
高效氯氰菊酯对甘蓝夜蛾幼虫中肠消化酶活性的影响
2018-06-29 04:58张仙红张建刚
江苏农业科学 2018年11期
张 丽, 张仙红, 张建刚
(山西农业大学,山西太谷 030801)
甘蓝夜蛾[Mamestrabrassicae(L.)]别称甘蓝夜盗蛾、地蚕、菜夜蛾等,属鳞翅目夜蛾科,是农业生产中重要害虫之一,此虫食性杂、寄主范围广,在国内各蔬菜产区均有发生和危害。甘蓝夜蛾具有突发成灾、危害严重、抗药性强、防治困难等特点[1],严重危害蔬菜作物。在我国多地曾出现过大面积的大白菜受甘蓝夜蛾危害,几天内造成绝收的现象[2],给农业生产造成巨大损失。
昆虫消化酶是由昆虫中肠上皮细胞形成和分泌的,其活性直接影响昆虫对食物中营养物质的消化、吸收。对消化系统组织结构的破坏或对消化酶系的干扰必将导致昆虫生长发育受阻乃至死亡。昆虫消化酶活性的高低直接反映了昆虫对营养物质吸收利用的能力。淀粉酶和蛋白酶是昆虫体内能量合成及消化必需重要的2种酶[3-4]。
在杀虫剂市场,拟除虫菊酯类杀虫剂使用量排在第2位,其在我国的施用面积占杀虫剂总施用面积的1/3以上。高效氯氰菊酯是拟除虫菊酯类农药的代表,它的生物活性较高,具有触杀和胃毒作用,由于其具有高效、广谱、低毒和低残留等特点,成为一种应用广泛的新型农药。拟除虫菊酯类农药对光和热较稳定[5],半衰期较长,因此在自然条件下很难快速降解,如任由其在环境中滞留,会造成严重的农药残留问题。若使用不当将成为环境污染物,因此拟除虫菊酯类农药在环境中的残留备受人们关注。因此,以甘蓝夜蛾不同龄期的幼虫为试材,用不同浓度的高效氯氰菊酯进行饲喂,测定饲喂 3~24 h 后幼虫中肠消化酶比活力,进行高效氯氰菊酯杀虫剂对甘蓝夜蛾幼虫消化酶影响的研究,并对农药的使用剂量提供指导性的建议,旨在为高效氯氰菊酯的合理使用、降低残留提供理论依据。
1 材料与方法
1.1 供试虫源
甘蓝夜蛾卵块,采自山西农业大学未施农药的园艺菜地,在室内用园艺菜地的甘蓝饲养至4、5、6龄期备用。
1.2 供试试剂
4.5%高效氯氰菊酯乳油,由北京中保绿农科技集团有限公司提供。
1.3 试虫处理
