
不同生境下的空心莲子草对南方菟丝子寄生的响应
2018-07-02 03:06郭素民高芳磊李钧敏
杂草学报 2018年1期
张 雪, 郭素民, 高芳磊, 闫 明, 李钧敏
(1.上海师范大学生命与环境科学学院,上海 200234; 2.台州学院/浙江省植物进化生态学与保护重点实验室,浙江台州 318000;3.山西师范大学生命科学学院,山西临汾 041004; 4.北京林业大学自然保护区学院,北京 100083)
生物入侵是指某些生物物种通过人类活动等传播到自然分布区以外的地区,这些物种被称为外来入侵物种或入侵物种[1],与物种的自然扩散不同。近年来,由于经济全球化,生物入侵现象更加恶劣,严重破坏了生态系统的稳定性及物种多样性,而且每年造成巨额经济损失[2]。生物入侵早已成为全球性环境问题,如何解决这一问题已成为生态学领域的热点研究。防治入侵植物的主要的方法有机械法、化学法和生物防治法[3]。其中生物防治方法,即利用入侵物种的天敌来控制或减缓入侵生物的扩散及生长[3],因为其绿色、生态友好等优势越来越受到生态学家的青睐。在入侵地引入入侵物种原产地的天敌,存在天敌可能扩散成为一个新入侵种的风险,因此,如何在入侵地寻找合适的天敌用于入侵植物防治受到了越来越多的关注[4-5]。菟丝子属(Cuscuta)为旋花科(Convolvulaceae)的全寄生草本植物。研究发现,菟丝子属植物偏好入侵植物,它们作为入侵植物的本地天敌有望成为一种新型生物防治剂,用于控制入侵植物的生长[4-5]。
植物的生长与防御之间往往存在着权衡[6]。植物在逆境环境下,如受到温度、水分、养分等非生物因子的胁迫,或受到食草动物、昆虫等生物因子的伤害时,将会通过减少对自身生长的投资,而加大对防御能力的投资,如增加防御物质等[6]。郭素民等野外采集南方菟丝子寄生的空心莲子草与无寄生的空心莲子草,经对比发现空心莲子草受到南方菟丝子寄生胁迫后,改变自身的生长-防御策略,减少营养生长投入而将更多的资源投向克隆繁殖,同时增强对“防御”物质的投入,增强其防御能力,以利于后代生存和繁衍[7]。
空心莲子草(Alternantheraphiloxeroides)为苋科(Amaranthaceae)莲子草属(Alternanthera)多年生草本植物,原产于南美洲,分布在巴拉圭、阿根廷南部巴拉那河流域和巴西南部沿海地带,20 世纪30 年代末空心莲子草作为饲料被引入中国[8]。由于它们具有极强的无性繁殖能力、抗逆能力与适应性,以及在入侵环境中缺乏专一的自然天敌的控制,引入不久便大肆蔓延,对入侵地的生态环境造成严重的威胁并带来一定的经济损失,是危害性极大的入侵物种,被列为中国首批外来入侵物种[9]。空心莲子草在原产地为水生生长[10],入侵我国后,逐渐可在陆地生长。马瑞燕等研究发现,引入中国的原产地专性天敌对水生生境的空心莲子草种群有显著的抑制作用,但对陆生生境空心莲子草种群影响却不显著,表明生境可以显著影响空心莲子草与其天敌之间的相互作用[11]。但不同生境是否会影响空心莲子草响应南方菟丝子寄生引起的生长与防御的权衡,目前仍是未知的。
