
黑河上游冰沟流域土壤养分与微生物空间异质性研究
2018-06-19 01:06高海宁李彩霞孙小妹陈年来张勇
草业学报 2018年6期
高海宁,李彩霞,孙小妹,陈年来*,张勇*
(1.甘肃农业大学资源与环境学院,甘肃 兰州 730070;2.甘肃省河西走廊特色资源利用省级重点实验室,甘肃 张掖 734000)
土壤有机碳是土壤碳中最具有活性的部分,可代表微生物量的大小,尽管有机碳在土壤全碳中所占的比例很小,但它是土壤有机质中的活性组分,其比例大小能够反映土壤微生物数量和活性,是评价土壤微生物量的重要指标[1]。一般而言,土壤中有机质含量越高,有机碳含量就越高,能够提供给微生物利用的碳源就越多,微生物数量也就越大。
土壤氮素主要来源于土壤有机碎屑、腐殖质等,通过微生物降解而形成植物可利用的有效态氮。土壤中大部分铵态氮和硝态氮很容易被作物直接吸收利用,是速效氮。在土壤中全氮量的消长与土壤有机碳含量的变化一样,主要决定于各地区有机质的积累和分解作用的相对强度[2]。不同立地环境及植被类型是影响微生物活性的因素,对土壤全氮量也具有较大的影响[3]。
土壤微生物是土壤生态系统中最重要的组成部分,通过参与土壤有机物质的循环、运输[4-5],进而主导土壤生态系统养分循环和能量流动,对维持生态系统稳定性和可持续性具有重要意义[6]。氮、磷、钾是植物生长发育必需的营养元素,土壤中N主要是通过固氮作用、硝化作用、反硝化作用及有机氮的矿化作用转化而形成,微生物在这些生化反应中起着主导作用,从而调控植物对氮源的吸收利用[7]。磷和钾主要是通过解磷解钾细菌参与磷和钾循环,从而促进植物对不溶性磷酸钙[8]和钾长石[9]的再吸收利用。在生态系统中,土壤微生物的种类、数量、群落多样性受海拔、土壤养分、植被组成和结构的综合影响[10]。
黑河发源于祁连山,是我国第二大内陆河,对整个流域的生态和经济发展起着关键性作用[11]。目前,由于气候变化和人为活动的干扰及超载放牧,导致天然草地退化,森林面积萎缩,生态系统的脆弱愈加严重,资源和环境发展失衡[12]。因此,许多学者致力于黑河上游祁连山的生态、土壤、植被和水源涵养功能等方面的研究[13-15],而对于不同海拔及其调控因子对于微生物种类的影响研究相对较少,仅见台喜生等[16]对祁连山不同海拔氮磷循环细菌数量变化特征研究,未见海拔梯度的垂直变化与土壤养分和微生物方面的研究工作。鉴于此,本研究选择10个海拔梯度,系统调查了样地植被,采集0~20 cm土壤样品,分析土壤理化性质和土壤可培养微生物随海拔变化而呈现的分布规律,并分析它们与环境因子之间的相关性,以期探讨影响土壤生态系统的主控因子,为揭示林地土壤发育的现状,评价林地土壤肥力提供依据。
1 材料与方法
1.1 样地概况
为了研究黑河上游冰沟流域不同海拔条件下微生物与土壤理化性质的关系,本研究依海拔梯度选取了10个样地。样地以海拔梯度间隔100 m设置,随机设置样方。样地植被类型有3类:草地、灌木、乔木。样地详细概况见表1。
研究地点位于青海省祁连县黑河上游冰沟流域(E 100°11′24″-100°23′55″,N 38°02′18″-38°10′39″,海拔2905~4401 m),年均气温0.5 ℃,降水量290.2~467.8 mm,年均蒸发量1051.7 mm,平均相对湿度60%,年日照时间2600 h[17],成土岩石为石灰岩、砂岩、砾岩,成土母质为残积母质和坡积母质。土壤类型为高山寒漠土、高山草甸土、高山灌丛草甸土、亚高山灌丛草甸土、森林灰褐土。乔木主要有青海云杉(Piceacrassifolia)、祁连圆柏(Sabinaprzewalskii)、山杨(Populusdavidiana)等;灌木主要有沙棘(Hippophaerhamnoides)、金露梅(Potentillafruticosa)、小叶金露梅(Potentillaparvifolia)、鬼箭锦鸡儿(Caraganajubata)、银露梅(Potentillaglabra)、杯腺柳(Salixcupularis)、高山绣线菊(Spiraeaalpina)等;草本有珠芽蓼(Polygonumviviparum)、唐古特虎耳草(Saxifragatangutica)、钉柱委陵菜(Potentillasaundersiana)、甘肃棘豆(Oxytropiskansuensis)、红棕苔草(Carexprzewalski)、蒿草(Kobresiamyosuroides)、草地早熟禾(Poapratensis)、四蕊山莓草(Sibbaldiatetrandra)、短筒兔耳草(Lagotisbrevituba)等。
