
白浆土玉米不同生长时期根际土壤细菌和真菌群落碳源代谢特征
2018-06-07 09:32朱珂王蕊李刚王晶修伟明赵建宁杨殿林
生态环境学报 2018年5期
朱珂 ,王蕊,李刚*,王晶,修伟明,赵建宁,杨殿林
1. 天津农学院农学与资源环境学院,天津 300384;2. 农业部环境保护科研监测所,天津 300191
土壤微生物是土壤生态系统最为活跃的组成部分,能够驱动土壤生态系统的物质和能量转化、循环和利用,保障土壤生态系统的稳定和正常运转(Mganga et al.,2016)。土壤细菌和真菌是土壤微生物的主要组成要素,共存于土壤环境中(Frey-Klett et al.,2011),是土壤生态系统中重要的分解者(Sun et al.,2016;Schimann et al.,2017),参与和调控关键的生态过程,如有机质的分解、土壤团聚体的形成和稳固等(Boddington et al.,2000;陈志豪等,2017),共同维护土壤生态系统健康。
吉林省和黑龙江省是中国玉米种植面积最大的2个省,2015年的播种面积占中国玉米总播种面积的25.24%,产量占玉米总产量的28.27%(中华人民共和国国家统计局,2016)。白浆土作为吉林省和黑龙江省的重要土壤资源,总面积约为4.18×105hm2,在玉米生产中发挥重要作用(孙元宏等,2017)。白浆土肥力低、酸性强、通透性差和易旱易涝等,被列为区域性低产土壤(贾会彬等,1997)。提升白浆土肥力以保障白浆土资源的可持续利用成为中国东北地区粮食生产亟待解决的问题。目前,关于白浆土的研究集中在机械改土方式优化以及通过秸秆还田改变重组有机碳及团聚体组成、种植绿肥和优化施肥方式提高土壤有机质含量等土壤改良措施的建立(于德清等,1993;匡恩俊等,2008;孟庆英等,2016;孙元宏等,2017),而对耕作白浆土的土壤微生物群落状况缺乏相关研究。
土壤微生物代谢特征反映土壤微生物群落功能多样性,与土壤养分元素的循环密切相关(曹永昌等,2016;王文娟等,2017)。本研究拟通过Biolog微平板技术,以中国东北玉米主产区重要的耕作土壤资源——白浆土为研究对象,探讨不同生长时期玉米根际土壤细菌和真菌群落功能多样性变化,从代谢特征的角度分析微生物群落结构的变化规律,丰富白浆土农田微生物的研究内容,为以土壤微生物资源开发和利用为基础的白浆土质量提升和可持续利用提供理论依据。
1 研究区域与研究方法
