
保护性耕作对蚕豆根际土壤微生物数量和酶活性的影响
2018-07-04 08:01:54黄召存王龙昌张小短马淑敏
干旱地区农业研究 2018年3期
黄召存,陈 娇,熊 瑛,2,王龙昌,张小短,邢 毅,马淑敏
(1. 西南大学农学与生物科技学院,南方山地农业教育部工程研究中心,三峡库区生态环境教育部重点实验室,重庆 400716;2. 河南科技大学农学院,河南 洛阳 471003)
我国西南地区旱作农田土层浅薄、水土流失严重,受季节性干旱影响严重,不利于农作物生长发育[1]。保护性耕作能改善土壤的物理化学性质,提高土壤质量[2-3]。土壤微生物群落结构组成及活性变化是衡量土壤质量、维持土壤肥力和作物生产力的一个重要指标[4]。土壤酶活性是土壤生物学活性的总体现,它表征了土壤的综合肥力特征及其变化状况、土壤养分的转化进程,可以作为评价土壤肥力水平的指标[5]。近年来,关于保护性耕作对西南地区旱作农田土壤影响的研究发现,垄作、秸秆覆盖等保护性耕作措施能有效控制水土流失[6]、提高农田水分利用效率和产量[7]、提高农田生态系统的碳汇能力[8-9]。然而对西南旱地保护性耕作措施下土壤微生物和酶活性的影响报道较少。本试验选择紫色土丘陵区蚕豆/玉米/甘薯旱三熟模式中的蚕豆为研究对象,比较不同耕作方式对蚕豆土壤微生物(细菌、真菌、放线菌)、酶活性(多酚氧化酶、转化酶、酸性磷酸酶)的影响以及它们之间的相关关系,旨在明确蚕豆与其根际土壤微生物和酶活性之间的相互作用,从而为蚕豆生产实践中选择合理的耕作方式奠定科学基础。
1 材料和方法
1.1 试验设计
试验地位于重庆市北碚区西南大学教学试验农场,北纬29°51′,东经106°27′,海拔244m,属亚热带季风湿润气候,年均太阳总辐射量为87108 kJ·cm-2,年均总日照时数1276.7 h,多年平均气温18℃,≥10℃积温5979.5℃,夏季最高气温达40℃左右,无霜期达359d,多年平均降雨量1133.7 mm。试验地土壤为旱地紫色土,坡度较缓,地力相对均匀。土壤容重1.21 g·cm-3,pH为6.47,土壤有机质含量28.00 g·kg-1,全氮含量1.68 g·kg-1,全磷含量1.46 g·kg-1,全钾含量34.54 g·kg-1,速效磷含量18.13 mg·kg-1,速效钾含量270.23 mg·kg-1,碱解氮含量35.23 mg·kg-1。