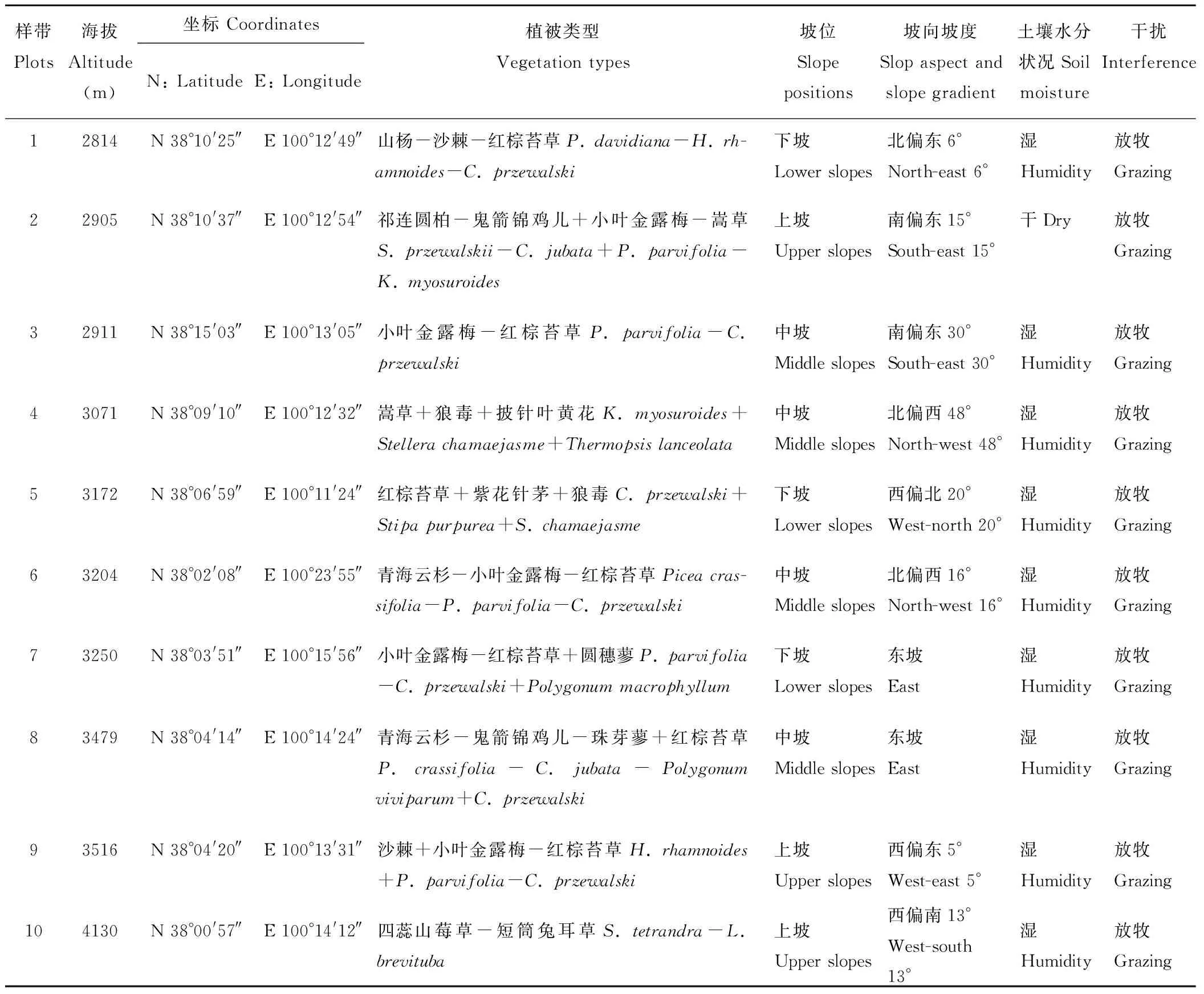
表1 样地概况
1.2 样地设置与土壤样品采集
土壤样品采集于2017年7月,在每一海拔设置3个样地,除去表层凋落物层后在剖面0~20 cm土层取土样,用于微生物分离培养的土样采用低温储运,用于土壤理化性质测定的土样自然风干。
1.3 土壤理化性质测定
土壤pH值、有机质、总有机碳、土壤全氮(TN)、全磷(TP)、总氮(TN)的测定参照《土壤分析技术规范》[18]的方法;速效磷、全钾、速效钾、铵态氮和硝态氮参照章家恩[19]的方法。
1.4 土壤微生物培养方法
各土样分别称取10.0 g 置于灭菌的装有100 mL水的三角瓶中,塞上无菌硅胶塞,在20 ℃、150 r·min-1的条件下振荡15 min,然后将土壤悬液进行梯度稀释后涂布平板上,涂布后的霉菌和放线菌在28 ℃培养5~8 d,细菌在30 ℃培养7 d,然后对每一个平板进行菌落计数,按稀释倍数换算成每克干土的菌落总数(cfu)。
真菌培养基: 10 g 葡萄糖,5 g 蛋白胨,1 g KH2PO4,0.5 g MgSO4·7H2O,0.033 g 孟加拉红,15~20 g 琼脂,溶于1000 mL去离子水中,pH自然,培养基临用前加入30 μg·mL-1链霉素[20]。
放线菌培养基: 20 g 可溶性淀粉,0.5 g MgSO4·7H2O,1 g KNO3,0.5 g NaCl,0.5 g K2HPO4,pH值7.2~7.4[22]。
解磷菌培养基: 10 g 葡萄糖,5 g Ca3(PO4)2,0.5 g MgCl2·6H2O,0.25 g MgSO4·7H2O,0.2 g KCl,0.1 g (NH4)2SO4,溶解在1000 mL去离子水中[16]。
解钾菌培养基:0.5 g 酵母膏,10 g 蔗糖,1 g (NH4)2SO4,2 g Na2HPO4,0.5 g MgSO4·7H2O,1 g CaCO3,1 g 钾长石粉(K2O·Al2O3·6SiO2),定容至1000 mL;每升富集培养基中加入15 g琼脂为选择培养基,培养基pH均为7.0[16]。
硅酸盐细菌专性培养基:5 g 蔗糖,2 g Na2HPO4,0.5 g MgSO4·7H2O,0.1 g NaCl,0.1 g CaCO3,1.0 g石英石,溶解在1000 mL去离子水中,pH为7.2[21]。
芽孢杆菌培养基:3 g 牛肉膏,5 g 蛋白胨,30 g 琼脂溶解在1000 mL去离子水中,pH为7.2,加1000 mL的麦芽汁(糖度 7°Be′)[22]。
1.5 数据统计分析
采用SPSS 16.0 软件分析土壤理化性质与土壤细菌数量的相关关系,显著性差异采用新Duncan复极法进行比较。主成分分析利用GPS 12.5软件,从样本相关矩阵出发,对不同海拔土壤养分和微生物数量的规格化特征向量进行主成分分析,根据累积方差贡献率达到85%以上确定主成分的个数。由土壤养分和微生物数量相关矩阵的特征向量,列出主成分的函数表达式,最后根据主成分值计算出不同海拔土壤养分和微生物的综合得分,对其进行比较排序。
2 结果与分析
2.1 不同海拔土壤养分
不同海拔梯度土壤养分含量见表2。土壤有机碳(SOC)、总氮(TN)、全磷(TP)随海拔的升高变化趋势明显,呈抛物线形,在海拔3250 m处达到最高,分别为119.16 g·kg-1、11.39 g·kg-1、1.62 g·kg-1。

表2 10个样地土壤养分含量Table 2 The soil nutrient contents in 10 plots
注: 不同小写字母表示不同海拔在P<0.05水平差异显著。下同。
Note: Different small letters indicate significant differences in different altitudes atP<0.05 level. The same below.
