
元谋干热河谷Savanna草地优势植物的种间关系
2018-06-19 01:05何光熊闫帮国纪中华史亮涛樊博范建成张梦寅
草业学报 2018年6期
何光熊,闫帮国,纪中华,史亮涛*,樊博,范建成,张梦寅
(1.云南省农业科学院热区生态农业研究所,云南 元谋 651300;2.云南省农业科学与农业环境资源研究所,云南 昆明 650205)
种间关系,也称种间亲和性(affinity),通常指群落中多个共存物种之间相互关系的总和,是群落重要的数量和结构特征之一[1]。群落中各个物种尤其是主要优势种之间的关系决定了群落的结构与动态,对群落优势植物种间关系的分析有助于正确认识群落的结构与功能,为自然植被恢复和生物多样性保护提供理论依据[2]。干热河谷是我国西南横断山区一类特殊的气候类型,受梵风效应影响,区域干旱少雨,光热充足,水土流失严重,加之强烈的人为活动,造就了以“稀树灌草丛”为主体的“萨瓦纳(Savanna)”脆弱植被[3-6]。近几十年来,区域可持续发展的需求及脆弱生态系统的典型性,使干热河谷区域成为植被恢复理论及技术研发的重要区域[7-8]。目前,已有大量学者对干热河谷“稀树灌草丛”植被的区系特征及群落结构[9-10]、物种的功能特性[11-12]、优势植物的化学计量特征及生物量分配策略[12-13]、群落与水土保持功能[14-15]等方面做了大量基础性研究工作。但对“稀树灌草丛”植被旱生适应模式的主要结构及运行机制等问题尚缺乏深入认识,制约了相关区域植被恢复模式构建和人工辅助恢复措施制定工作的开展。
对此,本研究基于干热逆境适应模式结构特征的基本问题,提出金沙江干旱河谷“稀树灌草丛”植被优势物种间关系是怎样的问题。随即在元谋干热河谷Savanna草地典型地段设置样方,对区域旱生适应“稀树灌草丛”植被优势物种种间关系进行分析,并对优势植物进行生态种组划分,明确元谋干热河谷Savanna草地适应模式的主要结构,为进一步开展干热逆境环境条件下群落的旱生适应机制研究奠定基础,为干热河谷植被恢复的物种筛选及措施制定提供理论依据。
1 材料与方法
1.1 试验地概况
试验样地位于金沙江干热河谷典型区云南省元谋县境内(25°23′ N,101°35′ E),海拔1210 m,属南亚热带干热季风气候,光热资源充足。年均日照2670.4 h,年平均气温达21 ℃,最高气温42 ℃,最低气温-0.8 ℃,≥10 ℃年积温7996 ℃;年均降水量613.8 mm,主要集中于5-10月,蒸发量为降水量的6.4倍,无霜期350 d。
1.2 研究方法
1.2.1试验设置及优势种确定 元谋干热河谷是金沙江干热河谷的典型地段,1999年始,元谋县政府在境内建立多个水土保持区,区域内多种植被类型得到有效保护,“江边-物茂-新华”一线海拔940~2200 m的区域保存了较为完整的Savanna稀树灌草丛植被。按照系统布点、全面调查及重点精查的原则,在系统了解区域Savanna植被特征的基础上,于2011年7-10月,沿元谋县江边-物茂-新华一线,选择海拔1000~1400 m稀树灌草丛典型地段设置样地7个,其中依据典型性及代表性原则重点对物茂、尹地、丙月3个样地进行清查,并参照样地面积设置1 m×1 m随机样方51个,按《植物群落清查的主要内容、方法和技术规范》[16]对样方进行清查,记录样方内物种出现-不出现、物种数、个体数和物种盖度,优势种随机选取5个植株测定高度,个体数少于5株的物种记录其个体植株高度的全部数据,按物种特征值占样方特征值的比例计算物种相对盖度、相对高度及相对频度,并按公式(1)计算植物种重要值作为优势物种选取的数量指标。本次调查共记录物种74个,选取重要值排序前列的18 个主要植物种(表1)进行种间联结及种间相关分析。
IV=(Rc+Rh+Rf)/3
(1)
式中:IV为某一植物种的重要值;Rc表示相对盖度;Rh表示相对高度;Rf表示相对频度。

其计算公式为:
VR=ST2/σT2
(2)
其中,
(3)
(4)
t=(T1+T2+…+Tn)/N
(5)
式中:VR值为群落内植物种间的总体联结指数;ST为所有样方物种数的方差;σT为所有物种出现频度的方差;S为总物种数;N为总样方数;Pi为物种i出现的频度;Tj为样方j内出现的所研究物种的总数;ni为物种i出现的样方数;t为样方中种的平均数。

