
黄河湿地包头段不同地被类型对土壤有机碳的影响
2018-07-04 01:59杨文焕王铭浩李卫平樊爱萍苗春林于玲红
生态环境学报 2018年6期
杨文焕,王铭浩,李卫平,樊爱萍,苗春林,于玲红*
土壤碳库一般分为易分解的活性有机碳和稳定的惰性有机碳(张俊华等,2012)。土壤碳库中的活性碳组分对环境因子变化响应最为敏感,虽然活性有机碳只占土壤有机碳总量的一小部分,却是反映土壤碳库周转的灵敏指标(万忠梅等,2011),也是最能反映土壤碳库稳定性的指标(李太魁等,2013;肖烨等,2015;张帅等,2015)。湿地由于长期处于淹水或水分过饱和状态,积累了更多的活性有机碳,对气候变化更为敏感(Dodla et al.,2012;Abril et al.,2014),因此,利用活性有机碳研究土壤碳库稳定性已成为研究热点(钟春棋等,2010)。张文敏等(2014)研究表明,在0~30 cm土层范围内活性有机碳含量占总有机碳含量的 32.74%~33.07%,互花米草(Spartina alterniflora)的入侵增强了滩涂湿地的固碳能力,有机碳活性组分能够反映有机碳库的变化。
土地利用与土地覆被变化是影响地球环境系统的最主要的人类活动方式(邰继承等,2011;房飞等,2013),它可以影响植被凋落物和残余量,影响土壤微生物的活动(王洪杰等,2003),引起土壤活性碳库的变化(Zou et al.,2005;Sun et al.,2011;赵鑫等,2006),且表层土壤有机碳对土地利用方式的响应和敏感程度极其显著。将湿地转变为耕地是常见的土地利用类型转变方式之一(Armentano et al.,1986;Bridgham et al.,2006),然而开垦却降低了湿地土壤有机碳含量和有机碳密度,碳库稳定性下降(钟春棋,2009)。目前,人类开垦活动的加剧,使得湿地成为受人类活动威胁最为严重的生态系统之一(Lemly et al.,2000),通过土地利用方式变化对湿地进行保护和管理成为重中之重,国内外关于土地利用方式对土壤有机碳的研究日趋增多(Yang et al.,2013;Bae et al.,2015;王丽丽等,2009;吴江琪等,2017),但大都集中在滨海湿地、沼泽湿地,而关于城市周边湿地的研究较少。
南海湿地为黄河湿地包头段重要组成,也是内蒙古重要湿地之一,2006年,南海湿地自然保护区被列入国家湿地保护工程规划的重点保护名录。南海湿地有黑鹳(Ciconia nigra)、遗鸥(Larus relictus)、大天鹅(Cygnus cygnuscygnus)等珍稀鸟类,在调蓄洪水、涵养水源、净化环境、促淤护岸、维护生物多样性等方面具有十分重要的作用。为了更好地保护包头南海湿地,满足生物多样性保护和区域社会经济可持续发展的需要,对南海湿地土地利用方式进行管理和对南海湿地生态系统进行保护,本研究以黄河湿地包头段——南海湿地土壤为研究对象,采集不同地被类型——裸地、芦苇(Phragmites australis)、旱柳(Salix matsudana)、玉米(Zea mays)、向日葵(Helianthus annuus)下的土壤样品,分析土壤有机碳组分的空间分布特征,探讨不同地被类型下土壤有机碳库稳定性及有机碳含量差异的原因,提出针对性的保护对策,旨在使南海湿地的功能效益得到提高。
1 材料与方法
1.1 研究区概况