土壤中硝态氮与铵态氮含量随海拔的变化呈现不同的趋势。土壤硝态氮含量在海拔3516 m处达到最大值32.85 mg·kg-1,在海拔4130 m处为最低值1.38 mg·kg-1。而土壤铵态氮含量有两个制高点,在海拔3204 m处达到最大值14.35 mg·kg-1,在海拔3479 m处含量仅次于海拔3204 m为13.22 mg·kg-1,而土壤铵态氮含量最低点位于海拔3516 m,含量仅为0.02 mg·kg-1。
不同海拔土壤全磷含量差异不显著,变化范围为0.44~1.62 g·kg-1,其中全磷含量在3250 m处最高达到1.62 g·kg-1,3071 m处含量最低为0.44 g·kg-1;各海拔土壤速效磷含量差异显著,变化范围为0.66~4.48 mg·kg-1,在3479 m处速效磷含量最高达到4.48 mg·kg-1,在3172 m处速效磷含量最低为0.66 mg·kg-1。土壤速效磷含量与全磷含量并未呈现完全相对应的相关性。
不同海拔土壤全钾含量变化范围在13.59~28.89 g·kg-1之间,其中全钾含量在3172 m处最高为28.89 g·kg-1,2905 m处最低为13.59 g·kg-1,不同海拔土壤速效钾含量差异显著(P<0.05),变化范围为25.2~320.6 mg·kg-1,土壤速效钾含量在2911 m处最高达到320.6 mg·kg-1,3172 m处最低为25.2 mg·kg-1,在土壤速效钾含量达到最大320.6 mg·kg-1时,土壤pH(水∶土=2.5∶1)为最大的8.37,在土壤速效钾含量最低为25.2 mg·kg-1时,土壤pH为最小的6.27。
2.2 不同海拔下微生物数量的变化
由表3所示,真菌、解磷菌、解钾菌、硅酸盐细菌、芽孢杆菌随海拔梯度变化无统一规律但差异显著(P<0.05),总体变化趋势大体相同;在海拔2814~4130 m范围内微生物群落数量呈现3个波峰,分别在海拔2911、3250、3516 m处解磷菌、解钾菌、硅酸盐细菌出现峰值。