王如魁等发现,野外南方菟丝子作为浙江省台州市本地新天敌可天然寄生于空心莲子草上,并且能够抑制空心莲子草的生长,促进本地植物的生长和群落的恢复[4]。本研究以空心莲子草-南方菟丝子为研究对象,分析浙江省台州市本地新天敌南方菟丝子寄生对不同生境空心莲子草生长与防御的影响。由于水生生境的空心莲子草未见有南方菟丝子寄生,因此,笔者采集陆生型与水陆两栖型空心莲子草为研究对象,比较分析其响应南方菟丝子寄生的生物量和次生代谢产物含量的变化,以阐明以下问题:(1)不同生境下南方菟丝子寄生对空心莲子草的生物量及其分配是否存在影响?(2)不同生境下南方菟丝子寄生对空心莲子草的次生代谢产物含量是否存在影响?(3)生境是否影响南方菟丝子寄生对空心莲子草生长-化学防御权衡关系?本研究结果不仅对于阐明入侵植物的入侵机制具有重要意义,还可为入侵植物生物防治开展的可行性研究提供理论依据。
1 材料与方法
1.1 试验基质的准备
将泥炭土、干净的河沙及蛭石按照体积比 6 ∶3 ∶1 的比例混匀,备用。装入体积为100 mL的一次性塑料杯(直径为4.5 cm,高为6 cm)中,每杯装基质100 mL,并对杯中的基质施肥,施肥标准为 0.08 g/杯,用于喜旱莲子草个体培养试验。
1.2 植物材料的准备及试验处理
于 2013年5月初,分别从浙江省临海市台州学院郊外农田和河道消落带中采集陆生和水陆两栖型2种不同生态型的空心莲子草,每个生态型采集20个个体,个体间隔3 m以上。从采集的空心莲子草上剪取长度粗细一致的茎段,扦插于上述已配制好的基质中,每杯1根茎段,每种生态型扦插20盆,共40盆,放入步入式人工智能气候室内进行培养,如下设置:白天温度32 ℃,相对湿度75%,14 h,夜晚温度25 ℃,相对湿度75%,10 h,定期进行灭菌和换气。其中,陆生型正常浇水,水陆两栖型一天置水中,一天正常浇水。待空心莲子草长到8 cm左右,选取长度为10 cm左右的菟丝子缠绕在空心莲子草的茎上进行寄生处理。
1.3 指标测定
试验持续60 d后,将空心莲子草植株分为根、茎、叶各部位,105 ℃杀青0.5 h后,于70 ℃烘箱烘干至恒质量,采用电子天平(精确至0.000 1 g)称取根、茎、叶各部分的生物量,并计算总生物量。用高速研磨仪将烘干的空心莲子草的茎研磨成粉末,过0.25 mm筛,用于4种次生代谢产物含量的测定。采用浓硫酸法测定木质素含量[12];以没食子酸为标准品,采用高锰酸钾测定法测定单宁含量[13];以没食子酸为标准品,采用紫外分光光度法测定总酚含量[14];采用苯酚-硫酸法测定总糖含量[15]。
1.4 数据处理
数据采用“平均值±标准差”的形式。采用双因素方差分析(two-way ANOVA)对2种生境空心莲子草在南方菟丝子寄生下各指标的差异进行显著性检验。数据分析采用SPSS 22.0软件,作图采用Sigmaplot 12.5软件。
2 结果与分析
2.1 南方菟丝子寄生对生物量及分配的影响