1.1 研究区概况
本研究在北京大北农生物技术有限公司吉林省试验示范基地(125°20′N,43°15′E)进行。该基地位于吉林省四平市伊通满族自治县民主村,属于中温带湿润季风气候区,年平均气温为4.6 ℃,无霜期约为138 d,年平均降雨量为627.8 mm。该地区为长白山脉向松辽平原过渡的丘陵区,其主要土地利用类型为农用坡耕地,以种植玉米为主。供试土壤为白浆土,是吉林省分布面积最大的代表性土壤类型(孙元宏等,2017),也是东北地区重要的土壤资源(孟庆英等,2016)。试验于2016年进行,在试验基地中随机选取长期种植玉米的3个小区(3次重复)进行试验,小区面积为10 m×15 m,玉米品种为当地主栽品种先玉335。于2016年4月30日播种,种植密度为每亩(1 hm2=15亩)4500株,采取垄沟种植模式,试验田施复合肥(N-P2O5-K2O:15-15-15),施用量平均为750 kg·hm-2,以基肥形式一次性施入,玉米播种后喷施乙莠合剂封闭处理,全生育期不喷施农药及不采取其他农事操作。
田间管理措施均按照当地常规操作进行。
1.2 土壤样品采集
分别于拔节期(6月 28日)、抽雄期(8月 4日)和乳熟期(8月29日)按照十字交叉法,在每个小区选取 5~10株玉米,采用抖落法收集紧紧附着在根上的土壤作为根际土。将每个生长时期同一小区收集到的所有植株的根际土充分混合均匀作为1个样品,装入自封袋中,放入低温样品储藏箱中带回实验室,共获得9份土壤样品(3个生长时期,3次重复)。将土壤样品过2 mm筛后然后分为两部分,一部分置于4 ℃冰箱中,用于土壤细菌和真菌群落代谢特征分析;另一部分用于土壤理化性质的测定。
采集土壤样品时,气候环境因子情况如下:拔节期周平均气温为 25.1 ℃,周平均降水量为 7.4 mm;抽雄期周平均气温为 25.3 ℃,周平降水量为86.8 mm;乳熟期周平均气温为16.2 ℃,周平均降水量为1.9 mm。
1.3 土壤基本理化性质测定
土壤硝态氮和铵态氮采用连续流动分析仪测定。土壤有机质、速效磷和pH测定方法见鲍士旦(鲍士旦,2008)《土壤农化分析》。基本理化性质的测定结果见表1。
1.4 土壤细菌和真菌群落代谢特征分析