3250 m处解磷菌和真菌数量最多,硅酸盐细菌数量在2911 m处最高,而芽孢杆菌在3071 m处最高,在高海拔4130 m处微生物的数量相对减少,其中硅酸盐细菌和芽孢杆菌减少程度最大,微生物总数表现为3250 m>2911 m>2814 m,即中高海拔>中低海拔>低海拔。
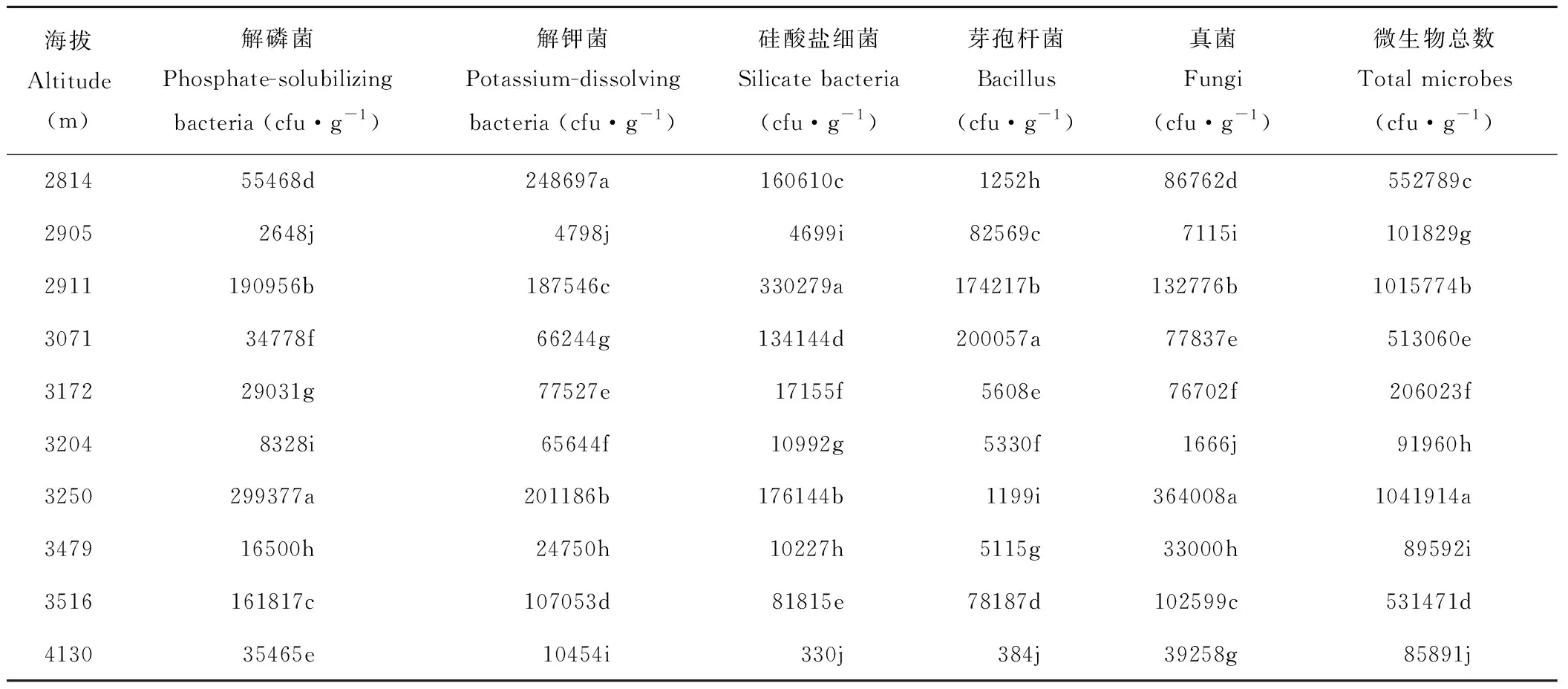
表3 海拔对微生物菌落数量的影响Table 3 The effect of elevation on the number of microbial colonies
2.3 土壤微生物量沿海拔梯度的变化与主要生态因子的相关性
对10个海拔样地的植被盖度、土壤有机碳、全N、全P、全K、硝态氮、铵态氮、速效磷、速效钾、pH与真菌、解磷菌、解钾菌、硅酸盐细菌、芽孢杆菌菌落数量进行相关性分析,其相关系数如表4所示。从表4可看出,植被盖度对微生物影响比较大,植被盖度与解钾菌、硅酸盐细菌呈极显著相关(P<0.01), 而与解磷菌、真菌呈显著相关(P<0.05);全N、全P与真菌之间存在极显著相关(P<0.01),而与解磷菌之间呈显著相关(P<0.05);pH值与各测试菌之间呈正相关,但仅与芽孢杆菌显著相关(P<0.05)。
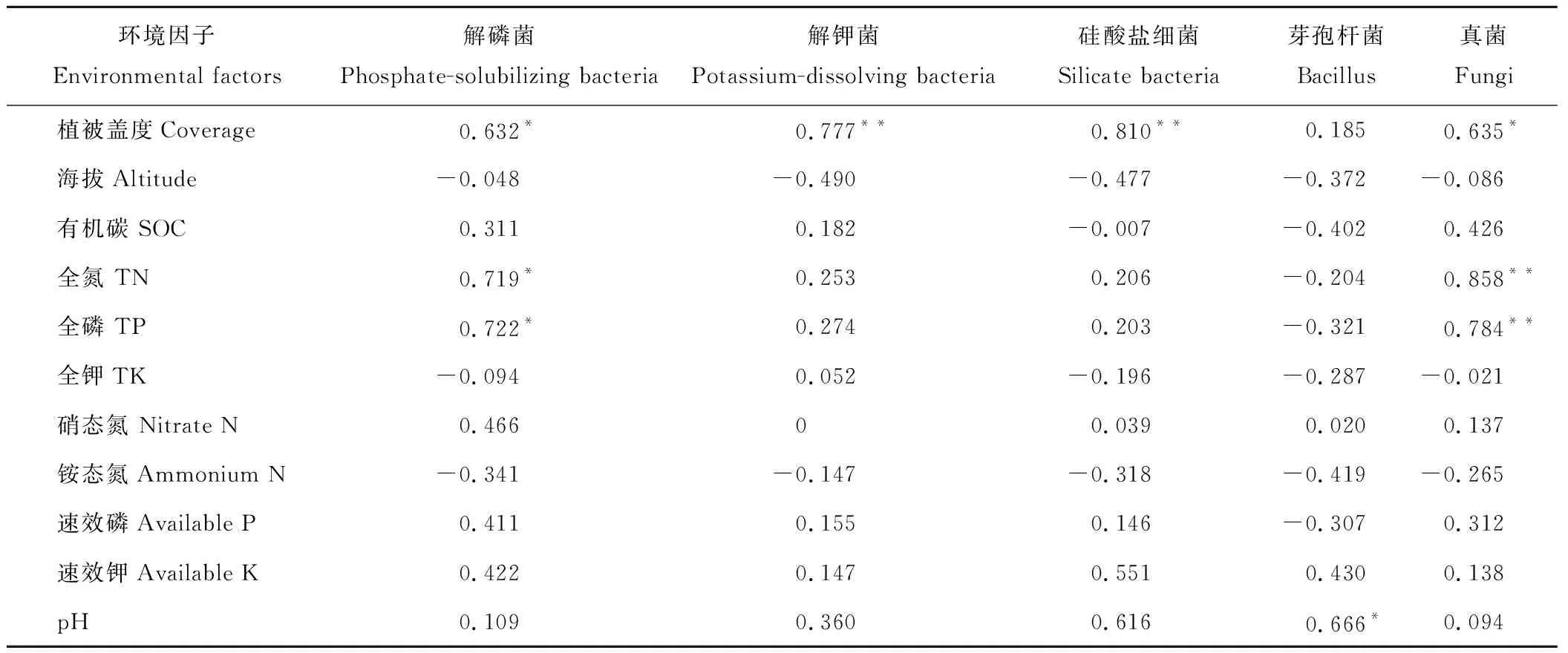
表4 不同海拔土壤可培养微生物数量与环境因子的相关性Table 4 The correlation of different altitude microbial quantity and environmental factors
注:*表示显著相关(P<0.05),**表示极显著相关(P<0.01)。下同。
Note: * indicates significant relevance atP<0.05 level, ** indicates extremely significant relevance atP<0.01 level. The same below.