试验取4.5%高效氯氰菊酯乳油稀释1 500、2 000、2 500 倍3个浓度,并以清水为空白对照组。将菜叶切成 10~15 cm 2小块于不同浓度的药液中浸渍30 s,取出并别上曲别针悬挂晾干,置于直径为15 cm培养皿中。取蜕皮后 1 d 4、5、6龄的幼虫饥饿6 h后,移入装有不同浓度农药的培养皿中。观察记录处理3、6、9、12、24 h的中毒情况。每组6头幼虫,重复3次。对照组死亡率小于10%的试验为有效试验。
1.4 酶液的制备
取样后,立即在冰盘上迅速解剖幼虫,然后加入冰冷(0.0~0.5 ℃)的0.15 mol/L生理盐水冲洗,截取试虫中肠,并于 -70 ℃ 冷冻冰箱冰冻保存。测试前,将中肠组织用滤纸吸掉表面水分后,按鲜质量1 g ∶20 mL加0.2 mol/L pH值为6.9的磷酸盐缓冲液,在冰浴中匀浆,匀浆液在4 ℃、4 000 r/min 下离心20 min,取上清液作为酶原。
1.5 蛋白酶活性测定
参照北京大学生物系生物化学教研室的方法[6]对蛋白酶活性进行测定。取20 μL酶液加入50 μL 0.5%酪蛋白,混匀后于37 ℃水浴15 min,然后加入50 μL 10%三氯乙酸,混匀后在4 ℃、10 000 r/min下离心15 min,取50 μL上清液移入离心管中,加入250 μL 0.55 mol/L碳酸钠溶液,25 μL福林-酚试剂,在37 ℃条件下水浴15 min。最后在波长为 680 nm 处测定吸光度D680 nm。
1.6 淀粉酶活性的测定
参照李志刚等的方法[7]对淀粉酶的活性进行测定。以 50 μL 2%淀粉为底物,加入200 μL 0.2 mol/L磷酸缓冲液(pH值=5.8),20 μL酶液与之反应。室温下反应10 min后,将其置于37 ℃条件下水浴60 min,加250 μL 3,5-二硝基水杨酸终止反应,然后沸水浴5 min,于波长550 nm处测定吸光度D550 nm。无酶反应体系用20 μL 0.2 mol/L磷酸缓冲液(pH值=5.8)代替酶液,作为对照。
1.9 数据处理
用SPSS 15.0软件进行方差分析,并采用Duncan’s新复极差法进行平均数的多重比较。
2 结果与分析
2.1 高效氯氰菊酯对甘蓝夜蛾幼虫蛋白酶活性的影响
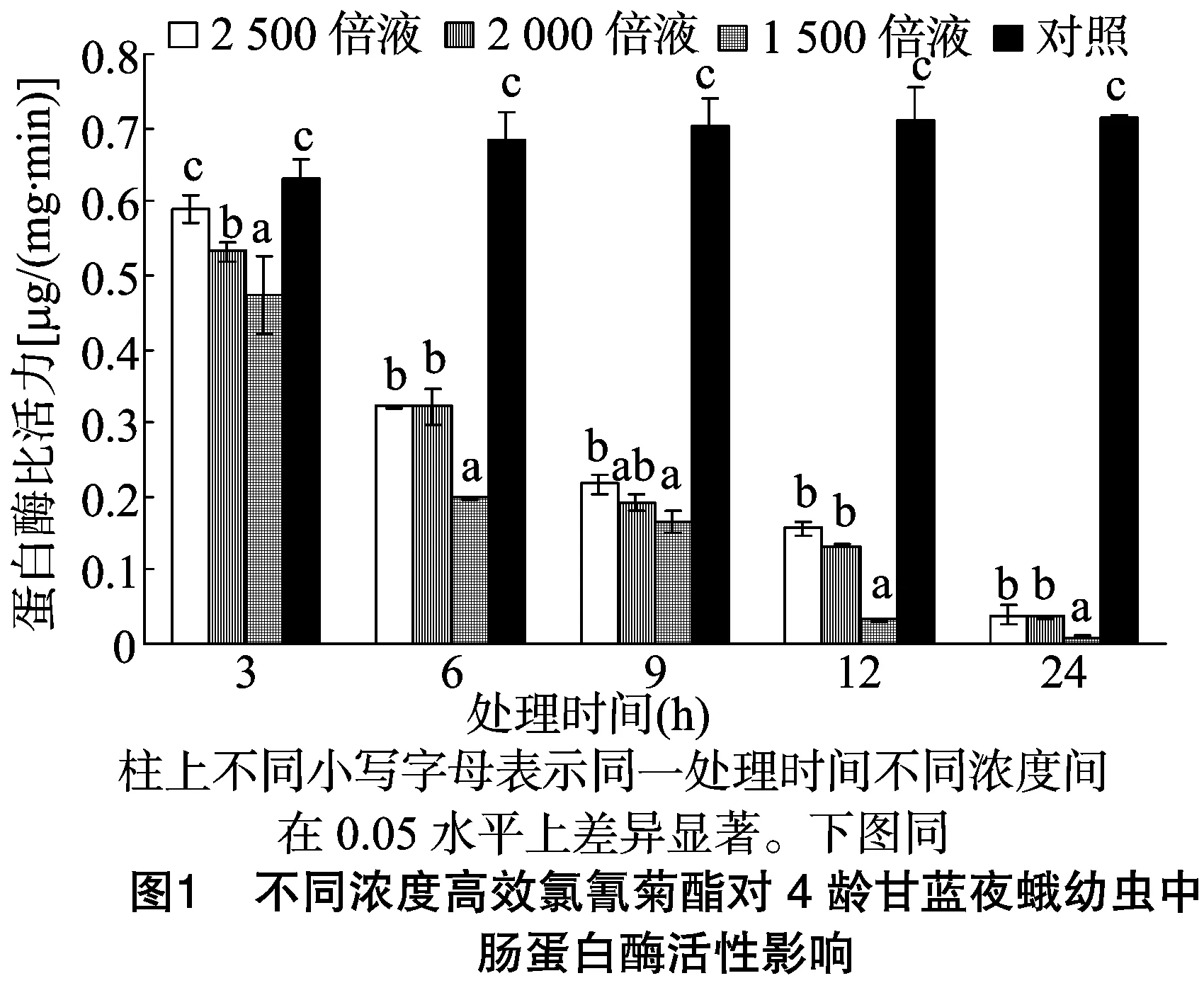