试验地已连续6年开展旱三熟种植模式下的垄作和秸秆覆盖保护性耕作研究,每年的耕作处理保持一致。于2013年11月至2014年5月以“蚕豆/玉米/甘薯”旱三熟种植模式中蚕豆农田为研究对象设置田间试验。其中参试作物为蚕豆(ViciafabaL.),品种为“陵西一寸”;于2013年11月初种植,2014年5月收获;蚕豆采取撬窝点播,种植在垄上,每条带2行,每行12窝,每窝3株,各处理均施复合肥225 kg·hm-2(含N15%、P2O515%、K2O15%),作为基肥在播种的同时施入。试验共设6个处理(如表1),采用随机区组排列,每个小区的面积为28.8 m2,长8.0m,宽3.6m。每个小区均分四厢共八个条带,每个条带宽度为1.0m,长度为3.6m。每个处理重复3次。覆盖处理所用的玉米秸秆,收获后直接覆盖于小区内,半量覆盖处理下的小区覆盖秸秆10.8kg(折合3750 kg·hm-2),全量覆盖处理下的小区覆盖秸秆21.6kg(折合7500 kg·hm-2)。田间管理措施同常规。
1.2 样品采集及指标测定
在蚕豆的苗期(2013年12月3日)、分枝期(2014年1月3日)、开花期(2014年2月3日)、结荚期(2014年3月6日)、成熟期(2014年5月1日),按照5点法取样,每个处理3个重复,以植株为中心,以15 cm为半径将整个植株从土壤中挖出,带回实验室后,抖掉与根系松散结合的土壤,然后用经火焰灭菌的镊子刮取附在根系上的一薄层(<10 mm)土壤作为根际土壤[10]。并将取得的土样弄碎混匀,按四分法取样,用无菌塑料袋包好,除去杂物后,取15~20 g根际土保存在 4 ℃冰箱内用于微生物分离、计数;其余部分自然风干,取一部分研磨,过1mm筛,测土壤酶活性。
细菌、放线菌和真菌数量均采用稀释平板法计数[11]。土壤转化酶活性(以葡萄糖质量计,mg·g-1,24h,37 ℃)采用3,5-二硝基水杨酸比色法[12],土壤多酚氧化酶(以紫色没食子素质量计,mg·g-1,24h,37 ℃),土壤酸性磷酸酶(以酚质量计,μmol·g-1,24h,37 ℃)则采用苏州科铭生物科技有限公司提供的土壤多酚氧化酶(PPO)及土壤酸性磷酸酶(S-ACP)试剂盒测定。