黄河湿地包头段——南海湿地(40°30′8″N~40°33′32″N,109°59′2″E~110°2′26″E)位于包头市东河区南侧,黄河北岸,是黄河改道南移后形成的湖泊和滩涂,拥有开阔的水面和大面积的疏林草地(解一凡等,2006)。因特殊的地理位置、气候特征和水文地质条件,其成为了高纬度寒旱区独特的河流湿地生态系统(于玲红等,2016)。该湿地南部与黄河相接,相接部分的黄河段长度为7.0 km,东至东河槽,西与包头市高新技术开发区接壤(高静湉等,2017)。南海湿地总面积为2104.6 hm2,其中保护区面积为1664.4 hm2。南海湿地主体地貌为黄河冲积平原,平均海拔为1020 m,地势呈阶梯形,北高南低,保护区内的地表水主要来源于黄河水,其次为地下水和大气降水。该区域属于半干旱中温带大陆性季风气候,年平均气温为8.5 ℃,日照充足,光能资源丰富,年日照时数为3177 h;年降水量为307.4 mm,年水面蒸发量为2343 mm;全年无霜期为148 d。南海湿地土壤类型分为草甸土、盐土和风沙土三类。草甸土主要分布在湖区及外围的沼泽地带,为南海湿地面积最大的一个土类,盐土呈斑状块散布于沼泽的外围,风沙土大多分布于黄河岸边的沙滩地。南海湿地成土母质主要是黄土状沉积物及沙质风积物,还有冰碛物和冰水沉积物,土壤质地以细沙和粉砂为主(李卫平等,2017)。
1.2 取样点的布设
对包头南海湿地进行实地勘察,根据湿地植被区域划分(裸地、芦苇、玉米、向日葵、旱柳)进行样品采集。由于南海湿地水域较广,一些区域不能进行土样采集,故选取具有代表性的区域划分点位进行样品采集。其中,芦苇为自然生长,旱柳、向日葵和玉米均为人工种植,种植年限分别为 10 a、17 a和29 a,旱柳为不施肥状态,向日葵和玉米均为人工施肥。各取样点位置见图1。
各取样点布设说明见表1。其中,编号为1、2、3、6和10的点位分别为裸地、树林、玉米、芦苇和向日葵取样区域点;编号为 4、5、7、8和9的点位分别为树林、芦苇、玉米、裸地和向日葵的对照区域点位,每个区域点位设置3个平行样方。
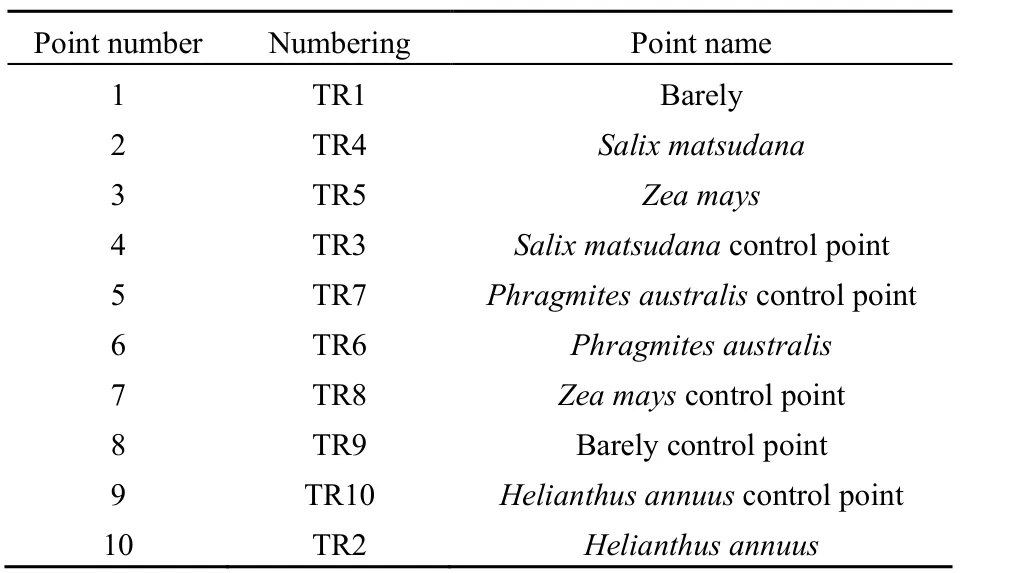
表1 包头南海湿地取样点布设说明Table 1 Baotou Nanhai wetland sampling site layout instructions
1.3 样品采集
由于湿地各类植被区域划分较散,每种地被类型选取一个区域进行取样恐不具有代表性,为了确保精确性,每种地被类型另选取1个对照点位进行取样,之后进行数据对比,如数据相差不大,再取平均值进行数据分析。每种地被类型研究区域内的每个取样点,根据实际情况选取3个2 m×2 m样方进行取样,共设置30个样方。在每个样方4个顶点及样方中心点上用土钻采集0~10、10~20、20~30、30~40、40~50、50~60 cm 共 6 个层次的土样,将同一样方的土样进行同层混合。土样装入自封袋中带回实验室自然风干,除去石块、动植物残体等杂物,研磨后用四分法取样并过100目(0.15 mm)筛备用。
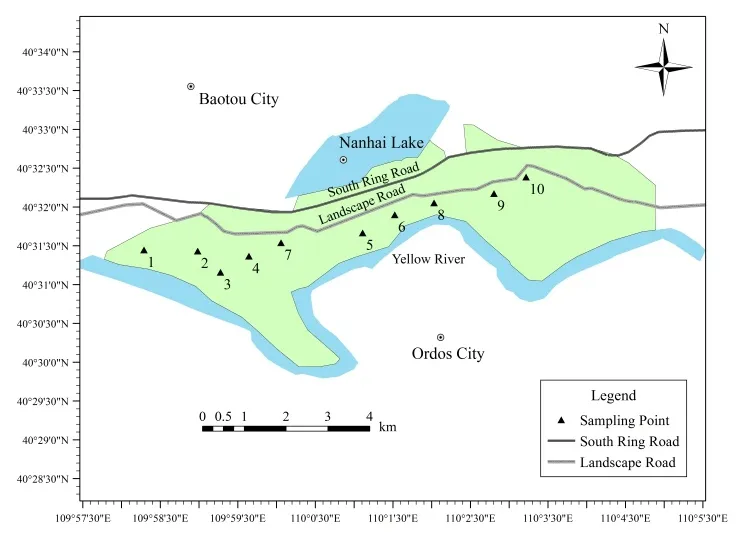
图1 包头南海湿地取样布点图Fig. 1 Baotou South China Sea wetland sampling layout
1.4 实验方法
有机碳总量采用重铬酸钾氧化法-硫酸外加热法测定(中国科学院南京土壤研究所,1978),活性有机碳含量采用333 mmol∙L-1高锰酸钾氧化法测定(Logninow et al.,1987;Lefroy et al.,1993),惰性有机碳取总有机碳与活性有机碳平均值的差值(张俊华等,2012)。
1.5 数据分析
运用Excel 2010对实验数据进行初步整理;运用Origin 9.0进行绘图;运用ArcGIS 10.2进行空间插值并制图。
2 结果与分析
2.1 土壤有机碳含量垂直分布特征