由图1可以看出,甘蓝夜蛾4龄幼虫取食不同浓度高效氯氰菊酯处理的甘蓝叶片后,其体内中肠蛋白酶比活力随着时间的推移逐渐降低,且高效氯氰菊酯浓度越高,中肠蛋白酶比整体越低。当甘蓝夜蛾4龄幼虫取食稀释2 500、2 000、1 500 倍的4.5%高效氯氰菊酯处理的甘蓝叶片3 h后,其蛋白酶比活力分别为对照的0.93、0.84、0.75倍;各处理经邓肯氏新复极差(Duncan’s multiple range test,简称DMRT)法统计分析,2 500倍浓度处理与对照差异不显著,另外2个浓度处理与对照差异显著;当取食6 h后,各处理蛋白酶比活力分别为对照的0.47、0.47、0.29倍;当取食 9 h 后,各处理蛋白酶比活力分别为对照的0.31、0.27、0.23倍;当取食12 h后,各处理蛋白酶比活力分别为对照的0.22、0.19、0.05倍;当取食24 h后,各处理蛋白酶比活力分别为对照的0.05、0.05、0.01倍;取食6、9、12、24 h 后各浓度处理的蛋白酶比活力与对照均表现显著差异。不同浓度的高效氯氰菊酯处理甘蓝夜蛾幼虫后,其体内蛋白酶比活力整体随着高效氯氰菊酯浓度的增加整体呈下降趋势,其中处理24 h后下降得更为明显,且显著低于对照,可见随着高效氯氰菊酯浓度的提高,对甘蓝夜蛾幼虫蛋白酶比活力的影响程度显著增加。
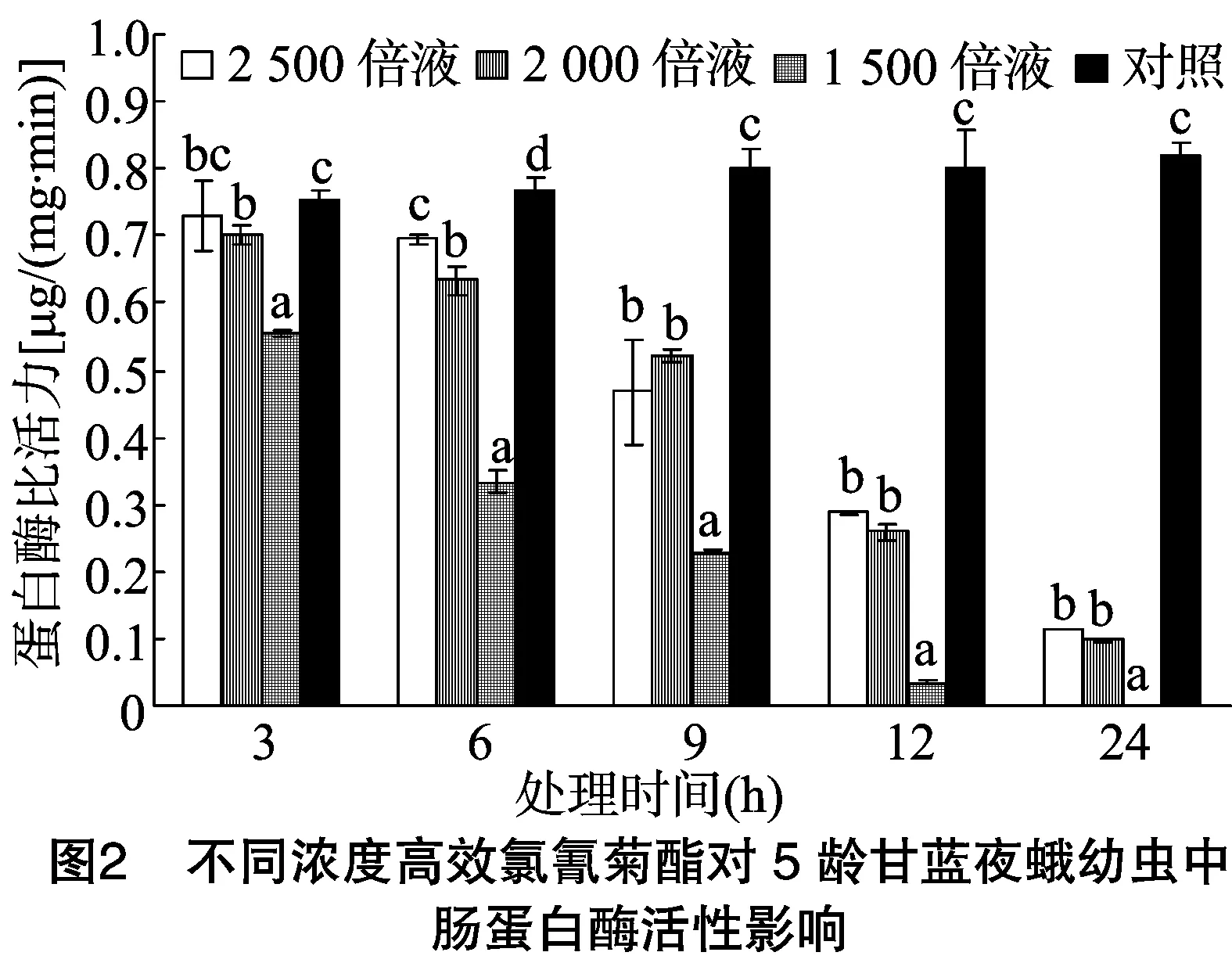
由图2可以看出,甘蓝夜蛾5龄幼虫取食不同浓度高效氯氰菊酯处理的甘蓝叶片后,其体内中肠蛋白酶比活力随着时间的推移逐渐降低,且高效氯氰菊酯浓度越高,中肠蛋白酶比活力(除9 h外)越低。当甘蓝夜蛾5龄幼虫取食稀释 2 500、2 000、1 500倍的4.5%高效氯氰菊酯处理的甘蓝叶片 3 h 后,其蛋白酶比活力分别为对照的0.97、0.93、0.73倍,其中稀释2 500倍浓度处理与对照差异不显著,另外2个浓度处理均与对照差异显著。