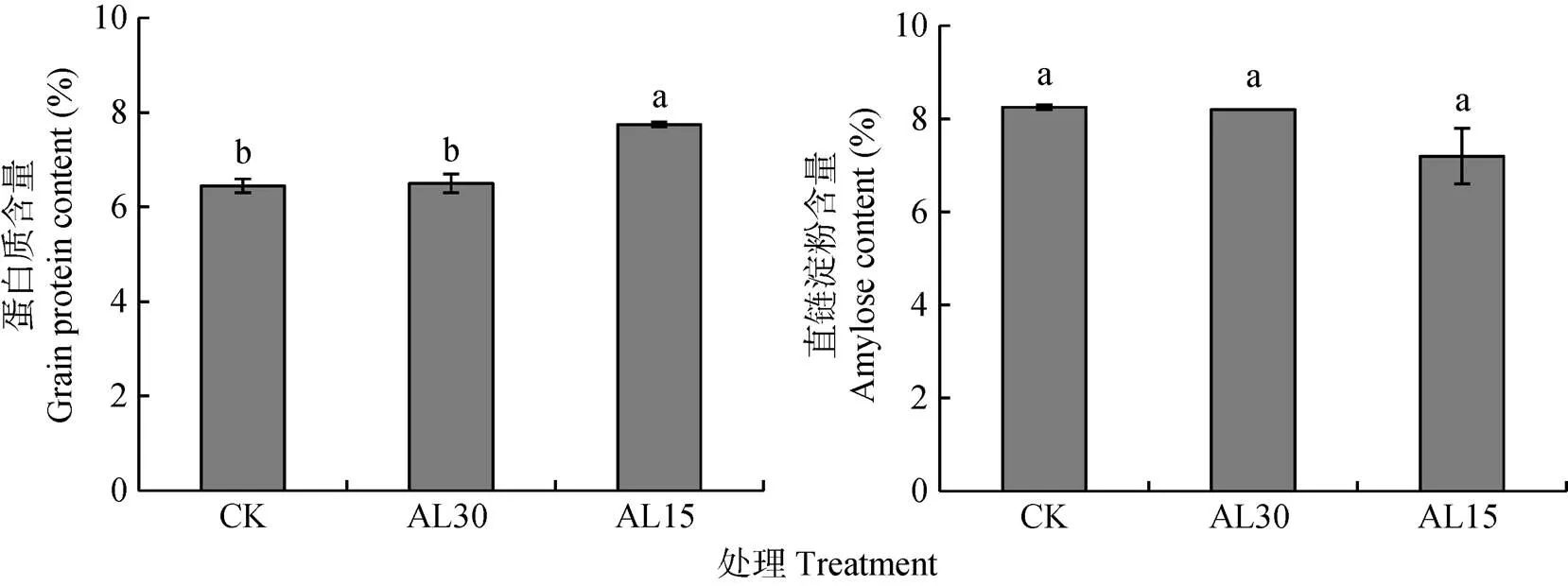
图6 结实期人为倒伏处理对‘南粳9108’稻米蛋白质和直链淀粉含量的影响
CK: 不进行人为倒伏处理; AL30: 抽穗后30 d人为倒伏处理; AL15: 抽穗后15 d人为倒伏处理。不同小写字母表示在0.05水平上差异显著。CK: no lodging as control; AL30: artificial lodging at 30 days after heading; AL15: artificial lodging at 15 days after heading. Different lowercase letters indicate significant differences at< 0.05.
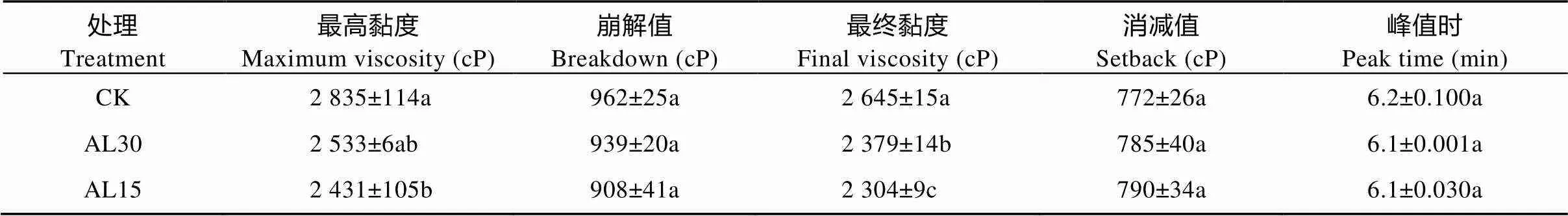
表2 结实期人为倒伏处理对‘南粳9108’淀粉黏滞特性的影响
CK: 不进行人为倒伏处理; AL30: 抽穗后30 d人为倒伏处理; AL15: 抽穗后15 d人为倒伏处理。不同小写字母表示在0.05水平上差异显著。CK: no lodging as control; AL30: artificial lodging at 30 days after heading; AL15: artificial lodging at 15 days after heading. Different lowercase letters indicate significant differences at< 0.05.