由表2可知,不同地被类型0~60 cm不同土层总有机碳含量的变异系数为14.42%~32.52%,平均为 19.68%;活性有机碳含量的变异系数为16.87%~30.26%,平均为 24.01%。根据雷志栋等(1985)和Nielsen et al(.1985)通过变异系数(CV)对空间变异性进行划分:CV<10%为弱变异性,10%~100%为中等变异性,CV>100%为强变异性。由此可知,南海湿地总有机碳和活性有机碳均属于中等变异。在0~10 cm层变异系数均为最大,表层受人为干扰和植被覆盖(枯枝烂叶腐烂分解使得有机碳含量增加)影响较大,随着土壤深度增加,变异系数逐渐减小,碳库稳定性越发趋于稳定。

表2 有机碳含量标准方差和变异系数Table 2 Standard deviation and coefficient of variation of organic carbon content
2.1.1 土壤总有机碳垂直分布特征
南海湿地0~10 cm、10~20 cm土层总有机碳含量较高,各地被类型有机碳含量峰值均出现在表层0~10 cm范围内,5种地被类型在30~60 cm的总有机碳含量均小于 10 g∙kg-1。除裸地有机碳含量从0~10 cm、10~20 cm下降趋势不太明显外,其他四类区域下降趋势都较大,呈现总有机碳含量随深度增加而减小的趋势(如图2),这种变化趋势与其他湿地土壤有机碳的研究结果(Mao et al.,2009)相似。王勇辉等(2016)研究发现艾比湖湿地7种主要类型土壤有机碳含量垂直分异规律很明显,即随剖面深度的增加,含量依次递减,但是减少的程度有所差异。其中盐化草甸覆盖下的土层自上而下,土壤有机碳含量减少的幅度最大,由 57.77 g∙kg-1减少到37.24 g∙kg-1;小乔木荒漠土壤有机碳含量由19.44 g∙kg-1减少到 11.93 g∙kg-1,其变化幅度较大;其他几种类型土壤有机碳含量随土层深度的增加而减少,其变化趋势基本一致,变幅不明显。

图2 5种地被类型下土壤总有机碳含量随深度的变化Fig. 2 Changes of soil total organic carbon (TOC) content with depth under five ground cover types
由图2可知,裸地、玉米、向日葵、旱柳、芦苇有机碳质量分数垂直变化范围分别为4.25~6.67、5.86~10.26、5.86~11.58、5.31~14.60、6.57~16.71 g∙kg-1。湿地开垦为不同地被类型后,玉米0~10 cm土层总有机碳含量降低39%,向日葵降低31%,旱柳降低13%,随着剖面深度的增加,差异变小,在其他分层无显著差异。这说明不同地被类型对湿地总有机碳含量的影响差别主要体现于 0~10 cm 土层。玉米和向日葵由于受人为耕作活动的扰动,有机碳含量在表层土(0~20 cm)中无显著差异。
2.1.2 土壤有机碳组分含量垂直分布特征
土壤碳库中的活性碳组分对环境因子变化响应最为敏感,作为能够反映土壤碳库稳定性的一个重要指标,活性有机碳是有效性较高、易被微生物分解利用部分有机质(Blair et al.,1995;Janzen et al.,1992),易受到外界干扰,不利于碳库的稳定;惰性有机碳的生物活性相对较低,很难被微生物利用,为稳定碳库。因此,把土壤有机碳分为总有机碳、活性有机碳和惰性有机碳来进行南海湿地土壤碳稳定性的研究。
由表3可知,南海湿地土壤活性有机碳质量分数在 1.22~6.17 g∙kg-1之间,平均值为 2.66 g∙kg-1,总有机碳质量分数在4.25~16.71 g∙kg-1之间,平均值为7.58 g∙kg-1,活性有机碳质量分数占总有机碳含量的30.73%~39.23%。张文敏等(2014)研究表明,在 0~30 cm 范围内,总有机碳质量分数在3.87~6.78 g∙kg-1之间,平均值为 5.33 g∙kg-1;活性有机碳质量分数在1.28~2.22 g∙kg-1之间,平均值为1.75 g∙kg-1,活性有机碳质量分数占总有机碳质量分数的32.74%~33.07%。