当取食6 h后,各处理蛋白酶比活力分别为对照的0.91、0.82、0.43倍;当取食9 h后,各处理蛋白酶比活力分别为对照的0.58、0.65、0.28倍;当取食12 h后,各处理蛋白酶比活力分别为对照的0.36、0.32、0.04倍;当取食24 h后,各处理蛋白酶比活力分别为对照的0.14、0.12、0.000 7倍;各处理经DMRT法统计分析,取食6、9、12、24 h后各浓度处理的蛋白酶比活力与对照均表现显著差异。不同浓度的高效氯氰菊酯处理甘蓝夜蛾幼虫后,其体内蛋白酶比活力同4龄呈相似的变化趋势,即随着高效氯氰菊酯浓度的增加,其蛋白酶比活力整体呈下降趋势,其中处理24 h后下降得更为明显,且各处理显著低于对照,可见随着高效氯氰菊酯浓度的提高,对甘蓝夜蛾幼虫蛋白酶比活力的影响程度整体显著增加,但整个下降趋势没有4龄幼虫时明显。
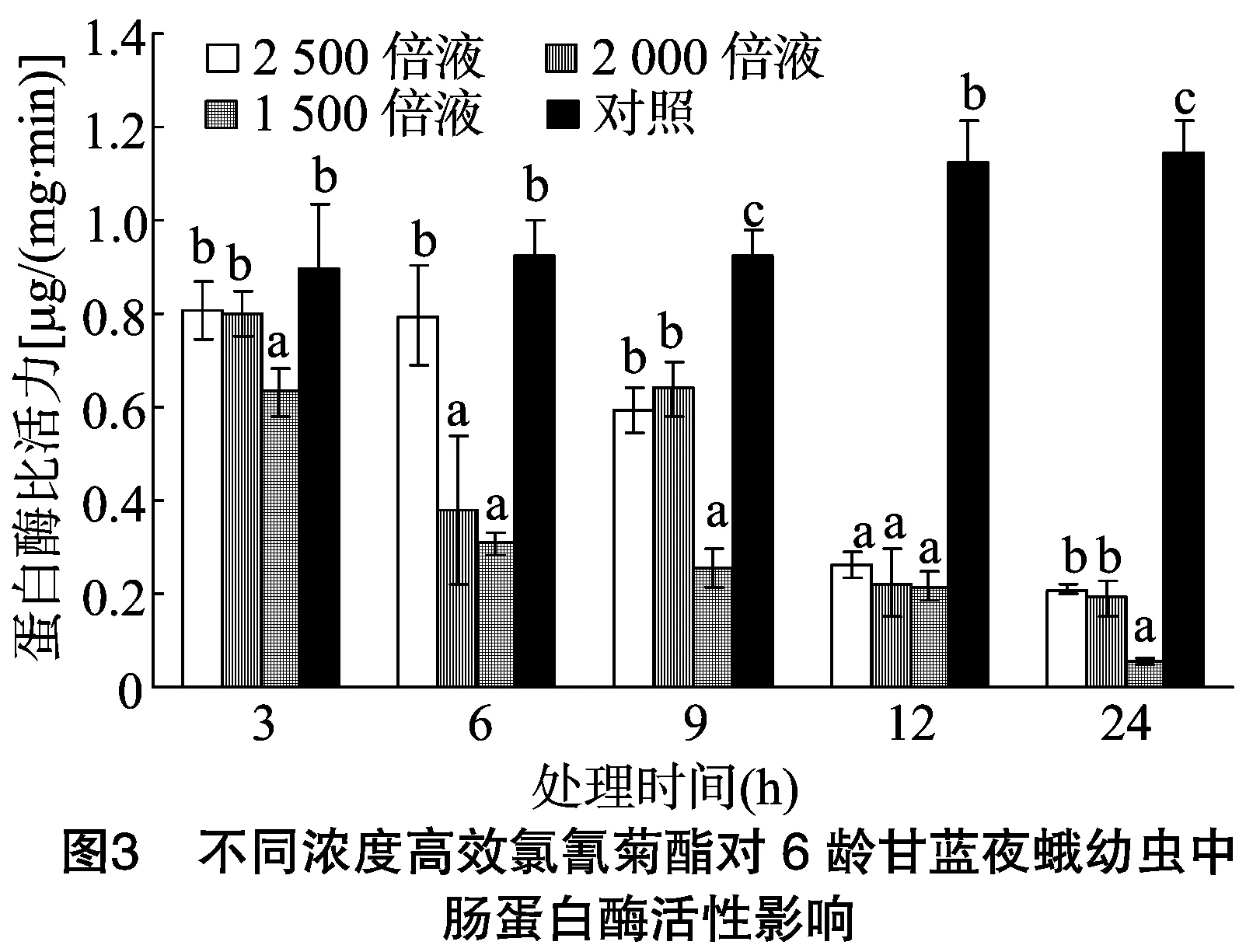
由图3可以看出,甘蓝夜蛾6龄幼虫取食不同浓度高效氯氰菊酯处理的甘蓝叶片后,其体内中肠蛋白酶比活力随着时间的推移逐渐降低,且高效氯氰菊酯浓度越高,中肠蛋白酶比活力越低。当甘蓝夜蛾6龄幼虫取食稀释2 500、2 000、1 500 倍的4.5%高效氯氰菊酯处理的甘蓝叶片3 h后,其蛋白酶比活力分别为对照的0.90、0.89、0.70倍,其中稀释 2 500倍、2 000倍浓度处理与对照差异不显著,稀释 1 500 倍浓度处理与对照差异显著。当取食6 h后,各处理蛋白酶比活力分别为对照的0.86、0.41、0.33倍,稀释2 500倍浓度处理与对照差异不显著,稀释2 000倍、1 500倍浓度处理与对照差异显著;当取食9 h后,各处理蛋白酶比活力分别为对照的0.64、0.69、0.28倍;当取食12 h后,各处理蛋白酶比活力分别为对照的0.23、0.20、0.19倍;当取食24 h后,各处理蛋白酶比活力分别为对照的0.18、0.17、0.05倍。取食9、12、24 h 后各浓度处理的蛋白酶比活力与对照均表现显著差异。不同浓度的高效氯氰菊酯处理甘蓝夜蛾幼虫后,其体内蛋白酶比活力同4、5龄有相似的变化趋势,即随着高效氯氰菊酯浓度的增加蛋白酶比活力整体呈下降趋势,其中处理24 h后下降得更为明显,且各浓度处理显著低于对照,可见随着高效氯氰菊酯浓度的提高,对甘蓝夜蛾幼虫蛋白酶比活力影响程度整体显著增加,但整个下降趋势没有4、5龄幼虫时明显。