水稻穗数和每穗颖花数在抽穗前即已确定, 故结实期倒伏处理对产量构成因素的影响主要表现在饱粒千粒重特别是饱粒率的明显下降, 这与之前的报道一致[18]。本试验相关分析亦表明, 水稻籽粒产量与穗数、每穗颖花数相关不密切, 但与饱粒率(=0.974,=6)和饱粒千粒重(=0.882,=6)均呈极显著正相关, 说明结实期倒伏水稻的产量损失主要与饱粒率大幅下降有关, 其次亦与粒重减少有关。结实期倒伏水稻的饱粒率大幅下降主要与灌浆过程受抑有关, 表现在两期倒伏水稻的秕粒率均大幅增加。对结实早期倒伏的水稻而言, 空粒率也呈增加趋势, 这可能与部分高位分蘖(特别是着生在伸长节间上的小分蘖)迟开小穗的败育有关。水稻饱粒千粒重与籽粒大小和充实度有关[29]。本研究表明, 人为倒伏水稻的粒重减轻主要与稻谷充实度显著下降有关, 稻谷体积降幅相对较小, 后者与谷壳大小在抽穗前就已确定有关[30-31]。与稻谷大小变化较小不同, 结实期人为倒伏处理使糙米宽度、厚度明显减少, 进而糙米体积大幅下降, 最大降幅达25%, 这与稻谷充实度显著下降相吻合。
本研究还表明, AL30处理仅使水稻整精米率显著下降(-7%), 而AL15处理使精米率(-3%)和整精米率(-15%)均显著下降。这与郎有忠等[20]的结果基本一致, 但本试验整精米率的降幅更为明显。结实期倒伏水稻加工品质变劣可能与叶片光合下调导致籽粒充实不良、籽粒紧实度下降有关。
蛋白质是稻米营养品质的重要指标, 对米饭的食味品质也有很大影响[32]。AL30处理对稻米蛋白质含量没有影响, 但AL15处理使之增加20%, 增幅明显大于朗有忠等[20]的结果。与正常成熟水稻相比, 倒伏水稻稻米蛋白质浓度增加可能与“浓缩机制”有关, 即逆境对淀粉积累的影响大于蛋白合成[33]。尽管AL15处理稻米蛋白质浓度增加, 但因产量下降导致单位面积蛋白质产量平均下降15.6%, 达极显著水平(数据未列出)。蛋白质含量的变化会影响蒸煮稻米的淀粉黏滞特性, 通常低的蛋白质含量与高的最高黏度和崩解值相关联[33]。本研究淀粉RVA谱特征值表明, 结实期发生倒伏使稻米崩解值呈减少趋势, 而消减值表现相反, 这种变化与倒伏水稻的稻米蛋白质含量增加相吻合。根据已有的报道[20], 倒伏水稻的食味品质呈下降趋势[34]。综合上述结果, 结实中期倒伏与正常成熟植株的稻米品质差异很小, 但结实早期发生倒伏可能导致稻米总体变劣。
本文研究了结实期人工模拟倒伏对超级软米稻生长、产量和品质的影响及其与倒伏时期的关系。结果发现, 灌浆中期倒伏(灌浆后30 d)对水稻生长、产量和品质的影响较小, 34个测定指标中只有12个指标达显著水平; 而灌浆早期倒伏(灌浆后15 d)使水稻生长明显受抑, 产量大幅下降(-36%), 且上述指标中有21个指标达显著水平。产量下降主要与灌浆结实受阻有关, 表现在饱粒的比例、重量以及充实程度均明显变小。结实早期倒伏还使多数稻米品质性状变劣, 特别是加工和食味品质。需要指出的是, 本试验为单品种两期人工倒伏处理, 未来研究还需设置更多品种以及倒伏时间梯度的试验, 以揭示不同类型水稻对多期模拟倒伏的响应及其生理生态学机制。
[1] 辛良杰, 李秀彬. 近年来我国南方双季稻区复种的变化及其政策启示[J]. 自然资源学报, 2009, 24(1): 58–65 XIN L J, LI X B. Changes of multiple cropping in double cropping rice area of southern China and its policy implications[J]. Journal of Natural Resources, 2009, 24(1): 58–65
[2] SETTER T L, LAURELES E V, MAZAREDO A M. Lodging reduces yield of rice by self-shading and reductions in canopy photosynthesis[J]. Field Crops Research, 1997, 49(2/3): 95–106
[3] ISLAM M S, PENG S B, VISPERAS R M, et al. Lodging-related morphological traits of hybrid rice in a tropical irrigated ecosystem[J]. Field Crops Research, 2007, 101(2): 240–248
[4] 苏仕华, 王珏, 孙成亮, 等. 水稻倒伏对产量影响的调查与分析[J]. 北方水稻, 2008, 38(6): 41–43 SU S H, WANG J, SUN C L, et al. Investigation and analysis of effect of lodging on rice yield[J]. North Rice, 2008, 38(6): 41–43
[5] WU W, HUANG J L, CUI K H, et al. Sheath blight reduces stem breaking resistance and increases lodging susceptibility of rice plants[J]. Field Crops Research, 2012, 128: 101–108
[6] KASHIWAGI T, HIROTSU N, UJIIE K, et al. Lodging resistance locusimproves physical strength of the lower plant part under different conditions of fertilization in rice (L.)[J]. Field Crops Research, 2010, 115(1): 107–115
[7] Solomon S, Qin D, Manning M, et al. Climate Change 2007: The Physical Science Basis: Contribution of Working Group Ⅰ to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change[R]. Cambridge, UK: Cambridge University Press, 2007