对比可知,南海湿地与杭州湾湿地有机碳含量较高,但活性有机碳含量所占比重相差不大,碳库稳定性较为相似,属中等变异。黄昕琦等(2015)研究表明,在0~40 cm范围内总有机碳质量分数在2.55~16.00 g∙kg-1之间,平均值为 9.28 g∙kg-1;活性有机碳质量分数在 0.28~6.96 g∙kg-1之间,平均值为 3.62 g∙kg-1,活性有机碳质量分数占总有机碳质量分数的 10.98%~43.5%。南海湿地与乌梁素海湿地土壤总有机碳含量相差不大,但南海湿地活性有机碳整体所占比重较乌梁素海湿地大,碳库稳定性也相对较差。
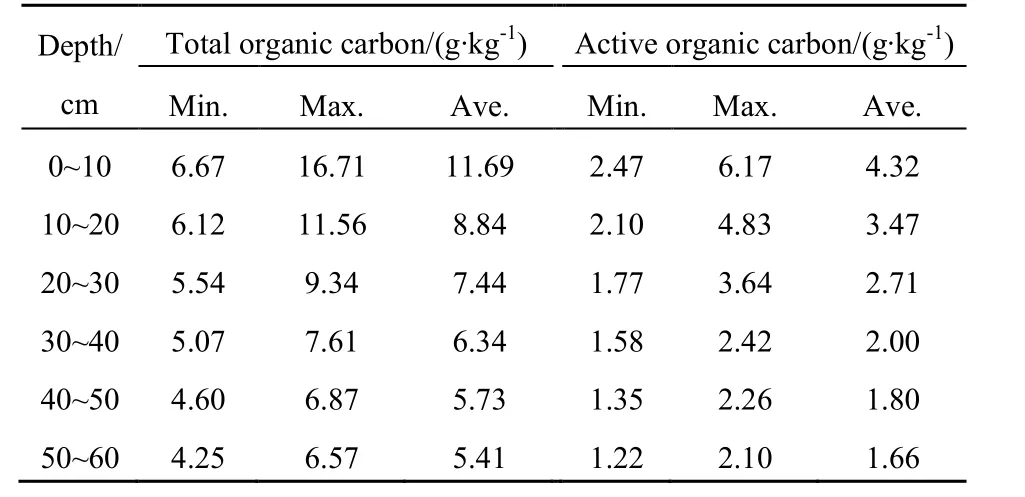
表3 总有机碳和活性有机碳含量Table 3 Total organic carbon and active organic carbon content
在不同土地利用方式下,由于植被类型不同,土壤有机物质数量和质量的返还程度不同,同时不同土地利用方式下,土壤环境差异较大,土壤有机碳的分解转化程度不同,因此土壤的活性有机碳也存在巨大差异。由图3可知,5种地被类型下,活性有机碳质量分数垂直变化范围分别为1.22~2.47、2.02~5.32、2.10~6.08、1.97~6.12、1.52~6.17 g∙kg-1,活性有机碳含量都要小于惰性有机碳含量,南海湿地碳库总体呈现稳定现象。
在不同土层,由于植被土壤环境、根系分布、生物活动、人工扰动等影响因子不同,不同类型土壤的活性有机碳在剖面的分布规律不同。本研究中,表层土壤活性有机碳含量明显高于中下层,0~10 cm层活性有机碳含量较大,各地被类型下活性有机碳质量分数范围为2.47~6.17 g∙kg-1,总有机碳质量分数范围为 6.67~16.71 g∙kg-1,活性有机碳质量分数占总有机碳含量的 36.91%~52.50%。其中,玉米和向日葵0~10 cm层活性有机碳含量最大,芦苇总有机碳含量较大,但活性有机碳含量所占的比例较少,低于裸地,因此,把湿地芦苇开发为玉米和向日葵降低了总有机碳含量,增加了活性有机碳所占比例,使得碳库趋于不稳定。而旱柳活性有机碳含量所占比重小于玉米和向日葵,又大于芦苇,同样也使得碳库趋于不稳定,但优于玉米和向日葵。由此可知,湿地芦苇碳库最为稳定。南海湿地北靠景观大道公路,南邻黄河,南海湿地土壤含水率较高,0~10 cm层土壤活性有机碳所占比例相对较高,玉米和向日葵尤其突出,受耕作影响,0~10 cm层活性有机碳含量高于惰性有机碳,随着植物生长,植物根系对碳进行吸收和稳定,随深度增加活性有机碳含量下降幅度很大,以惰性有机碳为主,碳库整体呈现稳定状态。

图3 5种地被类型下土壤有机碳组分含量随深度的变化Fig. 3 Changes in soil organic carbon composition with depth under five ground cover types
2.2 土壤有机碳含量水平分布特征
2.2.1 土壤总有机碳含量水平分布特征
由图4可知,0~10 cm层土壤总有机碳以芦苇(点位5和6)为最高,随着深度的增加,玉米、向日葵和旱柳土壤总有机碳含量逐渐与芦苇持平。湿地转化为耕地导致表层有机碳含量下降,向日葵总有机碳含量稍大于玉米,因向日葵区靠近路边且处于淹渍状态的时间长于玉米区,土壤内部缺氧或厌氧,有机碳分解缓慢,通过枝叶和根系等枯萎凋落向土壤中输入有机质(邵学新等,2011),使得向日葵总有机碳含量高于玉米。旱柳受人为影响相对较少,且表层土壤生有较多草类植物,其对土壤总有机碳也具有吸附作用,使得旱柳土壤总有机碳高于玉米区和向日葵区。裸地受人为踩踏影响较为严重,不利于土壤总有机碳积累(Bae et al.,2015),含量最低。