2.4 不同植被类型对微生物数量的影响
不同植被类型对微生物数量的影响如表5,由表中可知,灌木林微生物总数与草地和乔木林之间差异显著(P<0.05),在选取的10个样地中,植被类型为乔木的样地分别地处海拔2814、2905、3204和3479 m处,灌木的样地分别地处海拔2911、3250和3516 m处,均属于中间海拔样地,在海拔3516 m处微生物总数最少;草地样地均处于中高海拔样地,分别为地处海拔3071、3172、4130 m处的样地,不同植被类型,其不同微生物数量随海拔升高均呈现下降趋势,微生物总数的变化趋势是:灌木>草地>乔木。

表5 不同植被类型对微生物群落的影响Table 5 The effect of vegetation type on the number of microbial colonies
2.5 不同海拔下土壤养分及微生物数量的主成分分析
2.5.1土壤养分、微生物数量的主成分分析的方差贡献率 在主成分分析当中,方差代表着土壤养分、微生物数量在主成分方向上的分散程度,方差越大,主成分在样本数据分析中的作用越大[23]。根据主成分分析法中主成分个数选取原则,从表6可见土壤养分特征值λ>1时土壤养分有3个主成分,λ分别为3.3186、2.3277、2.3277,其方差累计贡献率80.141%,表明前3个主成分已经代表了全部土壤养分80.141%的综合信息。而微生物数量特征值λ′>1时只有2个主成分,λ′分别为3.170、1.212,其方差累计贡献率87.634%。因此,土壤养分选取前3个主成分,微生物数量选取前2个主成分作为综合指标。
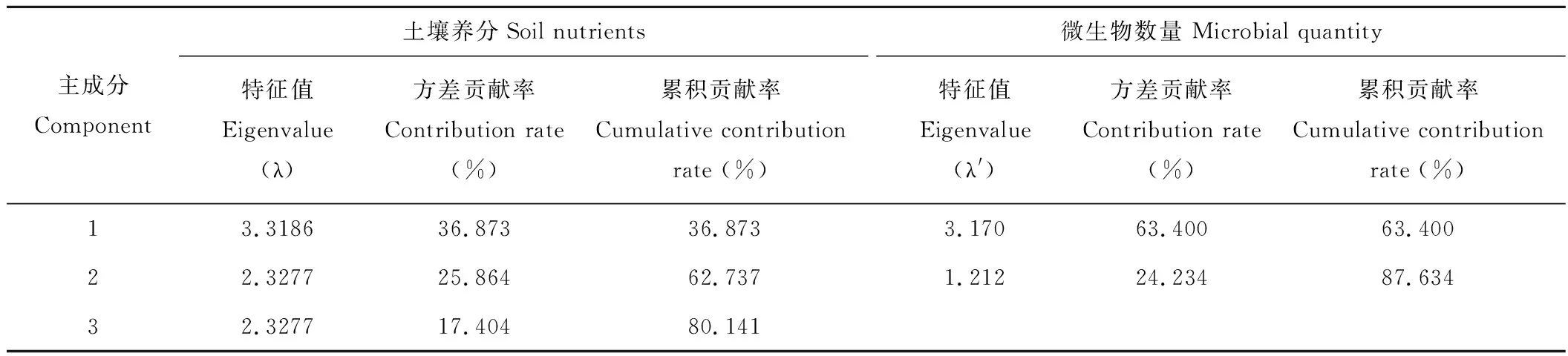
表 6 土壤养分、微生物数量的主成分分析的方差贡献率Table 6 Variance contribution rates of principal components
2.5.2不同海拔土壤养分和微生物数量的规格化特征向量 土壤养分和微生物数量的主成分载荷矩阵见表7,从表中可以看出,主成分1中有机碳、速效磷、全氮、全磷是影响土壤养分的主要因素,速效钾和pH值反映土壤养分及碱度等综合信息,而主成分3中铵态氮和硝态氮的系数较大,主成分3主要说明的是土壤速效氮的综合信息。微生物数量的主成分1中,解磷菌、硅酸盐细菌、真菌和解钾菌具有较高载荷量,反映了土壤中这些菌的综合信息,而主成分2中,仅芽孢杆菌具有较高的载荷量,表明主成分2主要反映土壤中芽孢杆菌的综合信息。

表7 不同海拔土壤养分和微生物数量的主成分载荷矩阵Table 7 The principal components load matrix of soil nutrients and microbial number at different elevations
2.5.3不同海拔土壤养分和微生物数量的主成分综合得分 根据各向量相关矩阵的特征向量(表7),土壤养分的前3个主成分及微生物数量的前2个主成分的函数表达式分别为:
F1=0.476x1+0.371x2+0.370x3-0.147x4+0.335x5+0.291x6+0.491x7+0.181x8-0.084x9
F2=-0.133x1-0.285x2-0.274x3-0.522x4+0.070x5+0.091x6+0.191x7+0.509x8+0.496x9
F3=0.161x1-0.421x2-0.441x3+0.291x4+0.246x5+0.563x6-0.017x7+0.093x8-0.363x9

土壤养分与微生物数量主成分的生物学内涵表明,上述3个土壤养分的主成分和微生物数量的2个主成分已较好地综合了土壤养分与微生物随海拔变化的主要特征。由于主成分较多,单一主成分难以对各海拔的信息综合做出客观判断。因此,以表6中土壤养分3个主成分的方差贡献率和微生物的2个主成分的方差贡献率作为权重进行加权求和,建立海拔与土壤养分及微生物数量综合评价数学模型(1)和(2):
土壤养分综合得分,F=0.460*F1+0.323*F2+0.217*F3
(1)
(2)
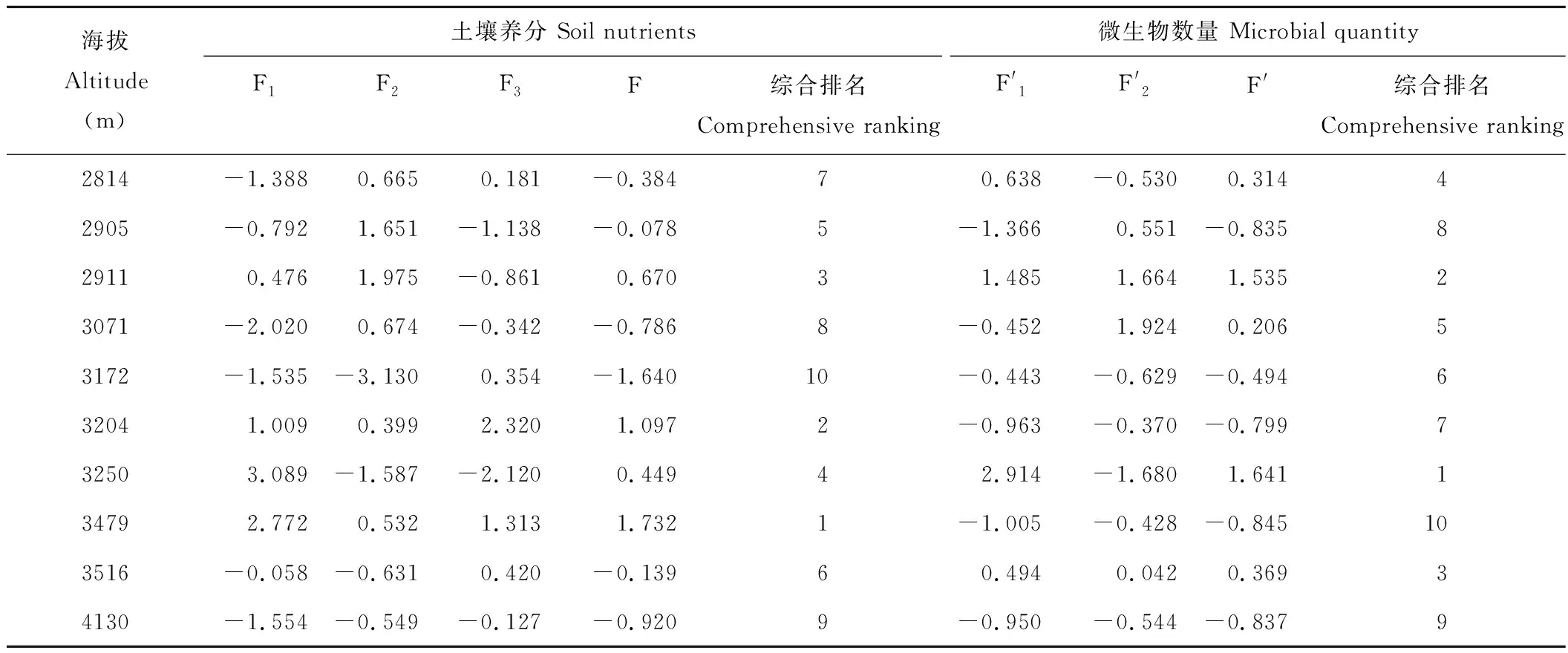
表8 冰沟流域各海拔土壤养分、微生物综合评价结果Table 8 Principal component values of soil nutrients and microbial numbers at different altitudes
冰沟流域各海拔土壤养分、微生物综合评价结果见表8,从表中可以看出,10个海拔样地在海拔3479 m处土壤养分综合评价最好,排序第一,其次为3204、2911 m处的土壤养分,在海拔3172 m处的排名最差,说明在中高海拔处的土壤肥力最好。因此通过土壤主成分综合得分可较好表征森林土壤的肥力水平的高低。 从表8还可以看出,土壤微生物数量在海拔3250 m处排序第一,2911 m处排序第二,3516 m处排序第三,3479 m处的微生物数量排序最低,这个结论与表3结果相一致。