由图1和表1可知,南方菟丝子寄生处理极显著降低了2种生境下空心莲子草的根生物量,而对茎、叶和总生物量影响不显著,但是茎生物量及总生物量有明显下降趋势,叶生物量有上升趋势;陆生生境中喜旱莲子草根、茎、叶和总生物量极显著高于水陆两栖生境。寄生处理与不同生境间不存在显著的相互作用。
由图2和表2可知,寄生处理极显著降低了空心莲子草根冠比和根生物量分配比例,而极显著提高了叶生物量分配比例,对茎生物量分配比例影响不显著;无论是在水生生境下还是在水陆两栖生境下,空心莲子草根冠比、根生物量分配比例、茎生物量分配比例和叶生物量分配比例差异均不显著。与陆生生境相比,在水陆两栖生境下寄生处理明显降低了空心莲子草的根冠比和根生物量分配比例;在陆生生境下寄生处理降低了茎生物量分配比例,然而在水陆两栖生境下寄生处理提高了茎生物量分配比例。
2.2 南方菟丝子寄生对次生代谢产物的影响
由图3和表3可知,南方菟丝子寄生极显著提高了2种生态型空心莲子草的总酚含量,对单宁和木质素含量影响不显著;单宁含量在陆生生境下极显著高于水陆两栖生境,总酚和木质素含量差异不显著。寄生处理后水陆两栖型单宁含量及总酚含量都有所升高,较陆生型空心莲子草变化更为明显,木质素含量下降较陆生型下降趋势低。


表1 寄生处理和生境对空心莲子草生物量影响的双因素方差分析结果Table 1 Two-way ANOVAs results of the effects of habitats and parasitism on the A. philoxeroides biomass
注:“**”表示影响极显著(P<0.01);“*”表示影响显著(P<0.05)。下表同。
3 讨论
菟丝子属全寄生植物可以显著抑制寄主植物的生长,尤其是偏好入侵植物[5]。本研究发现,南方菟丝子的寄生均可以极显著抑制陆生生境与水陆两栖生境的空心莲子草的生长,显著改变其生物量分配。与大量的研究结果相似,菟丝子属植物的寄生可以显著抑制入侵的寄主植物的生长及生长策略[4-5,16]。
平衡生长假说认为植物在适应环境变化的时候,可通过调节各器官中的生物量分配,来最大化地获取水分、营养和光等受限资源,从而提高植物的生长速率[17-20]。本研究表明,在受到南方菟丝子寄生胁迫时,2种生态型空心莲子草的生物量差异不明显,但根冠比和根生物量分配比例极显著变化。其中,2种生态型的空心莲子草的叶生物量均有增加,根冠比及根生物量下降,虽然并不显著,但也说明在受到寄生胁迫后,通过叶生物量的增加,提高光合作用能力,更有利于生存,而通过降低根生物量来减缓生长速率[21]。其中水陆两栖型最为明显,陆生型变化程度较小。造成这种现象的原因主要是生境的差异,水陆两栖型与陆生型空心莲子草生存环境比较可知,水陆两栖型具有充足的水分,但氧气含量较陆生型低,而空心莲子草的茎具有发达的通气组织,并且为主要的繁殖器官。所以水陆两栖型通过提高茎生物量分配比例使自己占据更多的生存空间,降低根生物量分配比例,将更多的资源用于吸收氧气和光合作用上。而陆生型空心莲子草的根冠比、根生物量及茎生物量均有所下降,虽然变化不明显,但说明在受到寄生胁迫后陆生型空心莲子草减少了生长资源的投入。本研究表明,在生物量分配上,陆生型与水陆两栖型变化程度不同,但2种生态型的空心莲子草对寄生胁迫有相似的响应。