[8] KNUTSON T R, MCBRIDE J L, CHAN J, et al. Tropical cyclones and climate change[J]. Nature Geoscience, 2010, 3: 157–163
[9] MASUTOMI Y, IIZUMI T, TAKAHASHI K, et al. Estimation of the damage area due to tropical cyclones using fragility curves for paddy rice in Japan[J]. Environmental Research Letters, 2012, 7(1): 17–35
[10] 艾治勇, 马国辉. 超级杂交稻抗倒高产肥料运筹技术的数学模型研究[J]. 植物营养与肥料学报, 2011, 17(4): 803–808 AI Z Y, MA G H. Study on mathematical models of fertilizer management technique for resisting lodging and high yield of super hybrid rice[J]. Plant Nutrition and Fertilizer Science, 2011, 17(4): 803–808
[11] 李杰, 张洪程, 龚金龙, 等. 不同种植方式对超级稻植株抗倒伏能力的影响[J]. 中国农业科学, 2011, 44(11): 2234–2243 LI J, ZHANG H C, GONG J L, et al. Effects of different planting methods on the culm lodging resistance of super rice[J]. Scientia Agricultura Sinica, 2011, 44(11): 2234–2243
[12] 李国辉, 钟旭华, 田卡, 等. 施氮对水稻茎秆抗倒伏能力的影响及其形态和力学机理[J]. 中国农业科学, 2013, 46(7): 1323–1334 LI G H, ZHONG X H, TIAN K, et al. Effect of nitrogen application on stem lodging resistance of rice and its morphological and mechanical mechanisms[J]. Scientia Agricultura Sinica, 2013, 46(7): 1323–1334
[13] 王云霞, 王晓莹, 杨连新, 等. 臭氧胁迫使两优培九倒伏风险增加——FACE研究[J]. 生态学报, 2011, 31(20): 6098–6107WANG Y X, WANG X Y, YANG L X, et al. Ozone stress increases lodging risk of rice cultivar Liangyoupeijiu: A FACE study[J]. Acta Ecologica Sinica, 2011, 31(20): 6098–6107
[14] 胡雅杰, 曹伟伟, 钱海军, 等. 钵苗机插密度对不同穗型水稻品种产量、株型和抗倒伏能力的影响[J]. 作物学报, 2015, 41(5): 743–757 HU Y J, CAO W W, QIAN H J, et al. Effect of planting density of mechanically transplanted pot seedlings on yield, plant type and lodging resistance in rice with different panicle types[J]. Acta Agronomica Sinica, 2015, 41(5): 743–757
[15] 许俊伟, 孟天瑶, 荆培培, 等. 机插密度对不同类型水稻抗倒伏能力及产量的影响[J]. 作物学报, 2015, 41(11): 1767–1776 XU J W, MENG T Y, JING P P, et al. Effect of mechanical-transplanting density on lodging resistance and yield in different types of rice[J]. Acta Agronomica Sinica, 2015, 41(11): 1767–1776
[16] 刘立军, 袁莉民, 王志琴, 等. 旱种水稻倒伏生理原因分析与对策的初步研究[J]. 中国水稻科学, 2002, 16(3): 225–230 LIU L J, YUAN L M, WANG Z Q, et al. Preliminary studies on physiological reason and countermeasure of lodging in dry-cultivated rice[J]. Chinese Journal of Rice Science, 2002, 16(3): 225–230
[17] 张明聪, 刘元英, 罗盛国, 等. 养分综合管理对寒地水稻抗倒伏性能的影响[J]. 中国农业科学, 2010, 43(21): 4536–4542 ZHANG M C, LIU Y Y, LUO S G, et al. Effects of integrated nutrient management on lodging resistance of rice in cold area[J]. Scientia Agricultura Sinica, 2010, 43(21): 4536–4542
[18] 陈书强, 杜晓东, 杨丽敏, 等. 水稻倒伏粒重损失及不同品种抗倒能力差异比较[J]. 黑龙江农业科学, 2015, (10): 34–37 CHEN S Q, DU X D, YANG L M, et al. Effect of lodging on yield and comparison of different varieties lodging resistance in rice[J]. Heilongjiang Agricultural Sciences, 2015, (10): 34–37