表1 优势物种及重要值Table 1 Name and important value of dominant species

(6)
(7)
1.2.4种对间Spearman秩相关分析 以优势种重要值作为Spearman秩相关分析的数量指标,将重要值数据秩化后转变为秩向量,并代入相关系数公式(8)进行计算,得到Spearman秩相关系数rs(i,k)。
(8)
2 结果与分析
2.1 群落总体水平的种间联结


表2 群落总体水平的种间联结Table 2 Inter-specific association for 18 dominant species
2.2 物种的种间联结
采用物种出现-不出现建立优势物种关联矩阵进行χ2检验,结果表明,由18个物种构成的153个种对中,表现为正联结的种对为130对,占总种对数的85.0%;表现为负联结的种对为23对,占总种对数的15.0%。其中,呈现显著关联的种对有5对,占总种对数的3.3%,正联结的种对3对,分别为三芒草(Aa)-独脚金(Sa)、叶下珠(Pu)-蔓草虫豆(Cs)、假杜鹃(Bc)-扁穗白莎草(Dp),负联结的种对2对,为橘草(Cg)-孔颖草(Bp),橘草(Cg)-独穗飘拂草(Fm)(图1)。总体来看,18个优势物种间联结性较弱,具有独立分布的特点。
依据18个优势物种的共同出现百分率绘制Jaccard指数半矩阵图(图2),结果显示,由18个物种构成的153个种对中,JI=0的种对有59对,占总种对数的38.6%;0
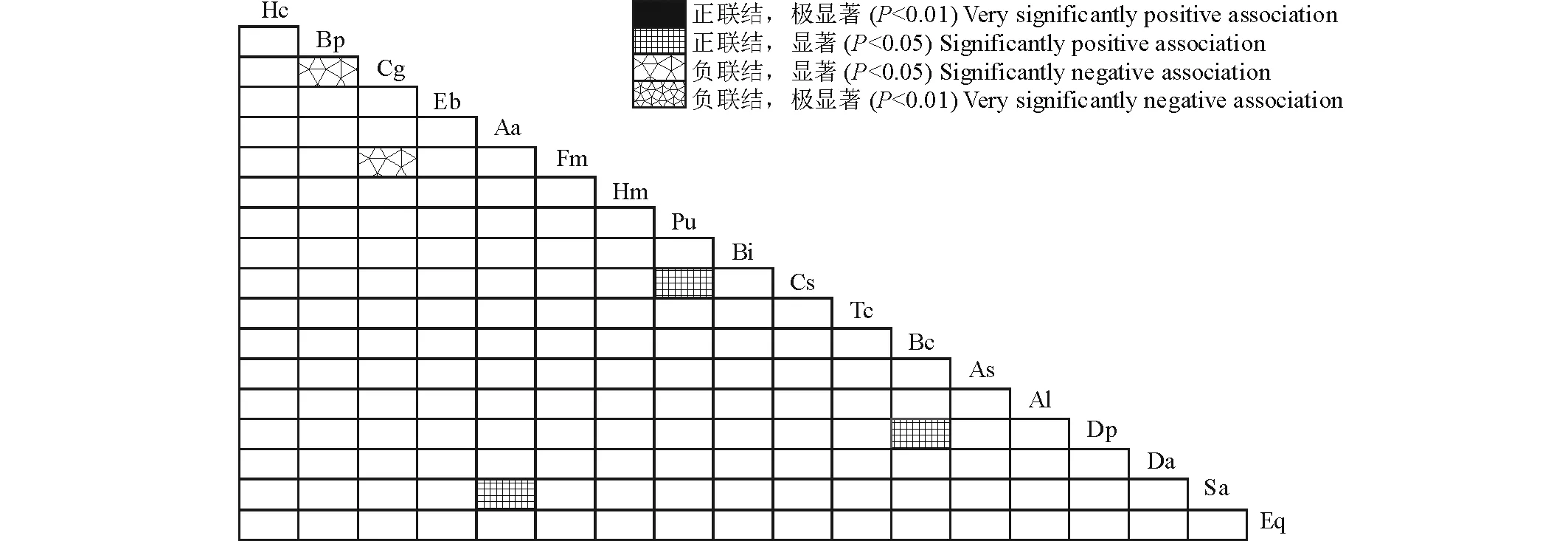
图1 草地18个优势种间χ2半矩阵Fig.1 Semi-matrix of χ2 test of association for 18 dominant species 图中Hc、Bp…、Eq表示的物种与表1物种相同。下同。Species represented by Hc, Bp…, Eq are same to herb species in Table 1. The same below.

图2 草地18个优势种间Jaccard指数半矩阵Fig.2 Semi-matrix of Jaccard’s association index for 18 dominant species
2.3 物种的种间相关
Spearman秩相关分析结果表明(图3),元谋干热河谷18个优势物种间相关关系中,呈正相关的种对有64对,占总对数的41.8%,呈负相关的种对有89对,占总对数的58.2%。其中,呈显著相关的种对31对,呈极显著相关的种对18对,分别占总种对数的20.3%和11.8%;呈显著相关的31个种对中,显著正相关的种对有19对,占总种对数的12.4%,显著负相关的种对有12对,占总种对数的7.8%;呈极显著相关的18个种对中,极显著正相关的种对有12对,分别为孔颖草(Bp)-独穗飘拂草(Fm)、孔颖草(Bp)-叶下珠(Pu)、橘草(Cg)-假杜鹃(Bc)、橘草(Cg)-矛叶荩草(Al)、橘草(Cg)-扁穗白莎草(Dp)、独穗飘拂草(Fm)-叶下珠(Pu)、白羊草(Bi)-独脚金(Sa)、蔓草虫豆(Cs)-双花草(Da)、假杜鹃(Bc)-矛叶荩草(Al)、假杜鹃(Bc)-扁穗白莎草(Dp)、刺芒野古草(As)-四脉金茅(Eq)、矛叶荩草(Al)-扁穗白莎草(Dp),占总种对数的7.8%;极显著负相关的种对有6对,分别为扭黄茅(Hc)-三芒草(Aa)、孔颖草(Bp)-橘草(Cg)、孔颖草(Bp)-拟金茅(Eb)、橘草(Cg)-独穗飘拂草(Fm)、橘草(Cg)-叶下珠(Pu)、拟金茅(Eb)-独穗飘拂草(Fm),占总种对数的3.9%。

图3 草地18个优势种间Spearman相关系数半矩阵Fig.3 Semi-matrix of Spearman’s rank correlation coefficients for 18 dominant species
2.4 生态种组的划分