3 讨论
土壤养分是森林生态系统中植物营养的主要来源,森林物种组成、群落结构及生产力均受土壤养分的影响,土壤养分的空间分布呈现一定的海拔梯度特征[24]。本研究结果表明,土壤养分随海拔变化差异明显,其中有机碳、全氮、全磷、硝态氮、铵态氮、速效磷在3204~3479 m处含量较高,而土壤全钾、速效钾在各海拔含量无显著规律。总体来看,垂直带土壤全氮分布与有机碳分布在海拔3204~3479 m区间显著高于在海拔3172 m以下和海拔3516 m以上,这与胡启武等[25]对祁连山北坡垂直带山地草原、森林、高山灌丛土壤有机碳和全氮的分布特征的研究结果一致。且有机碳和氮素含量均在海拔3250 m处达到最大值。在海拔3250 m处,灌丛植被覆盖率为83%,较高的植被覆盖率导致该处有机碳和氮素的含量较其他海拔处要高,而胡启武等[25]在对祁连山北坡垂直带山地草原、森林、高山灌丛土壤有机碳和全氮分布特征的研究结果也表明,土壤有机碳和全氮含量总趋势为山地草原<青海云杉林<高山灌丛,而位于该海拔处的植被类型为小叶金露梅-苔草灌丛,属于高山灌丛,是解释该海拔采样地有机碳和氮素含量高的又一有力证据。黑河流域不同海拔土壤养分变化的格局与植被类型,温、湿度,林地郁闭度和枯落物积累量有关。
土壤是一个多相、多界面的复杂系统,土壤微生物群落数的变化涉及植被类型、气候变化、土壤理化性状、凋落物分解和土壤呼吸等众多相互关联又相互影响的生物化学过程[26]。本研究结果显示,不同海拔所检测的几种微生物随海拔梯度变化而各不相同,在3250 m处解磷菌和真菌数量最多,硅酸盐细菌数量在2911 m处最高,而芽孢杆菌在3071 m处最高,在高海拔4130 m处微生物的数量相对减少,而硅酸盐细菌和芽孢杆菌减少程度最大,微生物总数为3250 m>2911 m>2814 m,即中高海拔>中低海拔>低海拔。
在不同的植被类型下微生物随海拔变化存在由高到低的变化规律,但微生物总数的变化趋势是:灌木>草地>乔木,并且均在高海拔处最低。这是因为林地土壤微生物的营养物质主要来源于地上植物的凋落物,植物根系分泌物,根的残体等。林地植被覆盖较高,从枯枝落叶分解可形成的腐殖质较多,因而有机质丰富,有充分的营养源支持微生物的生长[26]。乔木群落下土壤微生物群落最低,这与乔木植被较低的土壤有机质输入密切相关[27]。同时植被盖度对微生物群落数量也有较大的影响,不同海拔下黑河冰沟采样地微生物数量随植被盖度发生规律性变化,表现为随植被盖度的降低,微生物数量在减少,相关性分析表明,解钾菌、硅酸盐细菌、解磷菌和真菌的数量均与植被盖度正相关,作为陆地生态系统有机质的主要输入者,植被的组成往往决定了土壤微生物的群落结构特征[28]。研究表明,较适的土壤全氮、磷和土壤有机质可以促进土壤微生物的生长[29],本研究结果表明,土壤全N、全P与真菌之间相关性极显著,而与解磷菌之间显著相关,土壤pH值与芽孢杆菌呈正相关,土壤pH值影响土壤微生物的数量和分布。
研究发现不同海拔中有机碳、速效磷、全氮、全磷是构成土壤养分的主要要素,其次为速效钾、铵态氮和硝态氮,与有机碳、速效磷、全氮、全磷相关的PC1和PC2分别能解释变量方差的36.87%和25.86%,在主成分分析中起主要贡献的是有机碳、速效磷、全氮、全磷。在主成分综合评分中,土壤养分和微生物总数随海拔变化有明显的差异,在海拔3479 m处土壤的肥力最高,其次为3204和2911 m海拔处,3172 m处最低,这主要是在3479和3204 m海拔处以青海云杉、小叶金露梅和鬼箭锦鸡儿为优势种,其中青海云杉林较之其他植被带样地枯落物层较厚,枯落物可为土壤提供营养[30]。此外土壤微生物研究中,解磷菌、硅酸盐细菌、真菌和解钾菌在不同海拔的土壤中占主要优势,其次为芽孢杆菌。而微生物分值最高在3250 m处,其次为2911 m处,均为小叶金露梅灌丛植被,地形为下坡谷地和中坡,微生物数量高可能与下坡谷地部相对于中坡和坡顶跌落或下滑的枯落物和植物残体相对比较丰富有关,且土壤微生物在中高海拔处出现峰值,其研究结果与台喜生等[16]研究结果一致。因此,植被和微生物的活动可以在小尺度上影响土壤的理化性质,土壤理化性质又会影响土壤微生物群落组成和结构。
参考文献References:
[1] Chen H Z, Su D S, Lü X,etal. Dynamic characteristics of soil organic carbon and microbial biomass carbon of evergreen broad-leaved foresting Wuyi mountain. Journal of Agriculture, 2011, 1(10): 38-42.