2.2 高效氯氰菊酯对甘蓝夜蛾幼虫淀粉酶活性的影响
由图4可以看出,甘蓝夜蛾4龄幼虫取食不同浓度高效氯氰菊酯处理的甘蓝叶片后,其体内中肠淀粉酶比活力随着时间的推移逐渐降低,且高效氯氰菊酯浓度越高,中肠淀粉酶活力越低。当甘蓝夜蛾4龄幼虫取食稀释2 500、2 000、1 500倍的4.5%高效氯氰菊酯处理的甘蓝叶片3 h后,各处理淀粉酶比活力分别为对照的0.73、0.68、0.46倍;当取食6h后,各处理淀粉酶比活力分别为对照的0.66、0.62、0.39倍;当取食9 h后,各处理淀粉酶比活力分别为对照的0.48、0.45、0.29倍;当取食12 h后,各处理淀粉酶比活力分别为对照的0.37、0.34、0.22倍;当取食24 h后,各处理淀粉酶比活力分别为对照的0.17、0.16、0.04倍。取食3、6、9、12、24 h后各浓度处理的淀粉酶比活力与对照均表现为显著差异。不同浓度高效氯氰菊酯处理甘蓝夜蛾幼虫后,其体内淀粉酶比活力随着高效氯氰菊酯的浓度的增加明显下降,可见随着高效氯氰菊酯浓度的提高,对甘蓝夜蛾幼虫淀粉酶活性影响程度明显增加。
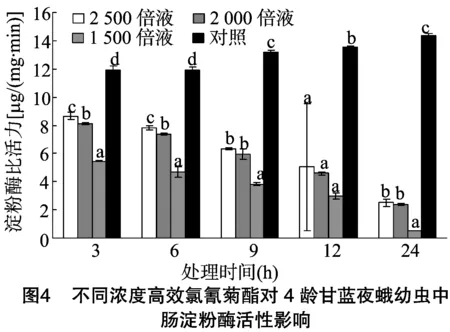
与同龄期蛋白酶比活力相比,随着高效氯氰菊酯浓度的增加,对甘蓝夜蛾幼虫中肠淀粉酶的影响更加明显,在取食 3 h 时各浓度处理即出现显著差异。
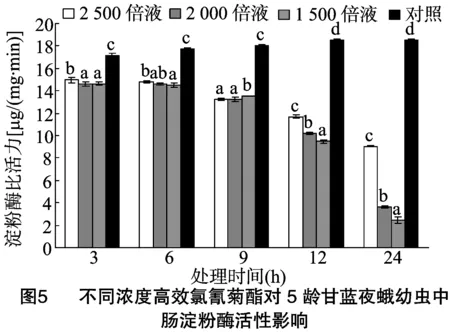
由图5可以看出,甘蓝夜蛾5龄幼虫取食不同浓度高效氯氰菊酯处理的甘蓝叶片后,其体内中肠淀粉酶活力随着时间的推移逐渐降低,且高效氯氰菊酯浓度越高,中肠淀粉酶比活力整体越低。当甘蓝夜蛾5龄幼虫取食稀释2 500、2 000、1 500倍的4.5%高效氯氰菊酯处理的甘蓝叶片3 h后,其淀粉酶比活力分别为对照的0.87、0.85、0.85倍;当取食6 h后,各处理淀粉酶比活力分别为对照的0.83、0.82、0.82倍;当取食9 h后,各处理淀粉酶比活力分别为对照的0.73、0.73、0.75倍;当取食12 h后,各处理淀粉酶比活力分别为对照的0.63、0.55、0.51倍;当取食24 h后,各处理淀粉酶比活力分别为对照的0.49、0.20、0.13倍。