包头南海湿地与其他类型湿地相比,闽江口湿地、三江平原湿地和艾比湖湿地土壤总有机碳含量都高于包头南海湿地,原因是南海湿地土壤pH较高,植被种类和数量等生物量均少于其他类型湿地,有机碳的补充和累积有限(李卫平等,2015;金宝石等,2016)。另外,区域气候、成土年龄、土壤潜育化程度等存在差异,如泥炭沼泽广布的三江平原,因其特殊的形成过程,有机碳含量远高于其他湿地;青海湖高寒湿地低温条件下,有机碳分解困难等,也是造成包头南海湿地土壤有机碳低于其他类型湿地的原因(訾园园等,2016)。
2.2.2 土壤活性有机碳含量水平分布特征
图4 5种地被类型下土壤总有机碳含量水平分布Fig. 4 Distribution of total organic carbon (TOC) content in soil under five ground cover types
由图5可知,5种地被类型下活性有机碳质量分数在1.22~6.17 g∙kg-1之间,总有机碳质量分数在4.25~16.71 g∙kg-1之间,活性有机碳质量分数占总有机碳质量分数的30.73%~39.23%,整体以惰性有机碳为主,南海湿地土壤有机碳属稳定碳库。0~10 cm层除裸地外,其他四类地被类型土壤活性有机碳含量基本持平,随着深度增加,芦苇土壤活性有机碳含量减小幅度大于玉米、向日葵和旱柳,而土壤活性有机碳含量低于这三类土地利用方式区域,总体土壤活性有机碳含量表现为向日葵>玉米>旱柳>芦苇>裸地。活性有机碳占总有机碳含量的比例分别为:裸地 28.73%~37%,平均值为 32.06%;玉米34.53%~51.85%,平均值为42.41%;向日葵35.86%~52.5%,平均值为43.26%;旱柳31.22%~41.92%,平均值为35.08%;芦苇23.14%~36.91%,平均值为28.08%,表现为向日葵>玉米>旱柳>裸地>芦苇,向日葵和玉米平均值最高,碳库稳定性最低,芦苇碳库稳定性最高,故土壤有机碳稳定性表现为向日葵<玉米<旱柳<裸地<芦苇。
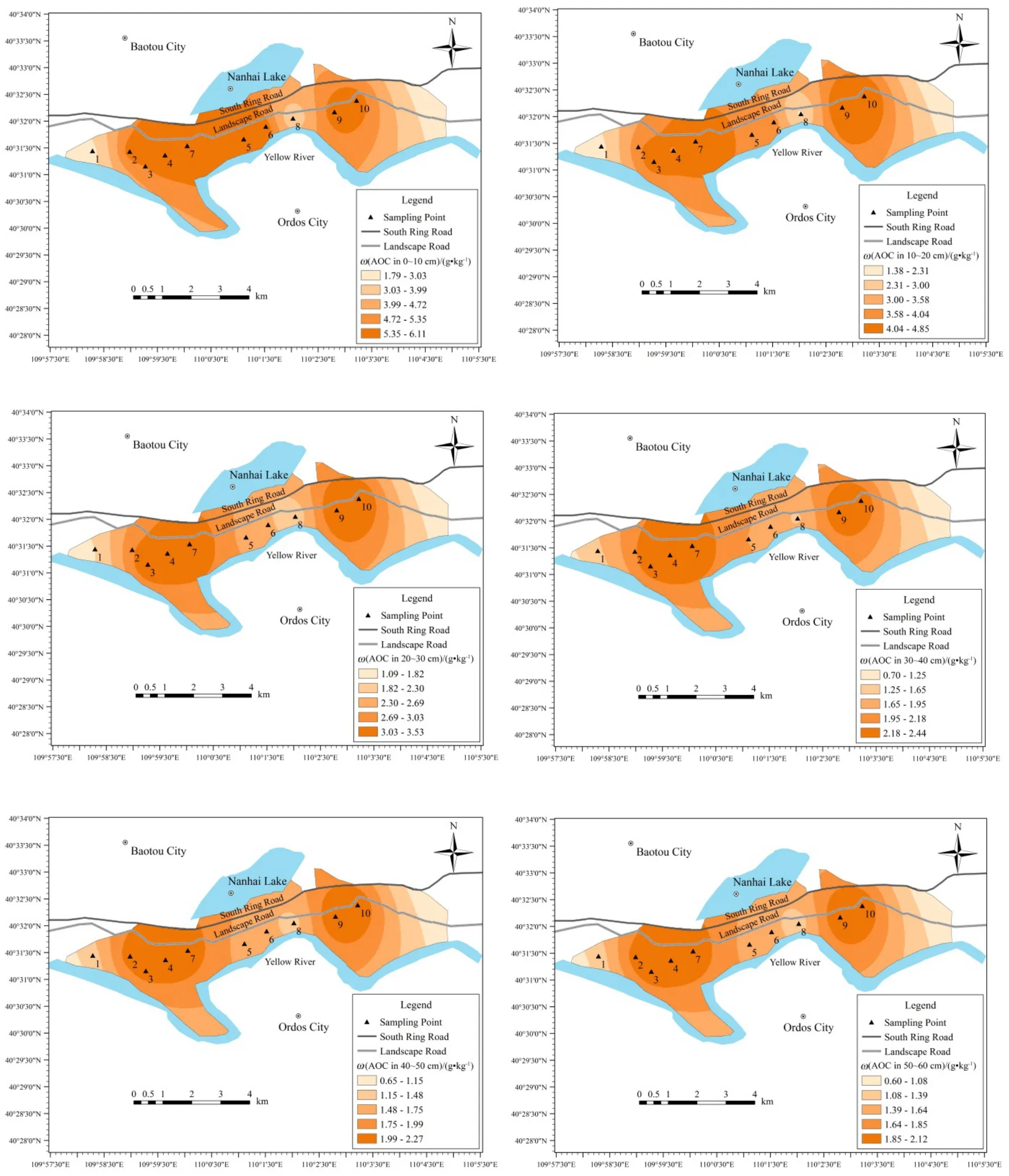
图5 5种地被类型下土壤活性有机碳含量水平分布Fig. 5 Distribution of soil active organic carbon (AOC) content in five land cover types
相比于玉米和向日葵,虽同属于耕作地区,但向日葵区含水率较高,土壤容重较低,故向日葵区表层土壤活性有机碳稍高于玉米区。旱柳受人为影响相对较少,且表层土壤生有较多草类植物,枯枝落叶的腐烂分解使得旱柳表层土壤活性有机碳含量较高,但随着深度增加,活性有机碳含量下降速度较大,其10~20 cm层活性有机碳含量小于玉米区和向日葵区。芦苇大面积种植且表层土壤芦苇残枝落叶较多,此外,芦苇含水量较高,更有利于微生物生长,故其表层土壤活性有机碳含量也较高,随着深度增加,芦苇根系对有机碳的吸附作用使得活性有机碳含量大幅降低,从10~20 cm层开始小于树林区。结合图 4可知,,芦苇区具有较高的总有机碳含量,较低的活性有机碳含量,说明种植芦苇对土壤碳库稳定性的影响高于其他几个区域,芦苇区土壤碳库稳定性较好;而旱柳土壤碳库稳定性高于玉米和向日葵,又低于芦苇,故旱柳对土壤碳库稳定性也有较大影响,但稍低于芦苇。
3 讨论
土壤有机碳在剖面上的垂直分布规律主要与植被凋落物和根系分布有关,地表大量的枯枝落叶,经腐殖质化及淋溶过程由表层向深层机械迁移。土壤表层是植被根系分布的重要区域(Liu et al.,2003;Kong et al.,2013),特别是旱柳和芦苇,表层凋落物较多,根系发达,植物根系随土壤深度增加而减少,死根腐解为土壤提供了丰富的碳源,因此旱柳和芦苇表层总有机碳含量较高,随着土壤深度增加,总有机碳含量下降幅度较大。旱柳因其大量的凋落物能够为表层土壤提供丰富的有机碳来源,另外,旱柳土壤没有耕作等人为干扰,土壤大团聚体受外力破碎的几率较低,使得大团聚体对有机碳的保护作用相对较强,有机碳含量较高(王浩等,2017)。芦苇种植密度高,植被覆盖率较大,植被凋落物及根系分泌物能够为土壤提供更多的有机质,土壤有机碳含量升高(覃灵华等,2016),此外,植被根系的分布直接影响了土壤有机碳的分布,因其大量的死根腐解,为土壤提供了丰富的碳源(Esteban et al.,2000),使得芦苇区总有机碳含量最高。