[19] 刘利华, 金再欣, 刘小丽, 等. 水稻倒伏对产量影响的试验结果分析[J]. 中国稻米, 2009, (2): 19–21 LIU L H, JIN Z X, LIU X L, et al. Influence trial of lodging on rice yield[J]. China Rice, 2009, (2): 19–21
[20] 郎有忠, 杨晓东, 王美娥, 等. 结实阶段不同时期倒伏对水稻产量及稻米品质的影响[J]. 中国水稻科学, 2011, 25(4): 407–412 LANG Y Z, YANG X D, WANG M E, et al. Effects of lodging at different filling stages on rice grain yield and quality[J]. Chinese Journal of Rice Science, 2011, 25(4): 407–412
[21] 程式华. 中国超级稻育种研究的创新与发展[J]. 沈阳农业大学学报, 2007, 38(5): 647–651 CHENG S H. Innovation and development of rice breeding for super high yield in China[J]. Journal of Shenyang Agricultural University, 2007, 38(5): 647–651
[22] 陈温福, 徐正进, 唐亮. 中国超级稻育种研究进展与前景[J]. 沈阳农业大学学报, 2012, 43(6): 643–649 CHEN W F, XU Z J, TANG L. Advances and prospects in research on super rice breeding[J]. Journal of Shenyang Agricultural University, 2012, 43(6): 643–649
[23] 朱大伟, 张洪程, 郭保卫, 等. 中国软米的发展及展望[J]. 扬州大学学报: 农业与生命科学版, 2015, 36(1): 47–52 ZHU D W, ZHANG H C, GUO B W, et al. The development and outlook of Chinese soft rice[J]. Journal of Yangzhou University: Agricultural and Life Science Edition, 2015, 36(1): 47–52
[24] 王才林, 张亚东, 朱镇, 等. 优良食味粳稻新品种南粳9108的选育与利用[J]. 江苏农业科学, 2013, 41(9): 86–88 WANG C L, ZHANG Y D, ZHU Z, et al. Breeding and utilization of the new japonica rice variety Nanjing 9108[J]. Jiangsu Agricultural Sciences, 2013, 41(9): 86–88
[25] 孙成效, 段彬伍, 谢黎虹, 等. 利用近红外透射光谱技术同步测定糙米的多项品质指标初报[J]. 中国水稻科学, 2006, 20(4): 451–454 SUN C X, DUAN B W, XIE L H, et al. Determination of several quality characteristics of brown rice by near infrared transmission spectroscopy[J]. Chinese Journal of Rice Science, 2006, 20(4): 451–454
[26] 杨连新, 王余龙, 石广跃, 等. 近地层高臭氧浓度对水稻生长发育影响研究进展[J]. 应用生态学报, 2008, 19(4): 901–910 YANG L X, WANG Y L, SHI G Y, et al. Responses of rice growth and development to elevated near-surface layer ozone (O3) concentration: A review[J]. Chinese Journal of Applied Ecology, 2008, 19(4): 901–910
[27] 郭贵华, 刘海艳, 李刚华, 等. ABA缓解水稻孕穗期干旱胁迫生理特性的分析[J]. 中国农业科学, 2014, 47(22): 4380–4391 GUO G H, LIU H Y, LI G H, et al. Analysis of physiological characteristics about ABA alleviating rice booting stage drought stress[J]. Scientia Agricultura Sinica, 2014, 47(22): 4380–4391
[28] 马一泓, 王术, 于佳禾, 等. 水稻生长对干旱胁迫的响应及抗旱性研究进展[J]. 种子, 2016, 35(7): 45–49 MA Y H, WANG S, YU J H, et al. Responses of drought stress in plant growth and study advances on drought resistance in rice[J]. Seed, 2016, 35(7): 45–49
[29] 杨连新, 王余龙, 董桂春, 等. 栽培和环境条件对水稻饱粒重的影响[J]. 江苏农业科学, 2002, 30(6): 9–13 YANG L X, WANG Y L, DONG G C, et al. Effects of different cultural and environmental conditions on fulfilled grain weight in rice[J]. Jiangsu Agricultural Sciences, 2002, 30(6): 9–13
[30] YANG L X, WANG Y L, DONG G C, et al. Effect of nitrogen on hull traits and its causes in Yangdao 6[J]. Agricultural Sciences in China, 2002, 1(7): 738–744
