
基于文献计量分析的疫霉菌抗甲霜灵研究进展
2018-06-26 05:04李坤缘王伟燕高智谋
信阳农林学院学报 2018年2期
刘 冬,李坤缘,王伟燕,李 萍,高智谋*
(1.安徽农业大学,安徽 合肥 230036;2.安庆职业技术学院,安徽 安庆 246003)
甲霜灵属苯酰胺类内吸杀菌剂,具有优良的保护、治疗和铲除作用,国内外广泛用于包括疫霉菌在内的卵菌所致植物病害的防治[1]。由于甲霜灵属于单作用位点内吸杀菌剂,病菌容易对其产生抗性突变[2-3]。目前相关报道显示已有10多种疫霉对甲霜灵产生了抗药性,疫霉菌对甲霜灵抗药性越来越严重和普遍,常导致植物病害防治效果下降甚至失败,使农业生产遭受巨大损失[4-6]。针对疫霉抗甲霜灵研究热点和相关研究论文数量快速增加,近几年对该方面的研究态势进行较为全面和系统的定量分析还未见相关报道,我们对CNKI和Web of science数据库中近十年来发表的学术文献为研究对象,以“疫霉菌”、“甲霜灵”和“抗性”为主题词进行检索,并通过阅读标题和摘要进行有效遴选得相关文献91篇。本文从年发表文章量、研究机构和基金等方面进行分析疫霉菌对甲霜灵的抗性发生现状、抗性遗传及分子机制、环境适合度和抗性治理等问题进行讨论,为疫霉菌抗甲霜灵的深入研究提供参考。
1 数据来源与研究方法
以CNKI和Web of science数据库中2008年1月1日至2017年12月31日发表的文献为数据源,利用文献计量学和知识图谱技术对检索遴选出的91篇文献进行整理分析。
2 结果与分析
2.1 文献数量年份分布、文献来源及疫霉抗甲霜灵的发生情况
在CNKI和Web of science数据库中以“疫霉菌”、“甲霜灵”和“抗性”为主题共检索文献91篇,其中CNKI数据库中45篇,Web of science数据库中46篇(图1)。91篇关于疫霉抗甲霜灵的学术论文,发表在64 种学术刊物,文献分布范围符合布拉德福提出的描述文献分散规律[7]。该领域学术论文分别发表在Phytopathology、Plant disease (缩写为Plant dis)、《农药》、Journal of Mycology and Plant Pathology(缩写为JMP Pathol)、《植物保护学报》和 Journal of Pathology(缩写为J Phytopathol)等刊物上(图2)。文献主要关注点包括疫霉菌抗甲霜灵的遗传及分子机制、疫霉菌对甲霜灵抗性的监测评估及环境适合度和甲霜灵的抗性治理等方面。1980年首次在荷兰、爱尔兰发现了马铃薯晚疫病菌抗甲霜灵菌株,随后在欧洲、北美洲、中美洲、亚洲和中东国家都发现了疫霉菌对甲霜灵产生抗性的现象,而抗药性主要集中在辣椒疫霉、致病疫霉和烟草疫霉等异宗配合菌株上[6,8-9]。辣椒疫霉抗甲霜灵菌株已先后在意大利、智利、美国、西班牙等欧美国家发现,有些田间抗性菌株比率高达81%[1-4,6,10-11]。