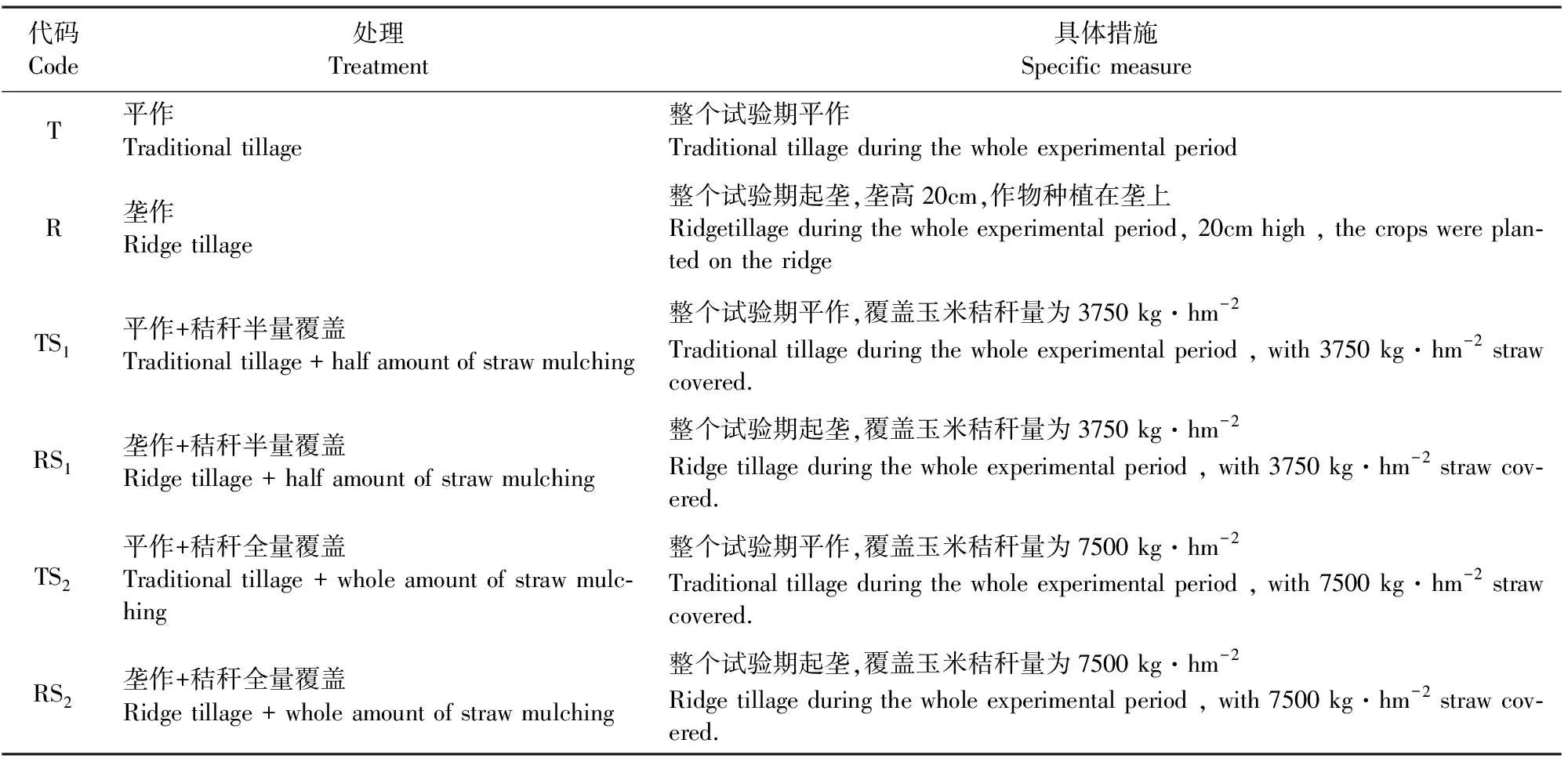
表1 试验处理描述
1.3 数据分析
用Excel 2007软件进行数据记录与处理,并制作图表,用SPSS 17.0软件进行显著性分析及相关性分析。
2 结果与分析
2.1 蚕豆根际土壤微生物数量变化
细菌、真菌、放线菌是构成土壤微生物的主要生物量,它们的类群组成和数量变化通常能反映出土壤生物活性水平,显示土壤中生命物质的旺盛程度。在蚕豆根际土壤微生物中,细菌居优势(1.80×105~22.93×105个/g),放线菌次之(1.15×105~4.39×104个/g),真菌最少(2.20×103~10.00×103个/g)(表2,表3,表4)。
由表2可知,在蚕豆的整个生育期间,其根际土壤细菌数量呈先增加后减少的趋势,在结荚期达到最大值;根际细菌数量在各生育期大致呈现RS2>TS2> RS1>TS1>R>T的趋势。在苗期, RS2、TS2、RS1处理的根际土壤细菌数量都显著高于T处理(P<0.05),分别是T处理的6.25倍、4.25倍和3.38倍;在分枝期,各处理间的趋势与苗期一致;其后,随着蚕豆的生长,各处理细菌数量迅速增加,在结荚鼓粒期达到最大值后到成熟期又急剧下降。结荚期,在RS2处理下的细菌数量达到最高值,是苗期其处理的1.83倍,达到22.93×105个/g, TS2、 RS1、TS1、R、T分别比苗期各提高了7.39倍、3.42倍、1.46倍、1.39倍、0.66倍,各处理间无显著差异;成熟期根际细菌数量在整个生育期内最小。除结荚期,TS2和RS2处理下的蚕豆根际土壤细菌数量均显著高于T、R处理(P<0.05); RS2和TS2、 RS1和TS1、R和T处理之间的根际土壤细菌数量差异不显著。说明全量秸秆覆盖能显著增加蚕豆根际土壤细菌数量,垄作也能增加,但是效果显著;全量秸秆覆盖下的垄作耕作方式对增加蚕豆根际土壤细菌数量效果最为明显。