取食3、6、9、12、24 h后各浓度处理的淀粉酶比活力与对照均表现显著差异。不同浓度的高效氯氰菊酯处理甘蓝夜蛾幼虫后,其体内淀粉酶比活力同4龄有相似的变化趋势,即随着高效氯氰菊酯浓度的增加淀粉酶比活力整体呈下降趋势,可见随着高效氯氰菊酯浓度的提高,对甘蓝夜蛾幼虫淀粉酶活力影响程度整体明显增加。但与4龄时相比,没有其下降趋势明显。
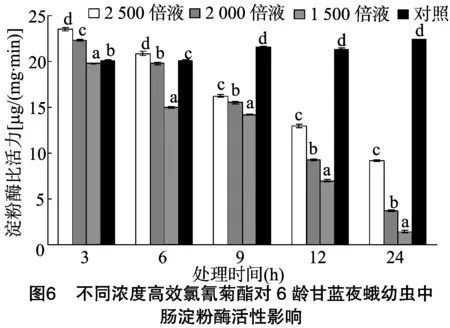
由图6可以看出,甘蓝夜蛾6龄幼虫取食不同浓度高效氯氰菊酯处理的甘蓝叶片后,其体内中肠淀粉酶比活力随着时间的推移逐渐降低,且高效氯氰菊酯浓度越高,中肠淀粉酶比活力越低。当甘蓝夜蛾6龄幼虫取食稀释2 500、2 000、1 500 倍的4.5%高效氯氰菊酯处理的甘蓝叶片3 h后,其淀粉酶比活力分别为对照的1.17、1.11、0.98倍,此时,稀释 2 500 倍、2 000倍浓度处理后淀粉酶比活力与对照相比偏高,且差异显著。当取食6 h后,其淀粉酶比活力分别为对照的1.04、0.98、0.74倍,稀释2 500倍浓度处理后淀粉酶比活力依然与对照相比偏高,且与对照差异显著,而稀释2 000倍、1 500 倍浓度处理后的淀粉酶比活力比对照低,且与对照差异显著;当药物随着时间不断渗透,取食9 h后,各处理淀粉酶比活力分别为对照的0.75、0.72、0.66倍;当取食12 h后,各处理淀粉酶比活力分别为对照的0.61、0.43、0.33倍;当取食24 h后,各处理淀粉酶比活力分别为对照的0.41、0.17、0.06倍。取食9、12、24 h 后各浓度处理的淀粉酶比活力与对照均表现显著差异。高效氯氰菊酯处理甘蓝夜蛾6龄幼虫,除稀释1 500倍浓度处理低于对照外,起始阶段(3 h)淀粉酶比活力与对照相比有所增强,但随着药物的不断渗透淀粉酶比活力逐渐降低,当处理 9 h 后,淀粉酶比活力要低于对照,并且下降幅度随着高效氯氰菊酯浓度的增加而增加。
3 结论与讨论