土壤细菌和真菌群落生理代谢图谱(community level physiological profiling,CLPP)分别使用Biolog-ECO微平板和 Biolog-FF微平板并结合Biolog微生物自动分析仪(Biolog,Hayward,CA,USA)测定。称取5 g新鲜土壤于无菌三角瓶中,加入0.85%的灭菌NaCl溶液45 mL,封好口后,180 r·min-1振荡30 min后静置5 min,取上清液加入灭菌NaCl溶液依次稀释至10-3。使用八道移液器吸取150 μL稀释液加到Biolog-Eco和Biolog-FF微平板孔中。Biolog-Eco微平板在 28 ℃下恒温避光连续培养168 h,每隔24 h读取590 nm和750 nm波长处的吸光值;Biolog-FF微平板在25 ℃下恒温避光连续培养168 h,每隔24 h读取490 nm波长处的吸光值。本研究选取Biolog-Eco和Biolog-FF微平板培养 96 h的吸光值进行微生物群落代谢活性和多样性指数分析。96 h是是细菌和真菌生长的中间点,介于快速增长期和缓慢增长期之间。。
1.5 数据处理与统计分析
土壤微生物群落整体代谢活性采用平均颜色变化率(AWCD)表征,多样性采用Shannon指数(H′)、Simpson指数(D)和 McIntosh指数(U)和丰富度指数(S)表征(曹永昌等,2016;邹春娇等,2016),计算公式如下:
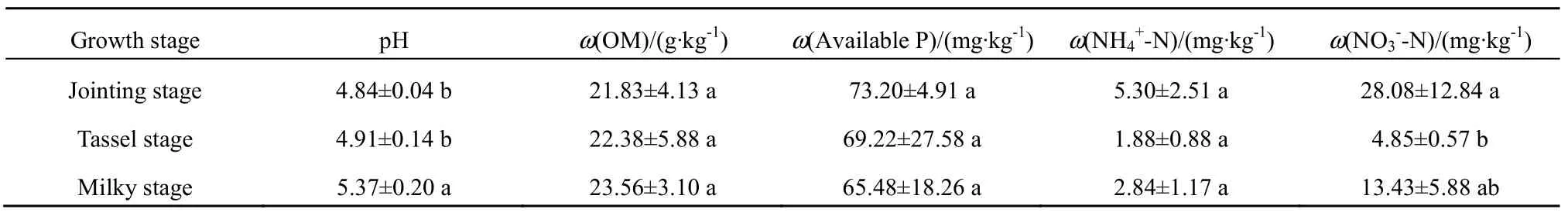
表1 玉米不同生长时期土壤基本理化性质Table 1 Basic physical-chemical properties of soils in different growth stages of maize
细菌群落代谢活性:
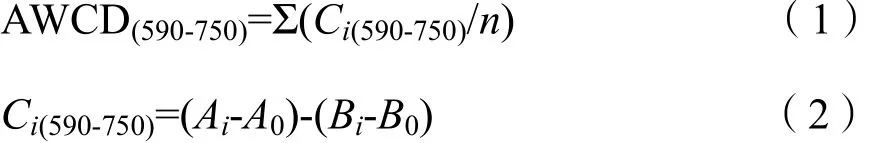
真菌群落代谢活性:
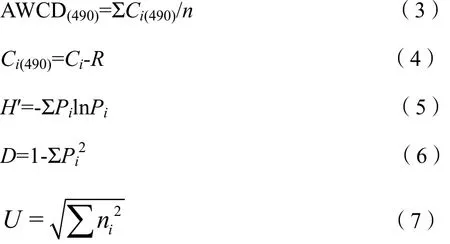
式中,n为培养基碳源的种类数,Biolog-ECO微平板的 n值为 31,Biolog-FF微平板的 n值为95;Ci(590-750)和 Ci(490)分别为细菌和真菌第 i孔的相对吸光值;Ai为第i孔在590 nm处的吸光度值;A0为对照孔在590 nm处的吸光度值;Bi为第i孔在750 nm处的吸光度值;B0为对照孔在750 nm处的吸光度值;Ci为第i孔在490 nm处的吸光值;R为对照孔在490 nm处的吸光值。Pi为第i孔的相对吸光值与整个平板相对吸光值总和的比值;ni是第i孔的相对吸光值,其中细菌为Ci(590-750),真菌为Ci(490)。