在我国江西省辣椒疫霉中高抗菌株占27.5%,安徽省中高抗菌株占36.8%,西藏、山西、云南、甘肃等省区也相继发现辣椒疫霉抗甲霜灵菌株的存在[12-14]。2013~2014年检测我国北方内蒙古、黑龙江、吉林、辽宁和河北等5省(自治区)的致病疫霉菌株中抗菌株占绝对优势,高抗菌株数量增加,并成为我国四川、云南、福建、甘肃等马铃薯主要产区产业发展的限制因素之一[15]。不同地区不同疫霉菌甚至是同一疫霉菌对甲霜灵的抗性有所差异,这与病害的发生时间长短、施药量和施药次数有一定的关系[12-15]。
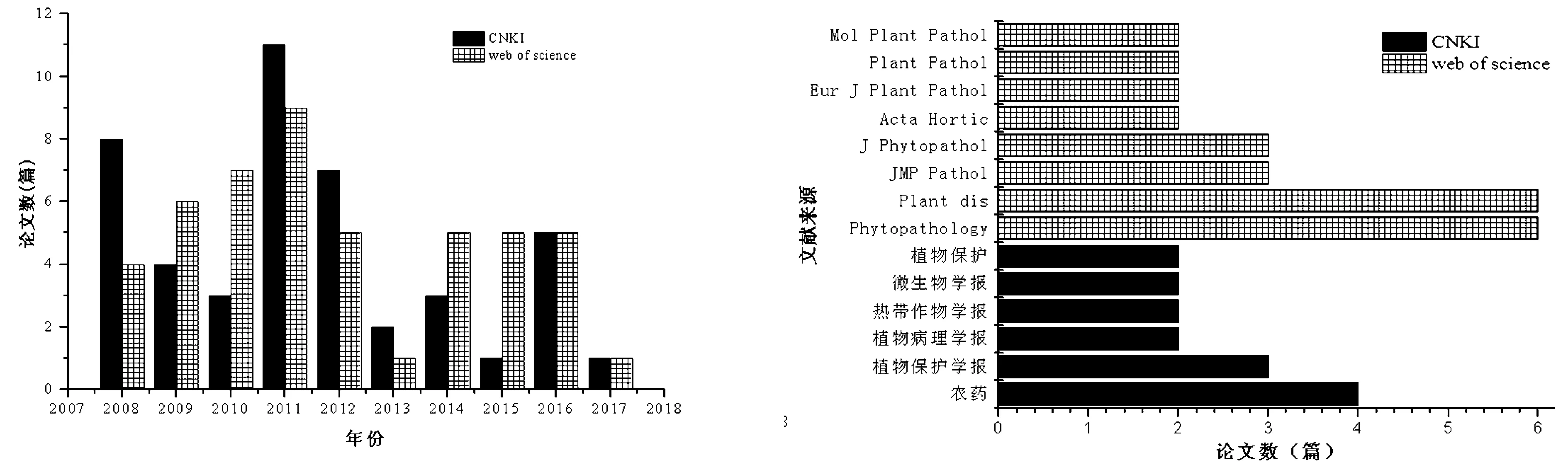
图1文献数量与年份分布情况图2文献来源分布情况
2.2 研究机构与基金分布及疫霉菌抗甲霜灵的分子机制
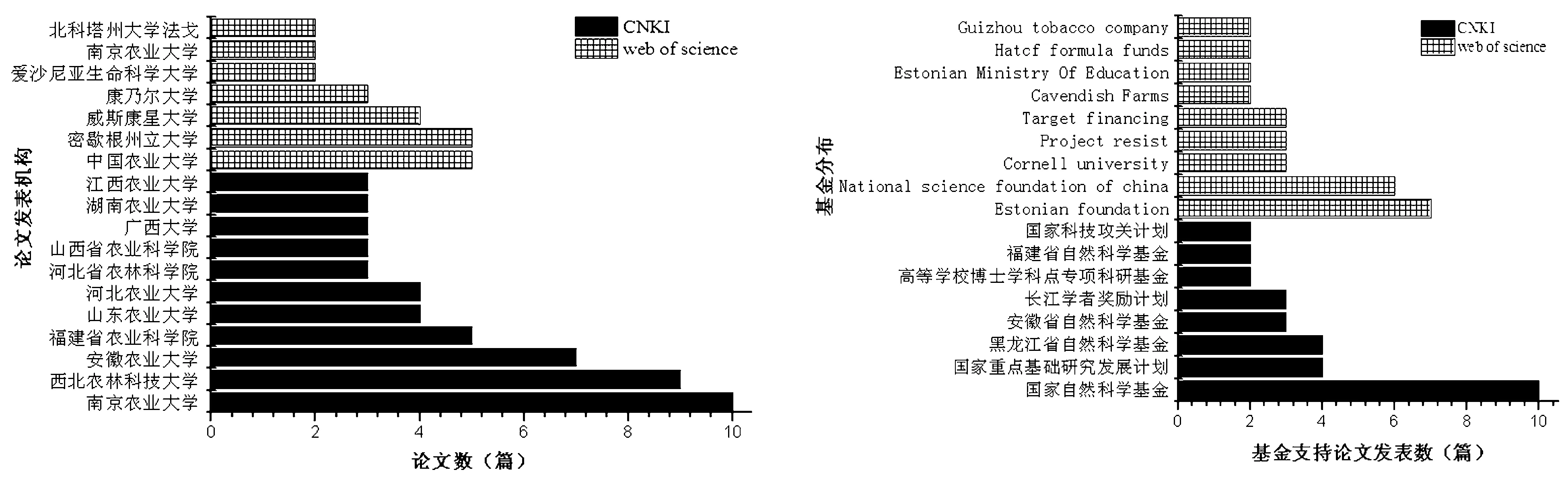
图3研究机构分布情况图4项目基金分布分布情况
91篇关于疫霉菌抗甲霜灵的文献主要来源于农业类院校和科研院所(见图3),如南京农业大学、西北农林科技大学、安徽农业大学、中国农业大学和福建省农业科学院等。在Web of science数据库中,来自密歇根州立大学、威斯康星大学和康乃尔大学的研究人员也在该领域开展了较多的研究。疫霉菌抗甲霜灵的相关研究获得国家自然科学基金、国家重点基础研究发展计划和Estonian foundation等十余类基金的支持(见图4),相关研究成果也同时受到较大的关注和应用。东北农业大学进行的“Pathotypes and metalaxyl sensitivity of Phytophthora sojae and their distribution in Heilongjiang, China 2011-2015”研究得到国家公益性行业( 农业) 科研专项和国家自然科学基金的支持,该成果显示在黑龙江省抗病大豆品种和大豆种子甲霜灵包衣技术相结合是有效控制大豆的根茎和茎腐病的方法。福建农林大学的研究成果“Analysis of RPA190 revealed multiple positively selected mutations associated with metalaxyl resistance in Phytophthora infestans”同样得到国家自然科学基金的支持,文章揭示 RPA190 基因抗甲霜灵的两种进化途径加深了人们对疫霉抗甲霜灵机理的认识。疫霉菌对甲霜灵抗性机理的研究结果表明菌体RNA聚合酶的活性发生了变化是重要的抗性机理[16],类似的结果在马铃薯疫霉中也被证实,马铃薯疫霉RNA聚合酶I大亚基序列的多样性对其抗甲霜灵起到了关键的作用[1]。另外,甲霜灵被菌体的生物降解也是抗性机理之一,敏感菌株RNA聚合酶的活性受到显著抑制,抗性菌株只表现轻微的抑制作用,且抗性菌株对甲霜灵有一定的降解作用[17]。