真菌参与土壤中有机质的分解,对土壤腐殖质的合成、氨化作用以及团聚体的形成有促进作用,并直接影响土壤肥力。在不同生育时期,不同耕作方式对蚕豆根际土壤真菌的影响不同(表3)。以T为对照,R能增加蚕豆根际土壤真菌的数量,在苗期、分枝期、开花期、结荚期、成熟期分别增加15.2%,26.5%,3.8%,19.1%,27.3%,但差异均不显著。以T为对照,RS2、TS2能增加蚕豆根际土壤真菌的数量,RS2在苗期、分枝期、结荚期较T分别提高27.2%,69.4%,48.1%,但差异不显著,在开花期、成熟期分别提高156.4%,79.5%,差异达显著水平(P<0.05);TS2在苗期、分枝期、结荚期较T分别提高15.0%,52.0%,34.4%,差异未达显著水平,在开花期、成熟期分别提高126.9%,47.7%,差异达显著水平(P<0.05)。说明全量秸秆覆盖能增加蚕豆根际土壤细菌数量,在开花期和成熟期效果最为明显;垄作也一定程度能增加蚕豆根际土壤细菌数量,但是效果不显著。
放线菌参与土壤有机质的分解,是形成土壤腐殖质的最稳定的有机化合物,与土壤肥力以及植物病虫害防治有着非常紧密的关系。从苗期到成熟期,蚕豆根际土壤放线菌的数量先增加后减少,在结荚期达到最大值,到成熟期迅速减少(表4)。整个生育期蚕豆根际土壤放线菌数量大致呈现RS2>TS2> RS1>TS1>R>T的趋势,但不同生育期不同处理间的差异量有所不同。以T为对照,R能增加蚕豆根际土壤放线菌的数量,在苗期和成熟期分别增加56.6%和45.7%,差异达显著水平(P<0.05),在分枝期、开花期、结荚期分别增加11.3%、30.3%、9.1%,差异均不显著;RS2、TS2较T在各生育期均能显著增加根际土壤放线菌的数量(P<0.05),其中RS2处理下的根际土壤放线菌数量在各生育期较T均增加1倍以上,在分枝期增加约1.8倍。说明垄作和秸秆覆盖在整个生育期均能增加蚕豆根际土壤放线菌数量,其中秸秆覆盖效果更为显著。
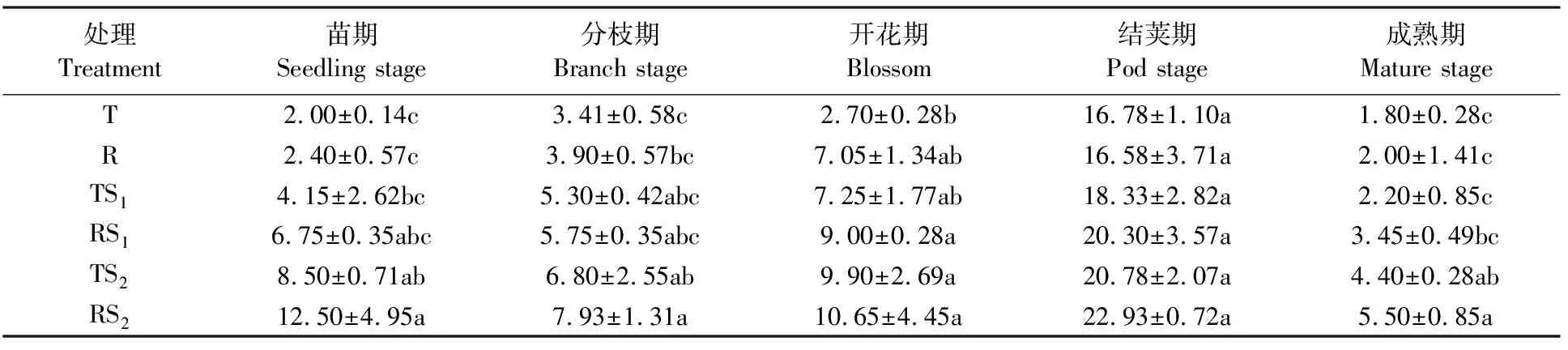
表2 不同耕作方式对蚕豆根际土壤细菌的影响
注: 同列中不同小写字母表示P<0.05水平差异显著.下同。
Notes: Different little letters show significant difference atP<0.05, the same as below.

表3 不同耕作方式对蚕豆根际土壤真菌的影响