依据18个优势种的Spearman秩相关关系,以相同种组内物种间具有显著正相关关系,不同种组内物种间具有显著负相关关系为基础绘制网络星座图,将18个优势种划分为4个生态种组(图4)。第1生态种组为:刺芒野古草(As)和四脉金茅(Eq);第2生态种组为:短萼灰叶(Tc)、黑果黄茅(Hm)、矛叶荩草(Al)、假杜鹃(Bc)、扁穗白莎草(Dp)、橘草(Cg)和拟金茅(Eb);第3生态种组为:蔓草虫豆(Cs)、叶下珠(Pu)、双花草(Da)、独穗飘拂草(Fm)、孔颖草(Bp)、三芒草(Aa)和扭黄茅(Hc);第4生态种组为:白羊草(Bi)和独脚金(Sa)。
从图4可以看出,第1生态种组和第4生态种组均只有2个物种组成;第2生态种组与第3生态种组均由7个物种组成。第2生态种组中的物种间形成的正相关关系的种对数(10对)较第3生态种组物种间形成的正相关关系的种对数(8对)多,具有更高的正联结比例。同时,第2生态种组中橘草(Cg)与第3生态种组的叶下珠(Pu)、双花草(Da)、独穗飘拂草(Fm)、孔颖草(Bp)4个物种间形成显著负相关种对,第2生态种组中拟金茅(Eb)与第3生态种组的叶下珠(Pu)、独穗飘拂草(Fm)、孔颖草(Bp)3个物种形成显著负相关种对,占第2生态种组与第3生态种组间负相关种对数的87.5%,成为2个种组种对形成负联结关系的关键种。

图4 禾草地18个优势种间Spearman秩相关系数星座图Fig.4 Constellation diagram of Spearman’s rank correlation coefficient for 18 dominant species
3 讨论
研究表明,干旱等逆境会导致不同物种间对限制性资源的竞争加剧,使不同物种间呈现负关联,而另一方面,有限的资源环境也促使物种间生态位分离而在群落中共存[18],在统计学参数中呈现“共同出现”或“利他”的正关联特征。本研究发现元谋干热河谷Savanna草地植物间主要呈显著的负关联,提示干热河谷草地物种间可能存在较高的资源竞争或生态关系的相互排斥。优势物种组成的种对间联结关系的χ2检验结果及Jaccard指数显示,18个优势物种间联结性较弱,在同一样方中18个优势物种间共同出现的频率较低,也表明干热河谷Savanna草地优势植物具有明显的独立分布特点。提示干热河谷Savanna草地群落容易受到干扰,因而一定的封育措施对于区域植被重建或恢复可能是必要的。
相关研究表明,在进行种间关系分析时,不同程度的χ2检验只能反映出种对间关联性显著与否,而经检验不显著的种对并不意味着它们之间未有关联性,且χ2检验不能区分关联程度的大小,因此,χ2检验只有与Jaccard等相似性系数结合使用才能更好地反应种间关系[14]。本研究发现,在元谋干热河谷18个优势物种组成的153个种对中,仅有6个种对Jaccard指数位于0.3
采用物种出现-不出现建立物种关联矩阵进行χ2检验,并结合物种的Jaccard共同出现百分率指数对群落物种间联结性进行定性分析是种间关联(interspecific association)的重要方法[24],Spearman秩相关系数是一种定量分析方法,能定量反映2个物种同时出现的可能性程度[25]。Spearman秩相关分析结果显示,元谋干热河谷18个优势物种间相关关系中,呈正相关的种对有64对,占总对数的41.8%;呈负相关的种对有89对,占总对数的58.2%。一般认为,种间关联和种间相关并没有必定的内在联系,但Spearman秩相关分析得出的呈显著的种对数要高于χ2检验,在研究种间关系时,将二者结合起来分析,能更完整地反映种间的亲和性及其程度,较确切地解释群落中物种的种间关系[21]。本研究结合种间关联定性分析及种间相关定量分析对元谋干热河谷Savanna草地优势植物的种间关系(种间亲和性)进行分析,均得出该区域优势植物具有明显的独立分布的结论,与采用方差比率法(表2)得出干热河谷草地物种间可能存在较高的资源竞争或生态关系的相互排斥的结论相对应,取得了较为可信的研究结论。