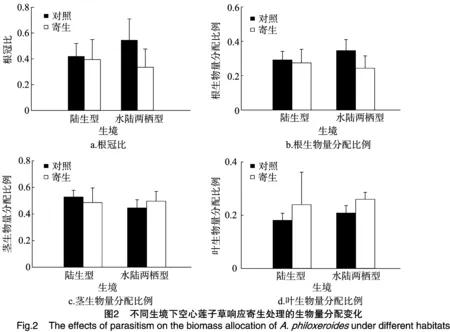

表2 寄生处理和生境对空心莲子草生物量分配影响的双因素方差分析结果Table 2 Two-way ANOVAs results of the effects of habitats and parasitism on the biomass allocation of A. philoxeroides
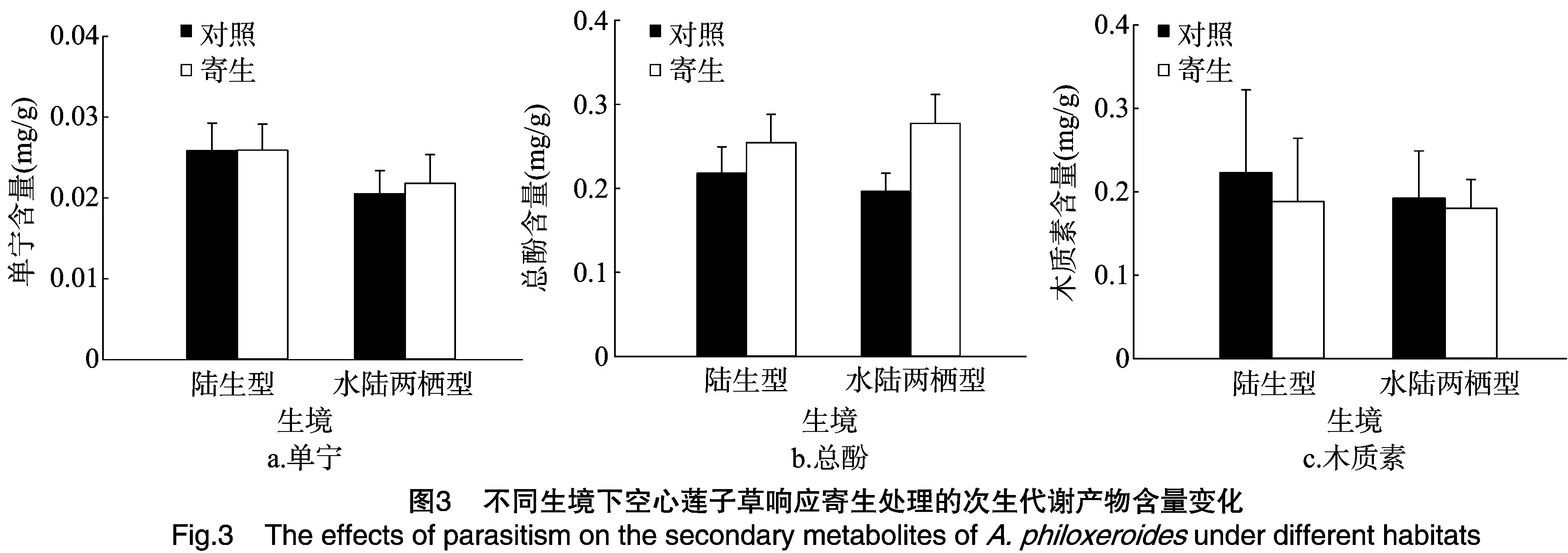

表3 寄生处理和生境对空心莲子草次生代谢产物含量影响的双因素方差分析结果Table 3 Two-way ANOVAs results of the effects of habitats and parasitism on the secondary metabolites of A.philoxeroides
次生代谢产物主要是由植物受到外界环境的影响而产生的,是植物的主要防御机制之一[22]。在本研究中,除木质素下降外,2种生态型空心莲子草的单宁和总酚含量有明显上升(陆生型不明显),其中水陆两栖型较陆生型更为明显。说明空心莲子草在受到南方菟丝子寄生胁迫后产生了一定的防御机制,提高了对菟丝子的抵抗能力。木质素主要存在于植物的茎部,具有支撑作用,本研究发现陆生型空心莲子草的木质素下降程度较水陆两栖型更为明显,说明寄生胁迫对陆生型空心莲子草的支撑作用有很大影响,这与陆生型空心莲子草茎生物量分配比例下降结果相一致。而2种生态型防御高低的不同与它们所生存的环境有很大关系,这与郭素民所研究的空心莲子草在受到胁迫后会通过次生代谢产物的变化提高自身的防御结论相一致[23],但其作用机制还需进一步研究。
本研究表明,不同生态型的空心莲子草在受到南方菟丝子寄生胁迫时,生长与防御的能量分配有相似的变化,但由于生境的差异,物质分配的量有所不同。结果表明,浙江省台州市新天敌南方菟丝子可以抑制空心莲子草的生长,使更多的资源转向防御,对减慢空心莲子草入侵扩散速度的研究具有重要的意义。但如何能够让菟丝子有效地抑制空心莲子草而不会对生态环境造成其他影响还需进一步研究。
参考文献:
[1]Lee C E. Evolutionary genetics of invasive species[J]. Trends in Ecology and Evolution,2002,17(8):386-391.
[2]徐承远,张文驹,卢宝荣,等. 生物入侵基质研究进展[J]. 生物多样性,2001,9(4):430-438.
[3]谢宗强,陈志刚,樊大勇,等. 生物入侵的危害与防治对策[J]. 应用生态学报,2003,14(10):1795-1798.