陈涵贞, 苏德森, 吕新, 等. 武夷山常绿阔叶林土壤有机碳和微生物量碳的动态特征. 农学学报, 2011, 1(10): 38-42.
[2] Meng Y, Xue J Y, Sha L Q,etal. Variations of soil NH4-N, NO3-N and N mineralization under different forests in Xishuangbanna, Southwest China. Acta Phytoecologica Sinica, 2001, 25(1): 99-104.
孟盈, 薛敬意, 沙丽清, 等, 西双版纳不同热带森林下土壤铵态氮和硝态氮动态研究. 植物生态学报, 2001, 25(1): 99-104.
[3] Hu Z D, Liu S R, Shi Z M,etal. Variations of soil nitrogen and microbial biomass carbon and nitrogen ofQuercusaquifolioidesforest at different attitudes in Balang Mountains, Sichuan. Forest Research, 2012, 25(3): 261-268.
胡宗达, 刘世荣, 史作民, 等. 川滇高山栎林土壤氮素和微生物量碳氮随海拔变化的特征. 林业科学研究, 2012, 25(3): 261-268.
[4] Yu S, Li Y. Study on the soil microbial biomass as a bio-indicator of soil quality in the red earth ecosystem. Acta Pedologica Sinica, 1999, 36(3): 413-422.
俞慎, 李勇. 土壤微生物生物量作为红壤质量生物指标的探讨. 土壤学报, 1999, 36(3): 413-422.
[5] Peng P Q, Zhang W J, Tong C L,etal. Ertical distribution of soil organic carbon, nitrogen and microbial biomass C, N at soil profiles in wetlands of Dongting Lake floodplain. Journal of Soil and Water Conservation, 2005, 19(1): 49-53.
彭佩钦, 张文菊, 童成立, 等. 洞庭湖典型湿地土壤碳, 氮和微生物碳, 氮及其垂直分布. 水土保持学报, 2005, 19(1): 49-53.
[6] Zhang X, Zhao X, Zhang M. Functional diversity changes of microbial communities along a soil aquifer for reclaimed water recharge. FEMS Microbiology Ecology, 2012, 80(1): 9-18.
[7] Chen D D, Zhang S H, Du G Z. Soil microbial biomass and nitrogen mineralization potential along an altitudinal gradient on the Northeastern Tibetan Plateau. Journal of Lanzhou University (Natural Sciences), 2010, 46(3): 86-96.
陈懂懂, 张世虎, 杜国祯. 青藏高原东北缘不同海拔梯度土壤微生物量与氮矿化的潜力. 兰州大学学报(自然科学版), 2010, 46(3): 86-96.
[8] Richardson A E, Simpson R J. Soil microorganisms mediating phosphorus availability update on microbial phosphorus. Plant Physiology, 2011, 156(3): 989-996.
[9] Ge H L, Ji X E. Screening identification and promoting effects of potassium-solubilizing bacteria in rhizosphere of cucumber. Northern Horticulture, 2017, (13): 21-25.
葛红莲, 纪秀娥. 黄瓜根际解钾细菌的分离筛选、鉴定及其促生效果. 北方园艺, 2017, (13): 21-25.
[10] Wu Z Y, Lin W X, Chen Z F,etal. Variations of soil microbial community diversity along an elevational gradient in mid-subtropical forest. Chinese Journal of Plant Ecology, 2013, 37(5): 397-406.
吴则焰, 林文雄, 陈志芳, 等. 中亚热带森林土壤微生物群落多样性随海拔梯度的变化. 植物生态学报, 2013, 37(5): 397-406.
[11] Zhang Y, Qin J H, Zhao Y C,etal. Change of soil physical and chemical properties organic carbon and nutrients of different soil profile on different forest Binggou of Heihe basin. Journal of Soil and Water Conservation, 2013, 27(2): 126-130.
张勇, 秦嘉海, 赵芸晨, 等. 黑河上游冰沟流域不同林地土壤理化性质及有机碳和养分的剖面变化规律. 水土保持学报, 2013, 27(2): 126-130.
[12] Mao W L, Tai X S, Wu X K,etal. Altitudinal variation characteristics of the cultivable soil bacterial community on the upper reaches of the Heihe river, Qilian mountain. Journal of Glaciology and Geocryology, 2013, 35(2): 447-456.
毛文梁, 台喜生, 伍修锟, 等. 黑河上游祁连山区土壤可培养细菌群落生境的垂直分异特征. 冰川冻土, 2013, 35(2): 447-456.
[13] Wang F, Meng H F, Hou D M,etal. Relationship between soil organic carbon distribution and soil properties among three woodlands at Ice Valley in the upper reaches of Heihe basin. Pratacultural Science, 2015, 32(4): 640-646.
王凤, 孟浩峰, 侯德明, 等. 黑河上游冰沟流域3种林地土壤有机碳分布特征与土壤特性的关系. 草业科学, 2015, 32(4): 640-646.
[14] Qiu L H, Qin J H, Zhang Y. Effect of banning grazing on alpine meadow soil organic carbon, physio-chemical properties and enzyme activities in Binggou Watershed, Qilian Mountain. Agricultural Research in the Arid Areas, 2017, 35(4): 178-184.