生态种组是对环境的适应能力、对资源的利用能力、对群落所起的功能作用等具有一致性的物种组合[26],因而,同一生态种组内,各物种间具有较强的正联结,而不同种组间联结较为松散,造成这种现象的主要原因是不同物种具有生物学特性的差异,对生境具有不同的生态适应性和相互分离的生态位[27]。依据Spearman秩相关分析的结果,本研究将干热河谷Savanna草地18个优势种划分为4个生态种组。从整体来看,4个生态种组中第2、3种组均有7个物种组成,而第2生态种组的正联结比例较其余3个生态种组高。干旱地区物种丰富度有限,物种间直接竞争弱,群落结构特征主要表现为环境条件对物种生存的限制[28],因此推测第2生态种组的物种可能更适应干热河谷干燥炎热的气候环境。但由于在受水分条件限制的群落中,物种生长发育的随机性大,物种对环境波动的响应模式,必然对物种多样性和群落功能产生重要影响[29]。橘草(Cg)、拟金茅(Eb)、孔颖草(Bp)和扭黄茅(Hc)是金沙江干热河谷区域的优势物种,分析结果显示第2生态种组橘草(Cg)和拟金茅(Eb)与第3生态种组的叶下珠(Pu)、独穗飘拂草(Fm)和孔颖草(Bp)是2个种组种对形成负联结关系的关键种,而扭黄茅(Hc)是与其他物种形成微弱联结关系的广布种,因此,这4个生态种组中的物种是如何响应干热逆境变化,进而对干热河谷Savanna草地物种多样性和群落功能造成影响的,应当成为进一步解读金沙江干热河谷特殊逆境植被修复及功能重建的关键环节。
组成群落的每一个物种,其在决定群落的性质和功能上并不具有完全相同的地位和作用,其中,优势种表现为个体数量多,体积大或生物量高,生活能力强,从而对群落的结构功能及群落环境的形成具有决定性作用[30]。总体来看,干热河谷Savanna草地中,扭黄茅(Hc)明显表现出个体数量多,分布范围广,生活能力强的特点,并与孔颖草(Bp)和橘草(Cg)等形成相对微弱的正相关关系,提示扭黄茅种群的建立可能为区域Savanna草地中其他物种的定植准备了必要的环境条件。因而在区域植被恢复的实践中,利用扭黄茅(Hc)作先锋物种,在局域形成单优种群后再引入孔颖草(Bp)和橘草(Cg)等其他物种,通过种间相互关系建立较为稳定的群落,可作为干热河谷区域植被恢复的重要技术环节。同时,从物种的生理特征来看,4个生态种组中,第1生态种组物种主要出现在砾石成分较多的沙砾土生境;第2生态种组的物种,如拟金茅(Eb)等有相对较低的生长速率和养分含量,趋向于旱生适应,在旱季形成较难分解的枯落物覆盖地表,对区域植被水土保持功能的形成具有较好作用;第3种组的物种,如孔颖草(Bp)等具有较高的补偿生长能力和相对较高的养分含量,因此对区域草地群落营养质量的提高及牧业支撑能力的提升具有重要意义;而第4生态种组的物种,主要出现在群落边际或土壤侵蚀区域容易受干扰因子影响的地段,具有重要的指示作用。通过种间关系的分析明晰这些物种或生态种组的适应策略及相互关系,可为干热河谷区域植被恢复措施的制定、功能性植被重建物种的选择及种组维持措施的制定奠定理论基础。
用数理统计的方法对种间关系进行测度虽可以揭示种间竞争的结果或群落的现状,但不能揭示其过程,难以说明物种种间关系形成的具体原因。但是,利用统计方法测度种间关系仍然是进行生物多样性保护和生态系统恢复的基本前提。本研究主要基于群落清查现状对元谋干热河谷Savanna草地优势植物的种间关系的现状进行分析,这意味着研究结论可能是多个因素共同作用的结果。因此,还需要借助野外控制或半控制试验研究等多种方法对群落进行动态监测,明确种间关系形成的特定原因及过程。
4 结论
元谋干热河谷Savanna草地植物间主要呈现显著的负关联,提示干热河谷“稀疏灌草丛”植被物种间可能存在较高的资源竞争或生态关系的相互排斥。
18个优势物种间联结性较弱,在同一样方中18个优势物种间共同出现的频率较低,具有独立分布的特点。元谋干热河谷18个优势物种组成的153个种对中,呈显著关联的种对有5对,占总种对数的3.3%,仅有6个种对Jaccard指数位于0.3 Spearman秩相关分析结果显示,由18个优势物种组成的153个种对中显著相关的种对31对,呈极显著相关的种对18对,分别占总种对数的20.3%和11.8%,主要呈现正相关关系。 18个优势种划分为4个生态种组。第2生态种组橘草(Cg)和拟金茅(Eb)与第3生态种组的叶下珠(Pu)、独穗飘拂草(Fm)和孔颖草(Bp)是两个种组种对形成负联结关系的关键种。 干热河谷Savanna草地优势物种种间关系较弱,趋向于独立分布,群落的结构依赖于扭黄茅、橘草和拟金茅等优势种或关键种,群落的功能依赖于生态种组物种的特征,整体易受环境或人为因素干扰。相应地,在植被生态管理的实践中,干热河谷Savanna草地群落适宜进行适度封育,植被恢复或功能性植被构建时可选择扭黄茅等先锋物种进行定植,而后引入橘草和拟金茅等不同生态种组物种,通过种间相互作用改善群落质量及功能,恢复过程中重视特殊生态种组的地位和作用。 