
磷脂酶A1辅助蛋白N端截短菌株的构建及其优化表达
2018-06-14 06:45朱昊薛正莲王洲陈阿娜杨蒙
食品与发酵工业 2018年5期
朱昊,薛正莲,2*,王洲,2,陈阿娜,2,杨蒙
1(安徽工程大学 生物与化学工程学院,安徽 芜湖,241000)2(微生物发酵安徽省工程研究中心,安徽 芜湖,241000)
磷脂酶A1是一类专一水解磷脂sn-1位酰基产生溶血磷脂和自由脂肪酸的酶类,广泛存在于各种动植物和微生物体中[1]。磷脂酶A1生物功能可归纳为3类:细胞膜结构的维护和修复;细胞内代谢机制和信号传导的调节及体内磷脂的消化[2]。 溶血磷脂可被广泛应用于食品、化妆品、饲料改良和医药等领域[3]。
苏燕南等[4]在研究粘质沙雷氏菌生产磷脂酶A1的过程中发现:在磷脂酶A1编码基因plaA的下游存在一段辅助蛋白编码序列plaS基因,它与磷脂酶A1在大肠杆菌中的高活性表达密切相关,同时显示对宿主细胞有抑制作用。进一步的研究表明,此辅助蛋白PlaS属于锚蛋白ANK家族[5]。ANK家族锚蛋白是一类广泛存在于自然界的调节蛋白质相互作用的蛋白质,在细胞生理功能的实现和调控方面具有重要意义[6]。如在铜绿假单胞菌中存在一段类似于磷脂酶辅助蛋白的基因(ANKB),这段基因和过氧化酶基因属于同一个操纵子中,且位于下游无重叠基因,属于ANK家族,ANKB蛋白位于细胞周质空间,具有信号肽,对于过氧化氢酶的表达具有重要的调控作用[7]。ALCOFORADO等[8]的研究也发现在粘质沙雷氏菌中六型分泌表达系统和辅助蛋白对其种间竞争有一定的调节作用。
为进一步揭示粘质沙雷氏菌中磷脂酶A1辅助蛋白PlaS对磷脂酶A1合成及对宿主细胞生长行为调控的机制,深入研究磷脂酶A1辅助蛋白PlaS具有重要的理论和实践指导意义。目前,国内外对磷脂酶A1辅助蛋白PlaS的相关研究报道较少。本实验室张爽等[9]在前期实验中发现辅助蛋白plaS基因在大肠杆菌表达系统中构建成功后很难表达,仅在Western Blot技术下才见有少量表达,而且对宿主菌的生长也有一定的抑制作用。国内外均有研究者通过截短氮端促进蛋白表达[10-13]。
本实验去除了plaS基因的前35个氨基酸,使其与pET-28a(+)载体连接,转化到大肠杆菌BL21(DE3)中,实现了目的蛋白的异源表达。通过对实验条件的优化,使目的蛋白达到最大表达量,为辅助蛋白胞外可溶性表达奠定了基础,也为辅助蛋白的纯化及后续功能的深入研究提供了参考。
1 材料与方法
1.1 菌株、质粒与仪器
粘质沙雷氏菌PL-06,BL21(DE3),质粒载体pET-28a(+)为本实验室保存,P28为本实验室在前期实验中将pET-28a(+)载体质粒导入BL21(DE3)表达宿主中构建的空载,SP28为本实验室在前期实验中以pET-28a(+)为载体,插入plaS基因片段,导入BL21(DE3)表达宿主中构建并保存的菌株。
本文使用宁波新芝牌超声波细胞粉碎机对细胞进行破碎。
1.2 培养基与试剂
LB培养基(1 L):酵母膏5 g、蛋白胨10 g、NaCl 10 g、H2O 1 000 mL、pH 7.0。
卡那霉素(Kan)溶液:向8 mL灭菌去离子水中加入0.5 g卡那霉素,溶解后定容至10 mL。经0.22 μm滤膜过滤,分装置于在-20 ℃保存。
100 mmol/L IPTG溶液:向40 mL灭菌去离子水中加入1.19 g 的IPTG,溶解后定容至50 mL。经0.22 μm滤膜过滤,分装置于在-20 ℃保存。
LB+Kan固体培养基:待融化的LB固体培养基冷却到一定温度时, 加入卡那霉素,使其终质量浓度为50 μg/mL。
1%琼脂糖凝胶:向20 mL 1×TAE缓冲液中加入0.2 g琼脂糖粉,加热使完全溶解,待冷却至50~60 ℃时加入1 μL EB,混匀后倒入制胶模具中插上梳子,室温冷却凝固即可。
限制性核酸内切酶BamHⅠ和HindⅢ 购自赛默飞世尔科技公司。Taq酶购于BBI生物公司,其他试剂为国产分析纯。
1.3 辅助蛋白N端截短菌株的设计和构建