研究结果表明,甘蓝夜蛾幼虫经高效氯氰菊酯处理后,淀粉酶和蛋白酶2种消化酶比活力有不同的影响,除了6龄幼虫在3、6 h时的淀粉酶比活力有高于对照的现象外,其他消化酶比活力均低于对照,其影响程度随着剂量的增加整体呈增大趋势,并且随着龄期的增大影响程度在逐渐弱化。这样就势必影响昆虫对食物的消化和对营养物质的吸收,减少了昆虫的取食量,从而干扰昆虫正常的生长和发育,抗逆性降低。所以在用药时龄期越小越容易控制。对甘蓝夜蛾施用高效氯氰菊酯时,应尽量在4龄前使用,这样在低剂量情况下不但可以完全控制种群数量,还可以减少药剂残留。
淀粉酶是昆虫消化系统中重要的消化酶,该酶可将昆虫体内的淀粉、糖原等水解成为可溶性麦芽糖、葡萄糖,参与体内的能量代谢[8]。抑制昆虫淀粉酶的活性,能够降低昆虫体内糖类的同化作用,从而抑制昆虫的生长发育[9];抑制昆虫体内淀粉酶的活性可以降低其体内综合生理功能的效率,同样影响其自身的生长发育[10-11]。本研究结果表明,高效氯氰菊酯处理甘蓝夜蛾6龄幼虫时,当处理3、6 h后,稀释2 500倍浓度处理的甘蓝夜蛾幼虫中肠淀粉酶比活力分别为对照的1.17、1.04倍,与对照相比偏高,且差异显著。但随着药物的不断渗透淀粉酶比活力逐渐降低;当处理9 h后,淀粉酶比活力低于对照。经分析可能是因为药物进入肠腔后,刺激肠壁细胞超量分泌消化酶来与药物结合或降解它,以缓和或消除其毒害作用,这样就使虫体消耗了大量用于合成消化酶的氨基酸和能量,导致体能衰竭。但随着药物不断渗透,甘蓝夜蛾幼虫取食量减少、酶底物的量不足,造成酶比活力逐渐下降。与莫建初等报道的结果[12-13]相似。
本试验中高效氯氰菊酯对甘蓝夜蛾幼虫中肠消化酶活性有一定的抑制作用,且对蛋白酶活性的抑制作用比其他消化酶更明显,研究结果与骆颖等所研究的苦瓜叶乙酸乙酯萃取物对斜纹夜蛾消化酶活性的影响结论[14]类似。蛋白酶比活力下降较多可能是因为试虫本身处于饥饿状态,虽然人们把原因归结为营养不足,但不能排除农药对酶结构的影响。近年来大量研究表明,抑制肠道蛋白酶的活性可降低许多鳞翅目昆虫肠道综合生理功能的效率[15]。昆虫取食了含蛋白酶抑制剂的食料后,由于消化功能受阻,表现为生长发育不良[16]。对昆虫中肠消化酶活性的抑制,使食物的营养转换不足,影响昆虫的生活力。
参考文献:
[1]吴 晓,付晓伟,赵新成,等. 甘蓝夜蛾MamestrabrassicaeL. 飞行能力研究[J]. 应用昆虫学报,2016,53(3):595-603.