邱丽华, 秦嘉海, 张勇. 禁牧对祁连山冰沟流域高山草甸土有机碳及理化性质和酶活性的影响. 干旱地区农业研究, 2017, 35(4): 178-184.
[15] Liu Z W, Chen R S, Song Y X,etal. Estimation of aboveground biomass for alpine shrubs in the upper reaches of the Heihe River Basin, Northwestern China. Environmental Earth Sciences, 2015, 73(9): 5513-5521.
[16] Tai X S, Yang X L, Liu G X,etal. Variations of N- and P-cycling bacterial populations along with altitude in the Qilian Mountain. Journal of Glaciology and Geocryology, 2014, 36(1): 214-221.
台喜生, 杨秀丽, 刘光琇, 等. 祁连山不同海拔氮磷循环细菌数量变化特征. 冰川冻土, 2014, 36(1): 214-221.
[17] Lu Q Z, Ma R P, Zhang L. Specien diversity of Qinghai Qilian mountain area. Qinghai Prataculture, 2009, 18(4): 24-26.
逯庆章, 马仁萍, 张灵. 青海祁连山地物种多样性分析. 青海草业, 2009, 18(4): 24-26.
[18] The National Agro-Tech Extension and Service Center. Soil analysis technology standard (2nd Ed). Beijing: China Agriculture Press, 2009.
全国农业技术推广服务中心. 土壤分析技术规范(第二版). 北京: 中国农业出版社, 2009.
[19] Zhang J E. A common experimental study of methods and techniques on ecology. Beijing: Chemical Industry Press, 2006.
章家恩. 生态学常用实验研究方法与技术. 北京: 化学工业出版社, 2006.
[20] Wu J S, Lin Q M, Huang Q Y,etal. Soil microbial biomass-method and application. Beijing: China Meteorology Press, 2006.
吴金水, 林启美, 黄巧云, 等. 土壤微生物生物量的测定方法及其应用. 北京: 气象出版社, 2006.
[21] Xiao G G, Sun D S, Cao F. Wearthering of silicate minerals by metabolites produced by silicate bacteria in culture experiments. Journal of Mineralogy and Petrology, 2013, 33(1): 8-15.
肖国光, 孙德四, 曹飞. 硅酸盐细菌代谢产物影响斜长石风化作用的模拟试验. 矿物岩石, 2013, 33(1): 8-15.
[22] Li Z G, Luo Y M, Teng Y. Research method of soil and environmental microorganism. Beijing: Science Press, 2008, 52-61.
李振高, 骆永明, 滕应. 土壤与环境微生物研究法. 北京: 科学出版社, 2008, 52-61.
[23] Yang J X, Guo B L, Zhang W H,etal. The studies of principal component analysis on the main economic character and superior variety selection of walnut. Journal of Agricultural University of Hebei, 2001, 24(4): 39-42.
杨俊霞, 郭宝林, 张卫红, 等. 核桃主要经济性状的主成分分析及优良品种选择的研究. 河北农业大学学报, 2001, 24(4): 39-42.
[24] Zhang J P, Zhang L B, Wang F Y,etal. Spatial variation of soil nutrient contents in the Jinggangshan National Nature Reserve. Soils, 2014, 46(20): 262-268.
张继平, 张林波, 王风玉, 等. 井冈山国家级自然保护区森林土壤养分含量的空间变化. 土壤, 2014, 46(20): 262-268.
[25] Hu Q W, Ouyang H, Liu X D. Distribution characteristics of soil organic carbon and total nitrogen along the altitudinal belt in the northern slope of Qilian Mountains. Journal of Mountain Science, 2006, 24(6): 654-661.
胡启武, 欧阳华, 刘贤德. 祁连山北坡垂直带土壤碳氮分布特征. 山地学报, 2006, 24(6): 654-661.
[26] Gao Y, Xu Y J, Chen W L,etal. Changes and coupling of C and N in hillslope cropland of purple soil and microbial biomass. Acta Scientiae Circumstantiae, 2014, 34(7): 1794-1800.
高扬, 徐亚娟, 陈维梁, 等. 紫色土坡耕地C, N与微生物C, N变化及其耦合特征. 环境科学学报, 2014, 34(7): 1794-1800.
[27] Xiang S M, Lu M, Xu L B,etal. Nitrogen content and microbial characteristics of forest soil of 5 stands. Journal of West China Forestry Science, 2008, 37(1): 41-45.
向仕敏, 陆梅, 徐柳斌, 等. 5种林分类型林地土壤氮含量与其土壤微生物学性质的研究. 西部林业科学, 2008, 37(1): 41-45.
[28] Liu G X, Dong X P, Zhang W,etal. The changing mechanisms of microbial number on surface soil with altitude. Journal of Glaciology and Geocryology, 2010, 32(6): 1170-1174.
刘光琇, 董小培, 张威, 等. 不同海拔表层土壤微生物数量消长的机理. 冰川冻土, 2010, 32(6): 1170-1174.
[29] Shi H X, Yu J L. Quantity of microbes in the different vegetation types on the Qinghai-Tibetan Plateau. Chinese Journal of Soil Science, 2012, 43(1): 47-51.
石红霄, 于健龙. 青藏高原不同植被类型土壤微生物数量及影响因子. 土壤通报, 2012, 43(1): 47-51.
[30] Liu B R, Zhang X Z, Hu T H,etal. Soil microbial diversity under typical vegetation zones along an elevation gradient in Helan Mountains. Acta Ecologica Sinica, 2013, 33(22): 7211-7220.
刘秉儒, 张秀珍, 胡天华, 等. 贺兰山不同海拔典型植被带土壤微生物多样性. 生态学报, 2013, 33(22): 7211-7220.
猜你喜欢
林业资源管理(2022年6期)2023-01-18
散文诗(2021年22期)2022-01-12
现代园艺(2021年23期)2021-12-01
林业勘查设计(2020年1期)2021-01-18
新农业(2020年18期)2021-01-07
环球时报(2019-05-23)2019-05-23
现代园艺(2017年21期)2018-01-03
绿色科技(2016年16期)2016-10-11
亚热带资源与环境学报(2015年1期)2015-01-22
水土保持研究(2014年4期)2014-09-21