通过对辅助蛋白的氨基酸序列预测与比对,发现PlaS辅助蛋白序列与锚定蛋白重组子家族(ANK家族)存在一定同源性,氨基酸序列在种间保守性较高,相似度达80%。对之前在NCBI上释放的辅助蛋白PlaS序列进行信号肽分析,发现前35AA为信号肽区域(图1)。

图1 信号肽分析Fig.1 Signal peptide analysis
为使蛋白成功表达,综合考虑后决定通过PCR方法去除前35AA。根据已知的plaS基因序列,用Primer 5.0软件设计截断N端的引物,引物序列见表1。

表1 dSP28引物序列Table 1 dSP28 Primers
注:划线部分分别为BamHⅠ、HindⅢ酶切位点。
以本实验室前期构建的SP28质粒为模板, P1,P2为引物,用Taq酶进行PCR扩增dplaS基因,扩增条件为:95 ℃ 5 min;95 ℃ 35 s;56 ℃ 35 s;72 ℃ 45 s;72 ℃ 8 min。对扩增后的基因片段进行胶回收,双酶切后与pET-28a(+)载体链接,并转化至BL21(DE3)感受态细胞中。随后挑取阳性克隆子进行PCR验证,并送上海生工生物有限公司测序,将测序正确的菌种进行保种,即得到1株构建成功的dSP28表达菌株。
1.4 dSP28诱导表达及条件优化
以P28空载为对照,从LB+Kan的平板上分别挑取P28和dSP28单菌落至100 mL LB+Kan的液体培养基中,37 ℃,200 r/min过夜培养,按2%的接种量接种至100mL LB+Kan的培养液中,至OD600nm值为0.7时加入0.2 mmol/L IPTG对其进行诱导,诱导4 h后取菌液制备蛋白样品,并进行SDS-PAGE蛋白电泳,确定目的蛋白表达部位。根据已确定的蛋白表达部位,分别对诱导时间、IPTG浓度、初始OD600nm值和温度进行优化,提高蛋白表达量。其中诱导时间分别为:2、4、6、8、10 h;IPTG诱导浓度分别为: 0、0.2、0.4、0.6、0.8、1.0 mmol/L;初始OD600nm值分别为: 0.1,0.2,0.4,0.7,0.9;温度分别为: 28、31、34、37、40、43、46 ℃。
1.5 P28、SP28与dSP28在最优表达条件下OD600nm值的测定
从LB+Kan的平板上分别挑取P28、SP28和dSP28单菌落至100 mL LB+K的液体培养基中,37 ℃,200 r/min过夜培养,按2%的接种量接种至100 mL LB+Kan的培养液中,按最佳诱导条件进行培养,每2 h取样测1次OD600nm值,并绘制发酵曲线。
1.6 SDS-PAGE蛋白电泳样品制备
1.6.1 发酵液上清蛋白的制备
(1)取20 mL菌液,用离心机于12 000 r/min离心10 min,取18 mL上清液至干净的离心管中;
(2)在4 ℃下,缓慢加入95%的乙醇18 mL;
(3)将二者充分振荡,于-20 ℃放置10 min;
(4)4 ℃,12 000 r/min,离心15 min,去上清,晾干;
(5)加入400 μL TE缓冲液重悬沉淀;
(6)从重悬沉淀中取80 μL,加入20 μL 5×Loading buffer混合后,100 ℃煮5 min;
(7)于12 000 r/min离心1 min,取上清蛋白20 μL进行蛋白电泳。
1.6.2 全菌样品的制备
取500 μL菌液,用离心机于9 000 r/min离心2 min,弃上清,加入80 μL TE,吹悬后加入20 μL 5×Loading buffer,100 ℃煮5 min,12 000 r/min,离心1 min,取上清蛋白20 μL进行蛋白电泳。
1.6.3 破碎上清蛋白的制备
(1)取20 mL发酵液,用离心机于4 000 r/min离心5 min,收集菌体沉淀;
(2)取20 mL PB缓冲液重悬菌体,于4 000 r/min离心5 min,收集菌体沉淀,重复此步骤1次;