湿地转化为耕地不仅导致表层有机碳含量下降,而且使得表层有机碳变化幅度缩小,改变了表层有机碳的分布结构特征(陈伏生等,2004),这可能是耕作措施使下层土壤不断翻至表层造成稀释的结果(李月梅等,2005),导致土壤底层呼吸显著降低,有机碳含量下降(宫超等,2015)。湿地开垦为玉米和向日葵后,植物残体向土壤输入的有机碳减少(霍莉莉,2013),土壤总有机碳含量下降,虽然长期施肥能增加土壤有机碳含量,但南海湿地每年仅种一茬玉米和向日葵,施肥量较少,土壤有机碳含量增加较为有限(吴瑞娟等,2018)。另外,耕作导致有机碳矿化分解也是原因之一,耕作使得土壤团聚体结构受到破坏,导致包裹在团聚体中的有机质遭到人为破坏,加快了有机碳矿化分解速度,使土壤有机碳含量降低(李英等,2017),故玉米和向日葵土壤总有机碳含量低于旱柳和芦苇。
土壤活性有机碳主要来源于植物凋落物、土壤腐殖质和根系及根系分泌物,旱柳和芦苇因其表层凋落物及根系分泌物较多,使得表层活性有机碳含量较高,然而随着土壤深度增加,土壤容重增大,有机碳含量下降幅度较大,地下生物量也随之减少,因而土壤活性有机碳含量降低较为明显(沈玉娟等,2011);且旱柳和芦苇以凋落物输入土壤的有机碳除满足自身生长的需要外,大部分以稳定的惰性有机碳形态贮存下来(王义祥等,2014),故旱柳和芦苇活性有机碳占总有机碳含量比例较低。湿地开垦为玉米和向日葵后,因其翻耕和其他耕作措施改变了土壤温度、空隙状况和土壤微生物活性,相对旱柳和芦苇而言,玉米和向日葵土壤更加疏松,更适合微生物活动,加速了土壤总有机碳的分解,使其总有机碳含量下降,活性有机碳含量升高(唐国勇等,2010),故玉米和向日葵活性有机碳占总有机碳含量比例较高。
4 结论
(1)在垂直方向上,5种地被类型下湿地土壤有机碳组分含量随土壤深度增加而减小,不同地被类型对湿地有机碳含量的影响差别主要体现于0~10 cm层;各分层总有机碳和活性有机碳变异系数均在10%~100%之间,属中等变异,南海湿地碳库稳定性不高。
(2)旱柳和芦苇因其表层凋落物以及根系分泌物较多,总有机碳含量较高,随着土壤深度增加,根系的固碳和地下生物量减少,活性有机碳降低较为明显;而玉米和向日葵受耕作措施影响较大,土壤疏松,更适合微生物活动,加速了总有机碳的分解,使总有机碳含量下降,活性有机碳含量升高。在水平方向上,活性有机碳占总有机碳含量比例表现为向日葵>玉米>旱柳>裸地>芦苇,土壤有机碳稳定性表现为芦苇>裸地>旱柳>玉米>向日葵。
(3)人类活动使得南海湿地土壤总有机碳含量流失,活性有机碳比例增加,碳库稳定性下降。
ABRIL G, MARTINEZ J M, ARTIGAS L F, et al. 2014. Amazon River carbon dioxide outgassing fuelled by wetlands [J]. Nature, 505(7483): 395-398.
ARMENTANO T V, MENGES E S. 1986. Patterns of change in the carbon balance of organic soil-wetlands of the temperate zone [J]. Journal of Ecology, 74(3): 755-774.