[2]张振海,杨振平,孟新房,等. 2.4%多杀霉素·高氯氟水乳剂防治甘蓝甜菜夜蛾田间药效试验[J]. 陕西农业科学,2011,57(2):45-47.
[3]Tatun N,Singtripop T,Tungjitwitayakul J,et al. Regulation of soluble and membrane -bound trehalaseactivity and gene expression of the enzyme in the larval midgut of the bamboo borerOmphisafuscidentalis[J]. Insect Biochemistry and Molecular Biology,2008,38(8):788-795.
[4]Wegener G,Tschiedel V,Schlöder P,et al. The toxic and lethal effects of the trehalase inhibitor trehazolin in locusts are caused by hypoglycaemia[J]. Journal of experimental Biology,2003,206(7):1233-1240.
[5]郭勇飞,尹明明,陈福良. 乳化剂对4.5%高效氯氰菊酯水乳剂物理稳定性的影响[J]. 农药学学报,2011,13(1):71-78.
[6]北京大学生物系生物化学教研室. 生物化学实验指导[M]. 北京:高等教育出版社,1987.
[7]李志刚,韩诗畴,郭明昉,等. 取食不同食料植物对安婀珍蝶的营养利用及中肠四种酶活力的影响[J]. 昆虫学报,2005,48(5):674-678.
[8]李艺琼,姚羽芯,彭正强,等. 不同生长阶段椰树叶片对椰心叶甲中肠消化酶活性的影响[J]. 江苏农业科学,2017,45(7):94-97.
[9]Ishimoto M,Yamada T,Kaga A. Insecticidal activity of anα-amylase inhibitor-like protein resembling a putative precursor ofα-amylase inhibitor in the common beanPhaseolusvulgarisL.[J]. Biochimica et Biophysica Acta,1999,1432(1):104-112.
[10]Slansky F J,Scriber J M. Food consumption and utilization[M]//Comprehensive insect physiology,biochemistry and pharmacology. Oxford:Pergamon Press,1985:87.
[11]Timmins W A,Reynolds S E. Azadirachtin inhibits secretion of trypsin in midgut ofManducasextacaterpillars:reduced growth due to impaired protein digestion[J]. Entomologia Experimentalis et Applicata,1992,63(1):47-54.
[12]莫建初,张 军,王问学. 芫花粗提物对鳞翅目幼虫消化酶和解毒酶的影响[J]. 中南林学院学报,2001,21(1):5-9.
[13]谭世强,张爱华,许永华,等. 人参总皂苷对黏虫体内蛋白质含量及消化酶活性的影响[J]. 中国中药杂志,2013,38(11):1692-1696.
[14]骆 颖,张茂新,凌 冰. 苦瓜叶乙酸乙酯萃取物对斜纹夜蛾消化酶活性的影响[J]. 环境昆虫学报,2012,34(3):289-294.
[15]Terra W R,Ferreira C. Insect digestive enzymes:properties,compartmentalization and function[J]. Comparative Biochemistry and Physiology Part B:Comparative Biochemistry,1994,109(1):1-62.
[16]王琛柱. 害虫防治中植物蛋白酶抑制剂的研究[J]. 世界农业,1992(12):28-29.
猜你喜欢
当代水产(2021年9期)2021-12-02
当代水产(2021年8期)2021-11-04
医学前沿(2021年18期)2021-04-14
世界农药(2019年2期)2019-07-13
食品与机械(2019年1期)2019-03-30
食品与发酵工业(2018年7期)2018-08-10
北方蚕业(2016年2期)2016-10-10
中国茶叶加工(2015年3期)2015-02-27
浙江工业大学学报(2014年5期)2014-08-25
河北大学学报(自然科学版)(2012年3期)2012-03-25