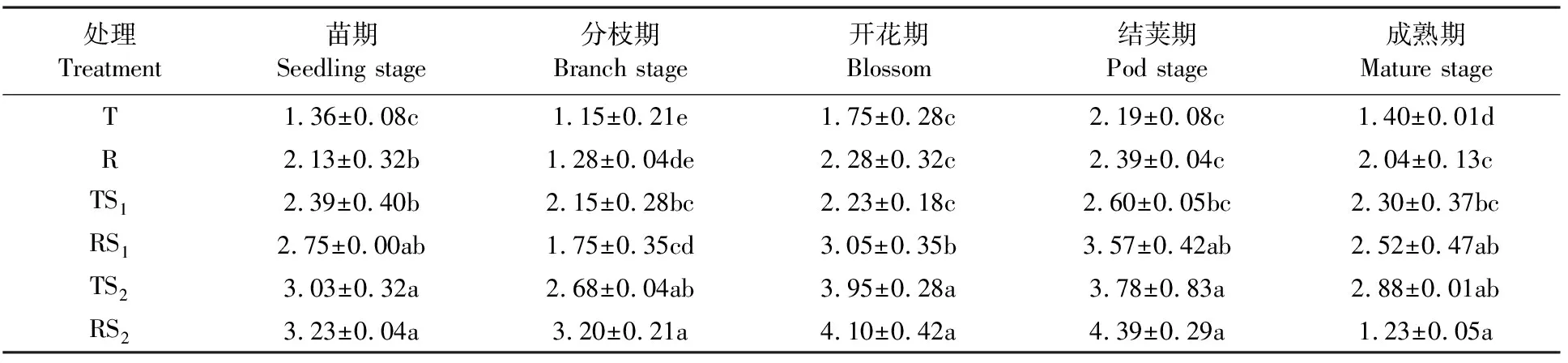
表4 不同生长时期蚕豆根际土壤放线菌数量的变化
2.2 蚕豆根际土壤酶活性的变化
土壤多酚氧化酶主要来源于土壤微生物、植物根系分泌物及动植物残体分解释放,催化土壤中芳香族化合物氧化成醌,醌与土壤中蛋白质、氨基酸、糖类、矿物等物质反应生成有机质和色素,完成土壤芳香族化合物循环,用于土壤环境修复。在蚕豆生长期内,不同处理土壤根际多酚氧化酶活性的变化趋势相似,苗期到分枝期多酚氧化酶活性下降,到开花期多酚氧化酶活性迅速升高,到结荚期出现高峰值,成熟期又迅速降低(表5)。在苗期、分枝期、结荚期、成熟期,RS2、TS2处理下的蚕豆根际多酚氧化酶活性分别显著高于处理R、T(P<0.05),R处理下的酶活性高于T,但是差异不显著。在开花期,RS2、TS2处理下的酶活性高于T,但只有RS2与T差异达到显著水平(P<0.05);R处理下的酶活性低于T,差异不显著。说明RS2、TS2能显著提高蚕豆根际多酚氧化酶的活性,TS1、RS1、R能一定程度提高蚕豆根际多酚氧化酶活性,但是效果不显著。
土壤转化酶能够水解蔗糖变成相应的单糖而被机体吸收,其酶促作用产物与土壤中有机质、氮、磷含量,微生物数量及土壤呼吸强度密切关,是评价土壤肥力的重要指标。由表6可知,不同处理下转化酶活性变化规律基本一致,即随着蚕豆生育时期的推进,各处理根际土壤转化酶活性从苗期到分枝期下降,到开花期后迅速上升,到结荚期出现高峰值,成熟期又迅速降低。在不同生育期,以T处理为对照,各处理均能提高蚕豆根际土壤转化酶的活性,但是不同处理之间的增加量有差异。在苗期,除了RS2处理与T处理达到显著性差异外(P<0.05),其它处理间无显著差异;在分枝期,各处理都和T处理达到了显著性差异(P<0.05),但其之间无显著差异;在开花期,RS2处理与T、R处理达到显著性差异(P<0.05),TS2、RS1处理的酶活性显著高于T处理(P<0.05);在结荚期,除了RS2处理与T、R处理达到显著性差异外(P<0.05),其它处理间无显著差异;成熟期,RS2、TS2的酶活性显著高于T、R、TS1、RS1的酶活性(P<0.05),其它处理间未达到显著性差异。说明在整个生育期,RS2、TS2、RS1、TS1、R处理均能提高蚕豆根际土壤转化酶的活性,其中RS2的效果最为显著。