参考文献References: [1] Zhang J T. Quantitative ecology. Beijing: Science Press, 2011: 100-109. 张金屯. 数量生态学. 北京: 科学出版社, 2011: 100-109. [2] Zhang J T. Quantitative plant ecology. Beijing: China Science and Technology Press, 1995: 107-189. 张金屯. 植被数量生态学方法. 北京: 中国科学技术出版社, 1995: 107-189. [3] Jin Z Z, Ou X K. The phytosociological classification of Braun Blangute’s syntaxa for the dry-Hot valley vegetation in Yunnan-Sichuan region. Acta Botanica Yunnanica, 1998, (3): 29-44. 金振洲, 欧晓昆. 滇川干热河谷植被布朗布朗喀群落分类单位的植物群落学分类. 云南植物研究, 1998, (3): 29-44. [4] Ou X K. The study of flora in Yuanmou dry-hot river valley. Acta Botanica Yunnanica, 1988, 10(1): 11-18. 欧晓昆. 元谋干热河谷植物区系研究. 云南植物研究, 1988, 10(1): 11-18. [5] Jin Z Z, Ou X K, Zhou Y. The general situation of natural vegetation in dry-hot river valley of Yuanmou, Yunnan province. Acta Phytoecologica et Geobotanica Sinica, 1987, 11(4): 308-317. 金振洲, 欧晓昆, 周跃. 云南元谋干热河谷植被概况. 植物生态学与地植物学学报, 1987, 11(4): 308-317. [6] Jin Z Z, Ou X K, Zhou Y. Apreliminary study on the vegetation of dry-hot river valley in Yuanmou, Yunnan. Journal of Southwest China Normal University (Nature Science Edition), 1988, (2): 68-78. 金振洲, 欧晓昆, 周跃. 云南元谋干热河谷植被的初步研究. 西南师范大学学报(自然科学版), 1988, (2): 68-78. [7] Sun T, Bi Y F, Zhao X S,etal. Study on different recover measures of degraded grassland in arid-hot valley along the Jinshajiang river. Grassland and Turf, 2006, (3): 39-44. 孙涛, 毕玉芬, 赵小社, 等. 金沙江干热河谷退化草地植被恢复技术的研究. 草原与草坪, 2006, (3): 39-44. [8] Fang H D, Ji Z H, Sha Y C,etal. Causes of gully formation and its vegetation restoration technology in Yuanmou dry-hot valley. China Forestry Science and Technology, 2006, 20(2): 47-50. 方海东, 纪中华, 沙毓沧, 等. 元谋干热河谷区冲沟形成原因及植被恢复技术. 