(3)将收集到的沉淀用20 mL PB缓冲液重悬,冰浴10 min,用细胞破碎仪300 W,3 s工作,3 s停顿,超声破碎10 min;
(4)将破碎后的菌液于12 000 r/min离心10 min,取18 mL上清液至干净的离心管中;
(5)在4 ℃下,缓慢加入95%的乙醇18 mL;
(6)将二者充分振荡数分钟,于-20 ℃放置10 min;
(7)4 ℃,12 000 r/min,离心15 min,去上清,晾干;
(8)加入400 μL TE缓冲液重悬沉淀;
(9)从重悬沉淀中取80 μL,加入20 μL 5×Loading buffer混合后,100 ℃煮5 min;
(10)于12 000 r/min离心1 min,取上清蛋白20 μL进行蛋白电泳。
1.6.4 破碎菌体蛋白的制备
(1)用20 mL PB缓冲液重悬超声破碎后离心得到的沉淀;
(2)从(1)中取500 μL重悬液,用离心机9 000 r/min,离心2 min,弃上清,加入80 μL TE,吹悬后加入20 μL 5×Loading buffer 100 ℃煮5 min,12 000 r/min,离心1 min,取上清蛋白20 μL进行蛋白电泳。
2 结果与分析
2.1 dSP28菌株的构建和验证
以引物P1和P2扩增得到的片段大小为651 bp(图2),将扩增好的片段与载体相连,转化。随后,从带有Kan抗性的平板上挑取转化子,接种至5 mL LB+Kan液体培养基中过夜培养,次日,从该菌液中提取质粒作为模板,以P1,P2为引物进行PCR验证,根据琼脂糖电泳图中条带大小判断阳性克隆子(图3),并将PCR验证正确的菌株送上海生工生物有限公司测序,用DNAMAN对序列进行分析比对,序列一致则对该菌株保种,用于后续实验。

M-Marker图2 SP28中dplaS的PCR扩增Fig.2 PCR amplification gene dplaS and in SP28

M-marker; 1-菌落PCR结果图3 dSP28菌落PCR验证Fig.3 The colony PCR of dSP28
2.2 dSP28目的蛋白表达位置判断

M-marker;1、3、5、7分别是P28的全菌、发酵液上清蛋白、破碎上清蛋白、破碎菌体蛋白;2、4、6、8分别是dSP28的全菌、发酵液上清蛋白、破碎上清蛋白、破碎菌体蛋白图4 P28和dSP28SDS-PAGEFig.4 The SDS-PAGE of P28 and dSP28
按1.4的方法,分别对P28和dSP28进行诱导表达,制备蛋白样品。SDS-PAGE蛋白电泳结果(图4)表明,dSP28中目的蛋白在全菌和破碎菌体中均有较高的表达量,后续优化实验可通过全菌中目的蛋白在电泳图中相应分子量处条带粗细变化来确定优化条件。
2.3 dSP28表达条件的优化
2.3.1 诱导时间的优化
以2%的接种量将dSP28种子液接种至100mL LB+Kan的液体培养基中,至OD600nm值为0.7时加入0.2 mmol/L IPTG,于37 ℃,200 r/min对其进行诱导,在诱导后2、4、6、8、10 h分别取样制备蛋白电泳。结果(图5)可知,最佳诱导时间为加入IPTG后8 h。

M-marker;1、3、5、7-P28的诱导时间分别为:2、4、6、8 h;2、4、6、8、9dSP28的诱导时间分别为:2、4、6、8、10 h图5 诱导时间对dSP28表达的影响Fig.5 Effect of induction time on dSP28 protein expression
2.3.2 IPTG浓度的优化
以2%的接种量将dSP28种子液接种至100 mL LB+Kan的液体培养基中,至OD600nm值为0.7时,分别加入0、0.2、0.4、0.6、0.8、1.0 mmol/L的IPTG,37 ℃,200 r/min诱导8 h,制备蛋白样品,进行电泳。通过SDS-PAGE蛋白电泳图发现(图6),最佳IPTG浓度为0.2 mmol/L。
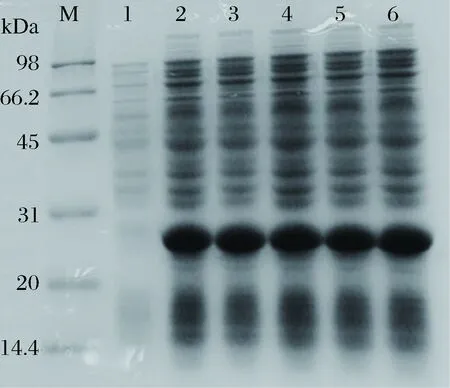