土壤磷酸酶是一类催化土壤有机磷化合物矿化的酶,其活性的高低直接影响着土壤中有机磷的分解转化及其生物有效性,是评价土壤磷素生物转化方向与强度的指标。从表7可以看出,土壤根际酸性磷酸酶活性随着蚕豆生育期呈现出双峰趋势,在结荚期达到酶活性高峰。在蚕豆整个生育期,RS2、TS2、RS1、TS1、R处理下的蚕豆根际土壤酸性磷酸酶的活性均高于T,并呈现RS2> TS2,RS1> TS1,R>T,RS2>RS1>R, TS2>TS1>T的趋势。以T为对照,在苗期、分枝期和成熟期,各处理酶活性之间无显著性差异;在开花期和结荚期,RS2处理下的根际土壤酸性磷酸酶活性显著高于对照处理(P<0.05),比对照增加20.20%和21.80%,其它处理与对照无显著差异。说明RS2在开花期和结荚期能显著提高蚕豆根际土壤磷酸酶的活性,TS2、RS1、TS1、R也能提高蚕豆根际土壤磷酸酶的活性,但是效果不显著。
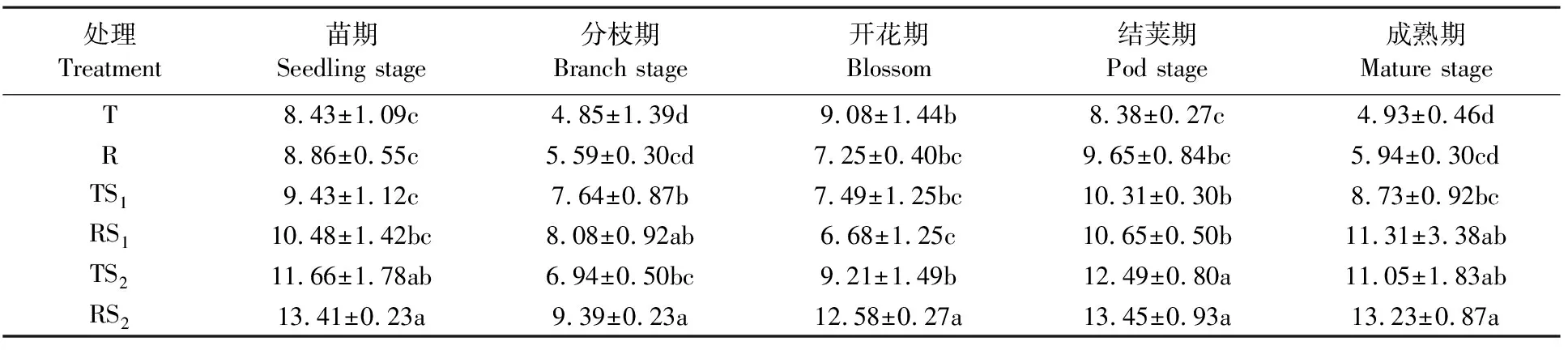
表5 根际土壤多酚氧化酶活性动态变化
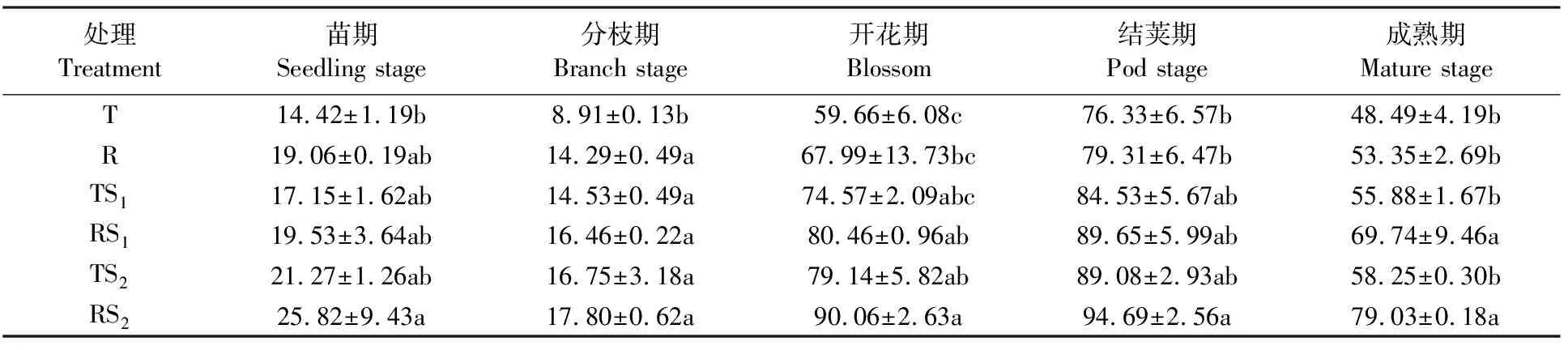
表6 根际土壤转化酶活性动态变化

表7 根际土壤酸性磷酸酶活性动态变化
2.3 土壤微生物数量和土壤酶活性相关性分析
土壤酶参与土壤的各种代谢过程和能量转化,与土壤的微生物数量有较好的相关关系。对土壤微生物数量与土壤酶活性进行皮尔逊相关性分析表明,两者存在一定的相关性(表8)。其中,细菌与酸性磷酸酶、转化酶、多酚氧化酶均呈极显著正相关;真菌与酸性磷酸酶、转化酶呈极显著正相关,与多酚氧化未达到显著相关;放线菌与酸性磷酸酶、转化酶、多酚氧化酶均呈极显著正相关。

表8 不同耕作方式下土壤微生物数量和土壤酶相关系数
注:**表示P<0.01 水平下极显著相关; *表示P<0.05 水平下显著相关。下同。
Note:**correlation is significant at the 0.01 level, and * correlation is significant at the 0.05 level. The same as below.