此外,研究人员在疫霉菌对甲霜灵的抗性遗传方面做了较多的工作,他们认为疫霉菌对甲霜灵的抗性由细胞核基因控制,包括不完全显性、完全显性和共显性3种情况,而不完全显性的单一基因对疫霉抗甲霜灵起到关键作用,它参与了辣椒疫霉、致病疫霉和大豆疫霉等病原菌抗甲霜灵的过程[1,4,16-17]。除主要基因效应外,还与几个次要效应基因相关,如ABC转运基因等[18]。王源超等[19]发现了大部分恶疫霉和大雄疫霉对甲霜灵的抗性在游动孢子和自交卵孢子后代均能稳定遗传,但同时发现两种疫霉的部分突变株对甲霜灵的抗性在游动孢子后代遗传不稳定,因此认为这些抗药性既可能是由可稳定遗传的细胞质线粒体基因控制,也可能是由细胞质不稳定遗传的线粒体基因控制。
2.3 疫霉菌对甲霜灵的环境适合度和抗性治理
对甲霜灵产生抗性的菌株能否很快发展形成抗药亚群体,除与药剂选择压力有关,还取决于抗性菌株对环境条件的适合度。适合度主要与抗性菌株的致病力和产孢量等生物学特性有关。郑小波等[20]发现室内绝大多数对甲霜灵有抗性的恶疫霉和大雄疫霉突变株的产孢子囊能力下降。戚仁德等[12]发现辣椒疫霉抗甲霜灵菌株的主要生物学性状与敏感菌株相比无显著差异。而王光飞和马艳[21]的研究表明抗甲霜灵辣椒疫霉菌株环境适合度弱于原始敏感菌株。这可能与疫霉菌本身的特性及地理来源不同有关,造成种间差异性较大。研究抗甲霜灵疫霉菌的环境适合度,对于评估甲霜灵防治疫霉菌的抗药性风险具有重要意义。
目前,疫霉菌对甲霜灵抗药性的治理对策主要是通过降低药剂的选择压力,预防和减缓抗药性的产生。主要包括四个方面:(1)开发应用具有多作用位点的低抗性风险新药剂;(2)筛选与甲霜灵无交互抗性关系或负交互抗性的高效药剂合理复配或混用等,避免病菌与单一杀菌剂的长期接触造成选择压力;(3)研发具有增效作用的杀菌剂混剂,采用不同作用机制的杀菌剂混用;(4)建立病情的预测预报系统,选择在合适的时机用药,提高化学防治的效率,减少用药次数,降低病菌与药剂接触发生突变的几率[6]。
3 结论
本文以CNKI和Web of science数据库中2008-2017年期间收录的关于疫霉菌抗甲霜灵相关研究的文献为研究对象,对疫霉菌抗药性发生现状、分子机制、环境适合度和抗药性治理对策等方面的研究进行分析讨论,对疫霉菌的防治具有一定的参考价值。
猜你喜欢
工业安全与环保(2022年12期)2022-12-30
当代水产(2022年1期)2022-04-26
西北农业学报(2021年7期)2021-08-10
南方农业·上旬(2020年3期)2020-05-06
今日农业(2019年14期)2019-09-18
世界农药(2019年3期)2019-09-10
农药科学与管理(2019年10期)2019-04-20
植物保护(2017年2期)2017-03-29
杂草学报(2015年2期)2016-01-04
中国钱币(2015年6期)2015-11-18