M-marker;1~6-IPTG 0、0.2、0.4、0.6、0.8、1.0 mmol/L图6 IPTG浓度对dSP28表达的影响Fig.6 Effect of IPTG concentration on dSP28 protein expression
2.3.3 初始OD600nm值的优化
以2%的接种量将dSP28种子液接种至100 mL LB+Kan的液体培养基中,在初始OD600nm值分别为: 0.1、0.2、0.4、0.7、0.9时,加入0.2 mmol/L IPTG,37 °C,200 r/min诱导8 h,制备蛋白样品。SDS-PAGE蛋白图电泳表明(图7),最佳初始OD600nm值为0.7。

M-marker;1~5-OD600nm值分别为:0.1、0.2、0.4、0.7、0.9图7 初始OD600nm值对dSP28表达的影响Fig.7 Effect of induction starting OD600nm on dSP28 protein expression
2.3.4 诱导温度的优化
以2%的接种量将dSP28种子液接种至100 mL LB+Kan的液体培养基中,在初始OD600nm值为0.7时,加入0.2 mmol/L IPTG,将诱导温度梯度设置为:28、31、34、37、40、43、46 ℃,诱导8 h,制备蛋白样品。结果发现(图8),最佳诱导温度为40 ℃。
M:marker;1~7-28、31、34、37、40、43、46 ℃图8 温度对dSP28表达的影响Fig.8 Effect of induction temperature on dSP28 protein expression
2.4 P28、SP28和dSP28发酵中OD600nm值的测定
经过表达条件的优化,得到dSP28中dPlaS蛋白的最佳表达条件:诱导时间为8 h,IPTG浓度为0.2 mmol/L,起始菌浓(OD600nm值)为0.7,温度为40 ℃,转速为200 r/min。以此表达条件对P28、dSP28和SP28诱导前后OD600nm值进行测定(图9),发现在按2%接种量接种后, 37 ℃,200 r/min,每0.5 h测1次OD600nm值时,3者菌浓差异并不明显。而加入IPTG开始诱导后,40 ℃,200 r/min,每2 h测1次OD600nm值时,SP28的菌浓先增长后降低,并趋于平稳,同时显示辅助蛋白PlaS对宿主细胞有较强的生长抑制作用。P28和dSP28则一直处于增长趋势,且增幅较大,直到终止诱导,其依旧处于上升状态。这一现象说明辅助蛋白N端截短对PlaS蛋白表达有一定的促进作用,且生长较快,对宿主菌的抑制作用大大减弱,为后面实验奠定了基础。

图9 P28、SP28和dSP28发酵中OD600nm值测定Fig.9 Determination of OD600nm in P28, SP28 and dSP28 fermentation
3 讨论
张爽等[9]在前期实验研究中发现,磷脂酶A1辅助蛋白dPlaS在大肠杆菌表达系统中构建成功后表达较为困难,只在Western Blot技术下有极少量的目的蛋白表达条带,且对构建的工程菌生长有一定的抑制作用,这可能与其信号肽有关。在本实验中对plaS基因N端截短,去除了信号肽后发现,辅助蛋白可以在细胞内进行大量表达,进行表达条件优化后,在诱导时间为8 h,IPTG浓度为0.2 mmol/L,起始菌浓(OD600nm)为0.7,温度为40 ℃,转速为200 r/min时在全菌中达到了最大表达量,而通过发酵中OD600nm值测定,发现其在最优表达条件下对宿主菌生长抑制作用明显减弱。这为后期蛋白大量胞外表达和纯化奠定了基础,也为进一步研究辅助蛋白对磷脂酶A1酶活的影响,及对辅助蛋白功能特性的深入研究提供了可能。
[1] RICHMOND G S, SMITH T K.Phospholipases A1[J].Mol Sci, 2011, 12(1):588-612.