3 讨 论
土壤微生物的数量分布,不仅可以反映土壤环境质量的变化,而且也是土壤中生物活性的具体体现。本研究表明蚕豆根际土壤中微生物以细菌数量最多,其次为放线菌,再次为真菌。蚕豆农田土壤中细菌、真菌和放线菌的数量呈双峰变化趋势,在分枝期最低,在结荚期达到最大值。这是因为在分枝期时,土壤温度最低,消弱了微生物的活动;而进入结荚期,土壤温度比较适宜,植株的代谢活动比较活跃,微生物的活动随之增强;到成熟期,植株停止生长,根叶开始变枯衰老退化,不能为微生物的活动提供足够的养分。前人研究表明,垄作和秸秆覆盖能增加土壤微生物数量。其原因:一是因为垄作后地表微地形发生了变化,土表面积增大,土层处于浸润状态,土壤通透性好,保温性强,水、热、气均衡,有利于土壤微生物的繁育[14];二是因为秸秆覆盖为微生物提供了丰富的碳源,能够改善土壤物理性状,增加土壤腐殖质,提高土壤养分[15]。本研究表明垄作和秸秆覆盖相互作用下能增加土壤微生物数量,但是垄作效果并不显著,这是因为在我国西南地区,冬季和春季阴雨天气较多,太阳辐射较差,垄作保温效果差。
土壤酶总体活力大小在一定程度上可以反映土壤肥力大小[13],土壤生态系统的物质循环和能量转换与土壤酶活力紧密相关[14,15]。研究表明,作物不同耕作、施肥制度和种植方式等因素可引起土壤酶活力发生变化[16,17]。本研究发现蚕豆根际土壤中多酚氧化酶、转化酶、酸性磷酸酶的活性在整个生育期呈双峰变化趋势,在分枝期活性最低,结荚期活性最高。这主要是因为酶活性受温度和土壤性质的影响。土壤温度较低时酶活性较弱,土壤温度适宜时酶活性增强;到成熟期,微生物活动减少,酶的活性也降低。同时本研究结果表明,垄作和秸秆覆盖均能不同程度提高土壤酶活性,在垄作和秸秆覆盖同时作用下效果最为显著,且全量秸秆覆盖处理比半量秸秆覆盖处理活性更强。这是因为秸秆覆盖既对土壤温度有调节作用,也对土壤有机质和速效养分有较大影响,而且能够增加土壤微生物数量,增加了可供微生物转化的底物,所以土壤酶活性会增加[18]。李倩和贾志宽等[19]的研究结果认为,秸秆覆盖能提高土壤酶活性。蔡大义等[20]研究结果表明不同量秸秆覆盖提高了土壤养分含量和土壤酶活性。
土壤微生物作为生态系统的重要组成部分,在自然界营养物质循环中起着重要作用,同时土壤酶是土壤进行物质交换的中介,绝大多数土壤酶来源于土壤微生物,所以土壤酶、土壤微生物之间存在着一定的相关性[21-22]。本研究也发现,细菌与酸性磷酸酶、转化酶、多酚氧化酶均呈极显著正相关,真菌与酸性磷酸酶、转化酶呈极显著正相关,放线菌与酸性磷酸酶、转化酶、多酚氧化酶均呈极显正相关。这是因为微生物在其生命活动过程中释放分泌酶[23];在土壤酶的作用下,土壤有机物质和有机残体分解成不同的中间产物和最终产物,为微生物提供了营养物质和能量[24]。
4 结 论
在不同耕作方式下蚕豆根际土壤微生物中以细菌的数量最多,其次为放线菌,再次为真菌。不同微生物的数量在蚕豆各个生长阶段其变化规律存在差异,以平作为对照,各处理均能在不同程度上提高细菌、真菌、放线菌的数量,其中RS2效果最为显著。RS2处理下根际土壤细菌在苗期、分枝期、开花期、结荚期、成熟期较T处理分别提高525.00%、132.55%、294.44%、23.85%、175.00%,真菌较T处理分别提高27.00%、69.39%、156.41%、48.09%、79.55%,放线菌较T处理分别提高138.01%、178.26%、134.29%、100.23%、130.36%。垄作和秸秆覆盖能提高土壤各种酶的活性,其中RS2效果最为显著,RS2处理下根际土壤多酚氧化酶在苗期、分枝期、结荚期、成熟期较T处理分别提高38.34%、43.24%、1.44%、48.96%、123.89%,转化酶较T处理分别提高79.13%、99.66%、50.95%、24.06%、63.00%,酸性磷酸酶较T处理分别提高26.32%、22.65%、25.32%、21.81%、22.38%。土壤微生物数量与土壤酶均呈正相关,细菌、放线菌与酸性磷酸酶、转化酶、多酚氧化酶均呈极显著正相关,真菌与酸性磷酸酶、转化酶呈极显著正相关。因此选择垄作和全量秸秆覆盖耕作方式,有助于提高土壤微生物数量和酶活性,为蚕豆生长创造良好的微生态环境,对保持农田生态系统的可持续生产能力具有十分重要的意义。
参考文献:
[1] 王龙昌,谢小玉,张臻,等. 论西南季节性干旱区节水型农作制度的构建[J]. 西南大学学报(自然科学版), 2010, 32(2): 1-6.
[2] Zotarelli L, Alves B J R, Urquiaga S, et al. Impact of tillage and crop rotation on light fraction and intra-aggregate soil organic matter in two Oxisols[J]. Soil and Tillage Research, 2007, 95(1-2):196-206.
[3] Lopez-Fando C, Dorado J, Pardo M T. Effects of zone-tillage in rotation with no-tillage on soil properties and crop yields in a semiarid soil from central Spain[J]. Soil and Tillage Research, 2007, 95(1-2): 266-276.