[4]王如魁,管 铭,李永慧,等. 南方菟丝子寄生对喜旱莲子草生长及群落多样性的影响[J]. 生态学报,2012,32(6):1917-1923.
[5]Li J M,Jin Z X,Song W J. Do native parasitic plants cause more damage to exotic invasive hosts than native non-invasive hosts? An implication for biocontrol[J]. PLoS ONE,2012,7(4):e34577.
[6]阎秀峰,王 洋,李一蒙. 植物次生代谢及其与环境的关系[J]. 生态学报,2007,27(6):2554-2562.
[7]郭素民,李钧敏,李永慧,等. 空心莲子草响应南方菟丝子寄生的生长-防御权衡. 生态学报,2014,34(17):4866-4873.
[8]郭连金,徐卫红,孙海玲,等. 空心莲子草入侵对乡土植物群落组成及植物多样性的影响[J]. 草业科学,2009,26(7):137-142.
[9]翁伯琦,林 嵩,王义祥. 空心莲子草在我国的适应性及入侵机制[J]. 生态学报,2006,26(7):2373-2381.
[10]潘晓云,耿宇鹏,Alejandro,等. 入侵植物喜旱莲子草——生物学、生态学及管理[J]. 植物分类学报,2007,45(6):884-900.
[11]马瑞燕,王 韧,丁建清. 利用传统生物防治控制外来杂草的入侵[J]. 生态学报,2003,23(12):2677-2688.
[12]Xiong S M,Zuo X F,Zhu Y Y. Determination of cellulose,hemi-cellulose and ligin in rice hull[J]. Cereal and Feed Industry,2005(8):40-41.
[13]Li J M,Jin Z X,Zhu X Y. Comparison of the total tannin in different organs ofCalycanthuschinensis[J]. Guihaia,2007,27(6):944-947.
[14]金则新,李钧敏,朱小燕. 不同生境夏腊梅营养器官总酚含量分析[J]. 安徽农业大学学报,2006,33(4):454-457.
[15]王联珠,李晓庆,顾晓慧,等. 干海参外源性总糖的测定方法[J]. 食品科学,2003,34(14):293-297.
[16]Hua Y,Jian L,He W M,et al.Cuscutaaustralisrestrains three exotic invasive plants and benefits native species.[J]. Biological Invasions,2011,13(3):747-756.
[17]Feng Y L,Lei Y B,Wang R F,et al. Evolutionary tradeoffs for nitrogen allocation to photosynthesis versus cell walls in an invasive plant[J]. Proceedings of the National Academy of Sciences,2009,106(6):1853-1856.
[18]丁 丹,陈 超. 红毛草(Rhynchelytrumrepens)入侵特性、地理分布和风险评估[J]. 杂草学报,2016,34(2):29-33.
[19]钟军弟,周宏彬,刘锴栋,等. 3种菊科入侵植物白花鬼针草、胜红蓟和假臭草的种子生物学特性比较研究[J]. 杂草学报,2016,34(2):7-11.
[20]田旭飞,曲 波. DNA条形码技术在入侵植物刺萼龙葵检验检疫中的应用[J]. 杂草学报,2017,35(1):30-35.
[21]张 静,闫 明,李钧敏. 不同程度南方菟丝子寄生对入侵植物三叶鬼针草生长的影响[J]. 生态学报,2012,32(10):3136-3143.
[22]郭艳玲,张鹏英,郭默等. 次生代谢产物与植物抗病防御反应[J]. 植物生理学报,2012,48(5):429-434.
[23]郭素民. 入侵植物喜旱莲子草对天敌的响应及机制研究[D]. 临汾:山西师范大学,2014.
猜你喜欢
——中国水陆画艺术精品展
中华书画家(2022年5期)2022-07-20
散文选刊·下半月(2022年6期)2022-06-23
文萃报·周二版(2021年48期)2021-12-16
科学与财富(2021年36期)2021-05-10
天天爱科学(2020年8期)2020-09-10
人与自然(2019年5期)2019-07-24
环球时报(2019-07-08)2019-07-08
环球时报(2017-05-31)2017-05-31
台声(2016年16期)2016-11-25
现代企业(2015年6期)2015-02-28