林业科技开发, 2006, 20(2): 47-50. [9] Zhang J L, Shen R, Shi W,etal. The structure and similarity characteristic of the grassland community in hot-dry valley upper middle and lower of Jinsha River. Ecology and Environmental Sciences, 2010, 19(6): 1272-1277. 张建利, 沈蕊, 施雯, 等. 金沙江流域干热河谷上中下游草地植物群落结构与相似性. 生态环境学报, 2010, 19(6): 1272-1277. [10] Jin Z Z. A phytosociological study on the semi-savanna vegetation in the dry-hot valleys of Yuanjiang River, Yunnan. Guihaia, 1999, (4): 289-302. 金振洲. 云南元江干热河谷半萨王纳植被的植物群落学研究. 广西植物, 1999, (4): 289-302. [11] Yan B G, He G X, Shi L T,etal. Element concentration in leaves and nutrient resorption efficiency on dry-red soil and vertisols in dry and hot valley in Yuanmou, China. Chinese Journal of Applied Ecology, 2016, 27(4): 1039-1045. 闫帮国, 何光熊, 史亮涛, 等. 元谋干热河谷燥红土和变性土上植物叶片的元素含量及其重吸收效率. 应用生态学报, 2016, 27(4): 1039-1045. [12] Yan B G, Liu G C, Fan B,etal. Relationships between plant stoichiometry and biomass in an arid-hot valley, Southwest China. Chinese Journal of Plant Ecology, 2015, 39(8): 807-815. 闫帮国, 刘刚才, 樊博, 等. 干热河谷植物化学计量特征与生物量之间的关系. 植物生态学报, 2015, 39(8): 807-815. [13] Yan B, Ji Z, Fan B,etal. Plants adapted to nutrient limitation allocate less biomass into stems in an arid-hot grassland. New Phytologist, 2016, 211(4): 1232-1240. [14] Li J L, Zhang J T, Yuan J Y. Interspecific relationships of dominant species of sub-alpine scrub community and sub-alpine meadow community in the Guandi Mountains. Acta Agrestia Sinica, 2004, 12(2): 113-119. 李军玲, 张金屯, 袁建英. 关帝山亚高山灌丛群落和草甸群落优势种的种间关系. 草地学报, 2004, 12(2): 113-119. [15] Yang D, Xiong D H, Zhang S,etal. Effects of grass belt length to runoff hydrodynamic properties in gully bed in Jinsha river dry-hot valley region. Journal of Soil and Water Conservation, 2014, 28(6): 77-81. 杨丹, 熊东红, 张素, 等. 金沙江干热河谷冲沟沟床草被对径流水动力特性的影响. 水土保持学报, 2014, 28(6): 77-81. [16] Fang J Y, Wang X P, Shen Z H,etal. Methods and protocols for plant community inventory. Biodiversity Science, 2009, 17(6): 533-548. 