BAE J, RYU Y. 2015. Land use and land cover changes explain spatial and temporal variations of the soil organic carbon stocks in a constructed urban park [J]. Landscape & Urban Planning, 136(136): 57-67.
BLAIR G J, LEFROY R, LISLE L. 1995. Soil carbon fractions based on their degree of oxidation, and the development of a carbon management index for agricultural systems [J]. Australian Journal of Agricultural Research, 46(7): 393-406.
BRIDGHAM S D, MEGONIGAL J P, KELLER J K, et al. 2006. The carbon balance of North American wetlands [J]. Wetlands, 26(4):889-916.
DODLA S K, WANG J J, DELAUNE R D. 2012. Characterization of labile organic carbon in coastal wetland soils of the Mississippi River deltaic plain: Relationships to carbon functionalities [J]. Science of the Total Environment, 435-436: 151-158.
ESTEBAN G. J, JACKSON R B. 2000. The vertical distribution of soil or ganic carbon and its relation to climate and vegetation [J]. Ecological Applications, 10(2): 423-436.
JANZEN H H, CAMPBELL C A, BRANDT S A, et al. 1992. Light-fraction organic matter in soils from long-term crop rotations [J]. Soil Science Society of America Journal, 56(6): 1799-1806.
KONG F L, XI M, LI Y, et al. 2013. Distribution and storage of DOC in a typical annular wetland of Sanjiang Plain [J]. Bulletin of Soil & Water Conservation, 33(5): 176-179.
LEFROY R D B, BLAIR G J, STRONG W M. 1993. Changes in soil organic matter with crop-ping asmeasured by organic carbon fractionsand 13C naturaliso-tope abundance [J]. Plant and Soil, 155(1):399-402.
LEMLY A D, KINGSFORD R T, THOMPSON J R. 2000. Irrigated agriculture and wildlife conservation: conflict on a global scale [J].Environmental Management, 25(5): 485-512.
LIU J S, YANG J S, YU J B, et al. 2003. Study on vertical distributon of soil organic carbon in Wetlands Sanjiang Plain [J]. Journal of Soil Water Conservation, 17(3): 5-8.
LOGNINOW W, WISNIEWSKI W, STRONY W M, et al. 1987.Fractionation of organic carbon based on susceptibility to oxidation[J]. Polish Journal of Soil Science, 20(1): 47-52.
MAO Z G, WANG G X, LIU J E, et al. 2009. [Influence of salt marsh vegetation on spatial distribution of soil carbon and nitrogen in Yancheng coastal wetland] [J]. Ying Yong Sheng Tai Xue Bao, 20(2):293-297.
NIELSEN D R, BOUMA J. 1985. Soil spatial variability: proceedings of a workshop of the ISSS and the SSSA, Las Vegas, USA, 30 November-1 December, 1984 [M]. Wageningen(Netherlands): Pudoc.
SUN B, HALLETT P D, CAUL S, et al. 2011. Distribution of soil carbon and microbial biomass in arable soils under different tillage regimes[J]. Plant & Soil, 338(1-2): 17-25.
YANG J S, LIU J S, HU X J, et al. 2013. Changes of soil organic carbon,nitrogen and phosphorus concentrations under different land uses in marshes of Sanjiang Plain [J]. Acta Ecologica Sinica, 33(6): 332-337.
ZOU X M, RUAN H H, FU Y, et al. 2005. Estimating soil labile organic carbon and potential turnover rates using a sequential fumigation-incubation procedure [J]. Soil Biology & Biochemistry,37(10): 1923-1928.
陈伏生, 曾德慧, 陈广生, 等. 2004. 开垦对草甸土有机碳的影响[J]. 土壤通报, 35(4): 413-419.
房飞, 唐海萍, 李滨勇. 2013. 不同土地利用方式对土壤有机碳及其组分影响研究[J]. 生态环境学报, 22(11): 1774-1779.
高静湉, 王晓云, 李卫平, 等. 2017. 包头南海湿地生态系统健康评价[J]. 湿地科学, 15(2): 207-213.
宫超, 宋长春, 谭稳稳, 等. 2015. 三江平原沼泽湿地垦殖对土壤微生物学性质影响研究[J]. 生态环境学报, 24(6): 972-977.
黄昕琦, 李琳, 吕烨, 等. 2015. 内蒙古乌梁素海湿地土壤有机碳组成与碳储量[J]. 湿地科学, 13(2): 252-257.
霍莉莉. 2013. 沼泽湿地垦殖前后土壤有机碳垂直分布及其稳定性特征研究[D]. 长春: 中国科学院研究生院(东北地理与农业生态研究所).
解一凡, 李建平, 董俊鲜. 2006. 南海子湿地对包头地区生态环境的作用[J]. 内蒙古林业, (10): 28-29.
金宝石, 闫鸿远, 张林海, 等. 2016. 中国滨海互花米草湿地土壤有机碳时空变化及其影响因素[J]. 生态环境学报, 25(12): 2021-2027.
雷志栋, 杨诗秀, 许志荣, 等. 1985. 土壤特性空间变异性初步研究[J].水利学报, (9): 12-23.
李太魁, 朱波, 王小国, 等. 2013. 土地利用方式对土壤活性有机碳含量影响的初步研究[J]. 土壤通报, 44(1): 46-51.
李卫平, 刘建龙, 鲍交琦, 等. 2015. 包头黄河湿地生态恢复植物类型的选择[J]. 湿地科学, 13(2): 211-216.