[31] 段俊, 田长恩, 梁承邺. 水稻结实过程中谷壳的作用及生理变化[J]. 作物学报, 2000, 26(1): 71–76 DUAN J, TIAN C E, LIANG C Y. Studies on effects and physiological changes of the hull on grain filling in rice[J]. Acta Agronomica Sinica, 2000, 26(1): 71–76
[32] 张启莉, 谢黎虹, 李仕贵, 等. 稻米蛋白质与蒸煮食味品质的关系研究进展[J]. 中国稻米, 2012, 18(4): 1–6 ZHANG Q L, XIE L H, LI S G, et al. Research progress of the relationship between rice protein and cooking and eating quality[J]. China Rice, 2012, 18(4): 1–6
[33] WANG Y X, FREI M, SONG Q L, et al. The impact of atmospheric CO2concentration enrichment on rice quality — A research review[J]. Acta Ecologica Sinica, 2011, 31(6): 277–282
[34] 李刚, 邓其明, 李双成, 等. 稻米淀粉RVA谱特征与品质性状的相关性[J]. 中国水稻科学, 2009, 23(1): 99–102 LI G, DENG Q M, LI S C, et al. Correlation analysis between RVA profile characteristics and quality in rice[J]. Chinese Journal of Rice Science, 2009, 23(1): 99–102
Influence of artificial lodging at grain-filling stage on plant growth, yield and quality of super rice*
ZHAO Xinyong1,2, SHAO Zaisheng1, WU Yanzhen1, ZHAO Yipeng2, WANG Yulong1, WANG Yunxia3, YANG Lianxin1**
(1. Jiangsu Key Laboratory of Crop Genetics and Physiology / Co-innovation Center for Modern Production Technology of Grain Crops, Yangzhou University, Yangzhou 225009, China; 2. Xuzhou Institute of Agricultural Sciences of Xuhuai District of Jiangsu Province, Xuzhou 221121, China; 3. College of Environmental Sciences and Engineering, Yangzhou University, Yangzhou 225009, China)
Rice lodging has been one of the main constraints to high yield and good quality rice production in China. Because lodging not only makes mechanized harvesting difficult, it also impedes assimilate, water and nutrient transport in grains, limiting the realization of rice yield potential. Lodging conditions under high humidity are prone to diseases and pests, resulting in poor grain quality. In recent years, the potential risk of rice lodging has been increasing because of the release of high-yielding varieties characterized by large panicle, increased nitrogen fertilizer application and shifts from labor-intensive to simplified planting techniques such as direct-seeding or seedling broadcasting. In addition, extreme weather conditions, such as typhoon accompanied with heavy rains at late growth stage, has been another main reason behind lodging, often affecting large areas of croplands. Therefore, it was of utmost importance to further study the mechanisms of lodging-resistance of rice and the related regulation strategies. In this experiment, the effect of artificial lodging at grain-filling stage on plant growth, yield and grain quality of super rice (‘Nanjing 9108’) was investigated by comparing plants with normal growth (CK), artificial lodging after 30 days of heading (AL30) and artificial