方精云, 王襄平, 沈泽昊, 等. 植物群落清查的主要内容、方法和技术规范. 生物多样性, 2009, 17(6): 533-548. [17] Schluter D A. Variance test for detecting species associations, with some example applications. Ecology, 1984, 65(3): 998-1005. [18] Tilman D. Resource competition between plankton algae: an experimental and theoretical approach. Ecology, 1977, 58: 338-348. [19] Guo Z H, Zhuo Z D, Chen J,etal. Interspecific association of trees in mixed evergreen and deciduous broadleaves forest in Lushan mountain. Acta Phytoecologica Sinica, 1997, 21(5): 424-432. 郭志华, 卓正大, 陈洁, 等. 庐山常绿阔叶-落叶阔叶混交林乔木种群种间联结性研究. 植物生态学报, 1997, 21(5): 424-432. [20] Guo X Y, Zhang J T, Gao H W. A study on interspecific association of dominant species inBothriochloaischaemumcommunity in Shanxi. Acta Prataculturae Sinica, 2003, 12(2): 14-19. 郭逍宇, 张金屯, 高洪文. 白羊草群落优势种种间联结性的分析. 草业学报, 2003, 12(2): 14-19. [21] Zhong Y, Zhang J, Yang W Q,etal. Interspecific relationships and ecological species groups of dominant herbage species inEucalyptusgrandisplantation. Acta Prataculturae Sinica, 2010, 19(3): 56-62. 钟宇, 张健, 杨万勤, 等. 巨桉人工林草本层优势种的种间关系及生态种组的划分. 草业学报, 2010, 19(3): 56-62. [22] Peng S L, Zhou H C, Guo S C,etal. Studies on the changes in interspecific association of zonal vegetation in Dinhushan. Chinese Bulletin of Botany, 1999, 41(11): 1239-1244. 彭少麟, 周厚诚, 郭少聪, 等. 鼎湖山地带性植被种间联结变化研究. 植物学报, 1999, 41(11): 1239-1244. [23] Peng L J. Changes in interspecific association of the climatic climax vegetation from 1984 to 2006 in Dinhushan, Guangdong, China. Acta Ecologica Sinica, 2006, 26(11): 3732-3739. 彭李菁. 鼎湖山气候顶极群落种间联结变化. 生态学报, 2006, 26(11): 3732-3739. [24] Wang B S, Peng S L. Studies on the measuring techniques of interspecific association of lower-subtropical evergreen-broadleaved forests Ⅰ. The exploration and the revision on the measuring formulas of interspecific association. Acta Phytoecologica et Geobotanica Sinica, 1985, 9(4): 274-285. 