李卫平, 王非, 杨文焕, 等. 2017. 包头市南海湿地土壤重金属污染评价及来源解析[J]. 生态环境学报, 26(11): 1977-1984.
李英, 韩红艳, 王文娟, 等. 2017. 黄淮海平原不同土地利用方式对土壤有机碳及微生物呼吸的影响[J]. 生态环境学报, 26(1): 62-66.
李月梅, 王跃思, 曹广民, 等. 2005. 开垦对高寒草甸土壤有机碳影响的初步研究[J]. 地理科学进展, 24(6): 59-65.
邵学新, 杨文英, 吴明, 等. 2011. 杭州湾滨海湿地土壤有机碳含量及其分布格局[J]. 应用生态学报, 22(3): 658-664.
沈玉娟, 赵琦齐, 冯育青, 等. 2011. 太湖湖滨带土壤活性有机碳沿水分梯度的变化特征[J]. 生态学杂志, 30(6): 1119-1124.
邰继承, 靳振江, 崔立强, 等. 2011. 不同土地利用下湖北江汉平原湿地起源土壤有机碳组分的变化[J]. 水土保持学报, 25(6):124-128.
覃灵华, 徐祥明, 冷雄, 等. 2016. 赣南不同母质及植被覆盖红壤有机碳矿化研究[J]. 生态环境学报, 25(9): 1453-1460.
唐国勇, 李昆, 孙永玉, 等. 2010. 干热河谷不同利用方式下土壤活性有机碳含量及其分配特征[J]. 环境科学, 31(5): 1365-1371.
万忠梅, 郭岳, 郭跃东. 2011. 土地利用对湿地土壤活性有机碳的影响研究进展[J]. 生态环境学报, 20(3): 567-570.
王浩, 姚昕, 杨慧, 等. 2017. 岩溶区不同土地覆被方式对土壤团聚体有机碳的影响[J]. 生态环境学报, 26(9): 1506-1513.
王洪杰, 李宪文, 史学正, 等. 2003. 不同土地利用方式下土壤养分的分布及其与土壤颗粒组成关系[J]. 水土保持学报, 17(2): 44-46.
王丽丽, 宋长春, 葛瑞娟, 等. 2009. 三江平原湿地不同土地利用方式下土壤有机碳储量研究[J]. 中国环境科学, 29(6): 656-660.
王义祥, 叶菁, 王成己, 等. 2014. 不同经营年限对柑橘果园土壤有机碳及其组分的影响[J]. 生态环境学报, 23(10): 1574-1580.
王勇辉, 焦黎. 2016. 艾比湖湿地土壤有机碳及储量空间分布特征[J].生态学报, 36(18): 5893-5901.
吴江琪, 马维伟, 李广, 等. 2017. 尕海湿地沼泽化草甸中不同积水区土壤活性有机碳含量[J]. 湿地科学, 15(1): 137-143.
吴瑞娟, 王迎春, 朱平, 等. 2018. 长期施肥对东北中部春玉米农田土壤呼吸的影响[J]. 植物营养与肥料学报, 24(1): 44-52.
肖烨, 黄志刚, 武海涛, 等. 2015. 三江平原不同湿地类型土壤活性有机碳组分及含量差异[J]. 生态学报, 35(23): 7625-7633.
于玲红, 王晓云, 李卫平, 等. 2016. 包头南海湿地冰封期水质特征[J].湿地科学, 14(6): 810-815.
于荣, 徐明岗, 王伯仁. 2005. 土壤活性有机质测定方法的比较[J]. 中国土壤与肥料, (2): 49-52.
张俊华, 李国栋, 南忠仁, 等. 2012. 黑河绿洲区耕作影响下的土壤粒径分布及其与有机碳的关系[J]. 地理研究, 31(4): 608-618.
张帅, 许明祥, 张亚锋, 等. 2015. 黄土丘陵区土地利用变化对深层土壤活性碳组分的影响[J]. 环境科学, 36(2): 661-668.
张文敏, 吴明, 王蒙, 等. 2014. 杭州湾湿地不同植被类型下土壤有机碳及其组分分布特征[J]. 土壤学报, 51(6): 1351-1360.
赵鑫, 宇万太, 李建东, 等. 2006. 不同经营管理条件下土壤有机碳及其组分研究进展[J]. 应用生态学报, 17(11): 2203-2209.
中国科学院南京土壤研究所. 1978. 土壤理化分析[M]. 上海: 上海科学技术出版社.
钟春棋, 曾从盛, 仝川. 2010. 不同土地利用方式对闽江口湿地土壤活性有机碳的影响[J]. 亚热带资源与环境学报, 5(4): 64-70.
钟春棋. 2009. 土地利用变化对闽江口湿地土壤有机碳的影响研究[D].福州: 福建师范大学.
訾园园, 郗敏, 孔范龙, 等. 2016. 胶州湾滨海湿地土壤有机碳时空分布及储量[J]. 应用生态学报, 27(7): 2075-2083.
猜你喜欢
绿色科技(2020年20期)2020-11-20
天津师范大学学报(自然科学版)(2019年4期)2019-09-17
山东林业科技(2018年4期)2018-09-01
作文周刊·小学四年级版(2018年12期)2018-04-10
江苏农业科学(2017年6期)2017-05-11
林业勘查设计(2017年1期)2017-03-11
现代农业科技(2016年21期)2017-03-06
中国农业文摘-农业工程(2016年5期)2016-04-12
现代农业(2016年4期)2016-02-28
环境科技(2015年2期)2015-11-08