lodging after 15 days of heading (AL15).Compared with CK, AL30 and AL15 artificial lodging treatments decreased dry matter production of aboveground organs of ‘Nanjing 9108’, which resulted in decrease in total dry matter production by 7.3% (< 0.05) for AL30 and 24.3% (< 0.01) for AL15. The proportion of biomass distribution in reproductive organs decreased for AL15, but had no significant change for AL30. AL30 and AL15 artificial lodging treatments decreased grain yield by 8.3% (< 0.05) and 36.4% (< 0.01), respectively. Yield loss caused by lodging at grain-filling stage was mainly associated with decreases in filled-grain rate and 1000-full-grain weight. Filled-grain rate and 1000-full-grain weight decreased by 5.6% (< 0.05) and 3.3% (< 0.05) for AL30, and by 30.8% (< 0.01) and 6.7% (< 0.01) for AL15. Lodging-induced decrease in grain weight was mainly associated with smaller brown rice, which was attributed to poorer grain filling at grain-filling stage. Head rice rate decreased by 7.5% (< 0.05) for AL30 and 14.7% (< 0.01) for AL15. AL30 treatment had no effect on protein or amylose contents of rice grain, but AL15 increased protein content by 20.2% (< 0.05). Artificial lodging had no significant effect on RVA profile of rice grain. The results suggested that the effect of plant lodging at late grain-filling stage (30 days after heading) on yield and grain quality of ‘Nanjing 9108’ super rice was relatively small. In contrast, plant lodging at early grain-filling stage (15 days after heading) inhibited dry matter production and grain-filling process, resulting in yield loss and grain quality deterioration.
Rice; Artificial lodging; Dry matter production and distribution; Grain yield; Yield component; Grain quality
, E-mail: lxyang@yzu.edu.cn
Dec. 20, 2017;
Mar. 22, 2018
S31
A
1671-3990(2018)07-0980-10
10.13930/j.cnki.cjea.171180
* 国家自然科学基金项目(31371563)和江苏高校优势学科建设工程资助
杨连新, 主要从事作物栽培、生理和生态方面的研究。E-mail: lxyang@yzu.edu.cn 赵新勇, 主要从事作物栽培学与耕作学研究。E-mail: 1151626431@qq.com
2017-12-20
2018-03-22
* This work was funded by the National Natural Science Foundation of China (31371563) and the Prior Discipline Construction of Jiangsu Higher Education Institutions.
赵新勇, 邵在胜, 吴艳珍, 赵轶鹏, 王余龙, 王云霞, 杨连新. 花后人为模拟倒伏对超级稻生长、产量和品质的影响[J]. 中国生态农业学报, 2018, 26(7): 980-989
ZHAO X Y, SHAO Z S, WU Y Z, ZHAO Y P, WANG Y L, WANG Y X, YANG L X. Influence of artificial lodging at grain-filling stage on plant growth, yield and quality of super rice[J]. Chinese Journal of Eco-Agriculture, 2018, 26(7): 980-989
猜你喜欢
湖南农业科学(2021年11期)2021-12-21
中国水稻科学(2021年5期)2021-09-13
中国稻米(2021年2期)2021-04-04
中国粮油学报(2019年8期)2019-09-11
阅读(快乐英语高年级)(2019年8期)2019-09-10
小火炬·智漫悦读(2018年12期)2018-03-13
莫愁(2017年36期)2017-12-25
亲子(2014年4期)2014-05-10
食品工业科技(2014年13期)2014-03-11
儿童故事画报(2013年3期)2013-06-24