王伯荪, 彭少麟. 南亚热带常绿阔叶林种间联结测定技术研究Ⅰ.种间联结测式的探讨与修正. 植物生态学与地植物学丛刊, 1985, 9(4): 274-285. [25] Zhang Z Y, Zhou L, Shangguan T L,etal. Interspecific relationships of dominant species inCaraganacommunities in semi-arid regions of China. Chinese Journal of Ecology, 2011, 30(9): 1868-1874. 张震云, 周伶, 上官铁梁, 等. 半干旱区锦鸡儿群落优势种种间关系. 生态学杂志, 2011, 30(9): 1868-1874. [26] Song C W, Jiang J, Fan J L,etal. Ecological species groups division based on interspecific association: a case study in the Gurbantunggut desert, Xinjiang, China. Journal of Desert Research, 2012, 32(1): 77-85. 宋春武, 蒋进, 范敬龙, 等. 从植物种间联结性探讨白梭梭种群生态种组分类——以古尔班通古特沙漠为例. 中国沙漠, 2012, 32(1): 77-85. [27] Pang F, Hu Y K, Zhang W,etal. Numerical analysis of inter-specific relationships in Alpine steppe community in Bayanbulak. Acta Ecologica Sinica, 2012, 32(6): 1898-1907. 房飞, 胡玉昆, 张伟, 等. 高寒草原植物群落种间关系的数量分析. 生态学报, 2012, 32(6): 1898-1907. [28] Huston M A, DeAngelis D L. Competition and Coexistence: the effects of resource transport and supply rates. The American Naturalist, 1994, 144(6): 954-977. [29] Shang Z H, Yao A X, Long R J. Analysis on the relationship between the species diversity and the productivity of plant communities in the arid mountainous regions in China. Arid Zone Research, 2005, 22(1): 74-78. 尚占环, 姚爱兴, 龙瑞军. 干旱区山地植物群落物种多样性与生产力关系分析. 干旱区研究, 2005, 22(1): 74-78. [30] Wang B S. Phytocoenology. Beijing: Science Press, 1987: 54-67. 王伯荪. 植物群落学. 北京: 高等教育出版社, 1987: 54-67.
猜你喜欢
河南师范大学学报(自然科学版)(2022年5期)2022-08-08
昆明医科大学学报(2022年2期)2022-03-29
食品安全导刊(2021年20期)2021-08-30
小哥白尼(神奇星球)(2021年3期)2021-07-22
小哥白尼(神奇星球)(2021年4期)2021-07-22
河南科学(2020年3期)2020-06-02
Zoological Research(2019年3期)2019-06-28
蔬菜(2018年5期)2018-05-17
乡村地理(2018年4期)2018-03-23
杂草学报(2015年2期)2016-01-04