[2] VICTOR C, DIANA M, CARLOS T, et al.Phospholipases in food industry: a review [J].Methods Mol Biol, 2012(861): 495-523.
[3] 王利.大豆磷脂酶水解技术及磷脂的HLB值测定[D].长春:中国人民解放军军需大学, 2002.
[4] 苏燕南.磷脂酶A1基因的克隆和原核表达[D].芜湖:安徽工程大学,2013.
[5] 薛正莲, 苏燕南, 王洲,等.粘质沙雷氏菌锚定蛋白重复子及其应用: CN, CN 103333229 A[P].2013.
[6] AL-KHODOR S, PRICE C T, KALIA A, et al.Functional diversity of ankyrin repeats in microbial proteins[J].Trends in Microbiology, 2010, 18(3):132-139.
[7] HOWELL M L, ALSABBAGH E, MA J F, et al.AnkB, a periplasmicankyrin-like protein inPseudomonasaeruginosa, is required for optimal catalase B (KatB) activity and resistance to hydrogen peroxide[J].Journal of Bacteriology, 2000, 182(16):4 545-4 556.
[8] ALCOFORADO D J, COULTHURST S J.Intraspecies competition inSerratiamarcescensis mediated by type VI-secreted Rhs effectors and a conserved effector-associated accessory protein[J].Journal of Bacteriology, 2015, 197(14):2 350-2 360.
[9] 张爽.粘质沙雷氏菌磷脂酶A1辅助蛋白PlaS功能特性研究[D].芜湖:安徽工程大学,2015.
[10] 程蓓蓓, 陈舜,汪铭书,等.四川白鹅CD4基因胞外去信号肽区的可溶性表达及纯化[J].四川动物,2015(6):875-879.
[11] 王明道, 聂慧慧, 王红阳,等.普鲁兰酶氮端氨基酸的截短对其酶学性质的影响[J].农业生物技术学报, 2016, 24(7):1 101-1 108.
[12] DUAN X, WU J.Enhancing the secretion efficiency and thermostability ofBacillusderamificanspullulanase mutant D437H/D503Y byN-terminal domain truncation[J].Applied & Environmental Microbiology, 2015, 81(6):1 926-1 931.
[13] GOPAL G J, KUMAR A.Strategies for the production of recombinant protein inEscherichiacoli[J].The Protein Journal, 2013, 32(6):1-17.
猜你喜欢
生物技术通报(2021年6期)2021-08-11
世界最新医学信息文摘(2021年12期)2021-06-09
昆明医科大学学报(2021年2期)2021-03-29
装备制造技术(2019年12期)2019-12-25
浙江农业学报(2017年3期)2017-04-08
中国烟草学报(2016年1期)2016-11-16
现代检验医学杂志(2016年1期)2016-11-12
天津医科大学学报(2015年2期)2015-12-22
华东理工大学学报(自然科学版)(2015年4期)2015-12-01
现代检验医学杂志(2015年2期)2015-02-06