[4] 刘淑琮,冯炘,于洁. 植物根际促生菌的研究进展及其环境作用[J]. 湖北农业科学, 2009, 48(11): 2882-2887.
[5] 曹慧,孙辉,杨浩,孙波,等. 土壤酶活性及其对土壤质量的指示研究进展[J]. 应用与环境生物学报,2003, 9(1):105-109.
[6] 杨勤,刘永红,柯国华,等. 坡耕地麦玉薯三熟保护性耕作水土保持效应的研究[J]. 西南农业学报, 2008, 21(2): 305-308.
[7] 邹聪明,胡小东,张云兰,等. 保护性耕作下西南旱三熟农田土壤水分动态及产量效应[J]. 水土保持通报, 2010, 30(6): 69-74.
[8] 张赛,王龙昌,黄召存,等. 保护性耕作下小麦田土壤呼吸及碳平衡研究[J]. 环境科学, 2014, 35(6): 2419-2425.
[9] 张赛,王龙昌,杜娟,等. 西南"旱三熟"区不同作物和秸秆覆盖对土壤团聚体及固碳潜力的影响[J]. 草业学报, 2016, 25(1):98-107.
[10] 胡小加. 根际微生物与植物营养[J]. 中国油料作物学报, 1999, 21(3): 77-79.
[11] 路超,李絮花,董静,等. 渗灌条件下果园覆盖的保水效果及对根际土壤养分和微生物特性的影响[J]. 水土保持学报,2013, 27(6):134-139.
[12] 关松荫,张德生,张志明. 土壤酶及其研究法[M]. 北京: 农业出版社, 1986, 274-339.
[13] 徐华勤,章家恩,冯丽芳,等. 广东省典型土壤类型和土地利用对土壤酶活性的影响[J]. 植物营养与肥料学报, 2010, 16(6): 1464-471.
[14] Li C. Development of deer antler model for biomedical research[J]. Recent Adv Res Updat, 2003, 4: 256-274.
[15] Li C, Yang F, Sheppard A. Adult stem cells and mammalian epimorphic regeneration insights from studying annual renewal of deer antlers[J]. Current Stem Cell Research & Therapy, 2009, 4(3): 237-251.
[16] 张星杰,刘景辉,李立军,等. 保护性耕作对旱作玉米土壤微生物和酶活性的影响[J]. 玉米科学, 2008, 16(1): 91-95.
[17] Yun W, Bin H, Zhongqiang S. Effects of conservation tillage on soil microbial characters and soil enzyme activites[J]. Journal of Soil and Water Conservation, 2006, 20(4): 120-123.
[18] 张彬,何红波,白震,等. 保护性耕作对土壤微生物特性和酶活性的影响[J]. 土壤通报,2010,41(4):230-236.
[19] 李倩,张睿,贾志宽. 玉米旱作栽培条件下不同秸秆覆盖量对土壤酶活性的影响[J]. 干旱地区农业研究, 2009,27(4):152-154.
[20] 蔡太义,贾志宽,黄耀威,等. 中国旱作农区不同量秸秆覆盖综合效应研究进展Ⅰ. 不同量秸秆覆盖的农田生态环境效应[J]. 干旱地区农业研究, 2012, 30(5): 63-68.
[21] 周礼恺,张志明,曹承绵. 土壤酶活性的总体在评价土壤肥力水平中的作用[J]. 土壤学报, 1983, 20(4): 413-417.
[22] 邱莉萍,刘军,王益权,等. 土壤酶活性与土壤肥力的关系研究[J]. 植物营养与肥料学报, 2004,10(3): 277-280.
[23] 林大仪.土壤学(第3版)[M].北京: 中国林业出版社, 2002:70-72.
[24] 周礼恺.土壤酶学[M].北京:科学出版社,1987:167-170.
猜你喜欢
今日农业(2021年15期)2021-10-14 08:20:24
今日农业(2021年11期)2021-08-13 08:53:36
国际呼吸杂志(2019年22期)2019-12-09 09:20:36
农家科技中旬版(2018年5期)2018-07-14 01:58:02
中国组织化学与细胞化学杂志(2017年1期)2017-06-15 20:27:45
广西林业科学(2016年1期)2016-03-20 05:33:00
医学研究杂志(2015年8期)2015-06-22 14:00:57
应用海洋学学报(2014年4期)2014-11-22 07:43:54
天然产物研究与开发(2014年6期)2014-04-27 14:15:56
中国医学科学院学报(2013年6期)2013-03-11 20:26:04