利用SPSS 22.0软件对不同生长时期土壤微生物群落代谢活性和多样性指数以及碳源相对利用率进行单因素方差分析(One-way ANOVA),并根据Duncan法进行多重比较;利用CANOCO 4.5.1软件进行主成分分析(PCA);利用Origin 9.0软件进行绘图。
2 结果与分析
2.1 平均颜色变化率(AWCD)
由图1A可知,根际土壤细菌的AWCD值随培养时间延长而逐渐增加,各处理在培养前 24 h增加均比较缓慢;培养24 h后,抽雄期根际土壤细菌的AWCD值急剧增加,而拔节期和乳熟期的AWCD值在培养48 h后才快速升高。根际土壤真菌的AWCD值变化情况与细菌相同(图1B)。对培养96 h的AWCD值进行分析,发现抽雄期根际土壤细菌的AWCD值最高,拔节期次之,乳熟期最低,抽雄期分别比拔节期和乳熟期高 26.44%和53.37%(P<0.05),拔节期和乳熟期间差异不显著(P>0.05)(图1A)。根际土壤真菌的AWCD值变化情况与细菌相同,但 3个时期间均无显著差异(P>0.05)(图 1)。
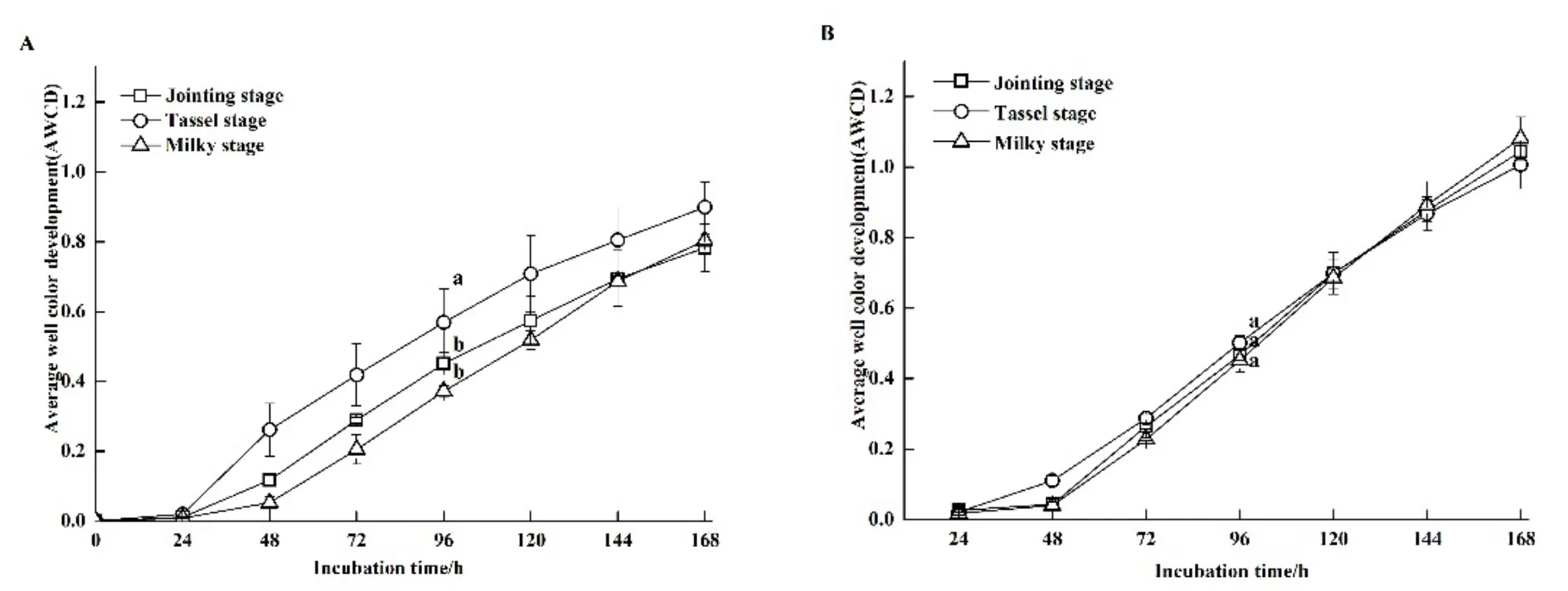
图1 根际土壤细菌(A)和真菌(B)的AWCD值随培养时间的变化Fig. 1 Change of average well color development (AWCD) of rhizosphere soil bacteria (A) and fungi (B) over incubation time
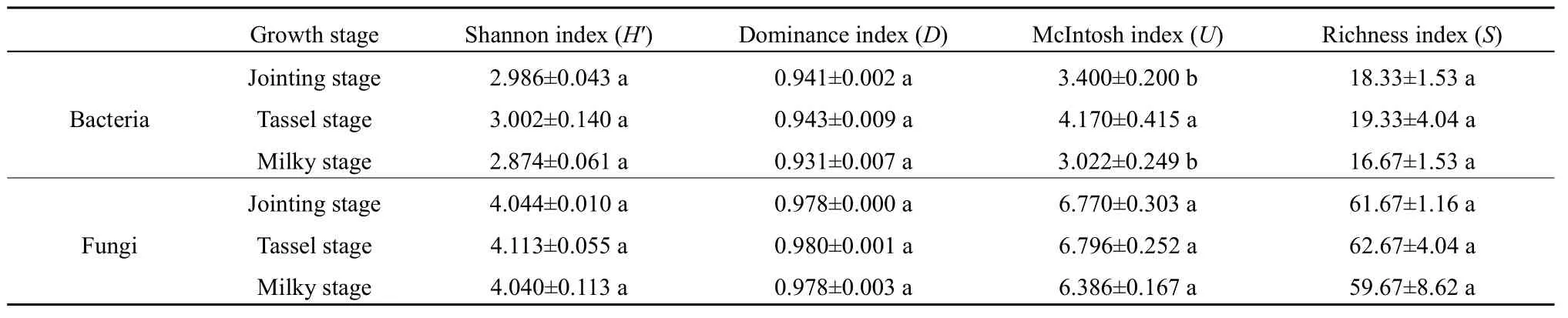
表2 不同生长时期根际土壤细菌和真菌群落代谢多样性指数Table 2 Metabolic diversity index of rhizosphere soil bacterial and fungal community in different growth stages
2.2 根际土壤细菌和真菌群落代谢多样性指数变化
由表2可知,根际土壤细菌群落的4种多样性指数随生长时期的变化表现一致,即抽雄期>拔节期>乳熟期,其中抽雄期的 McIntosh指数显著高于拔节期和乳熟期(P<0.05),拔节期与乳熟期间差异不显著(P>0.05),其他3种多样性指数在不同生长时期间差异均不显著(P>0.05)。根际土壤真菌群落的 4种多样性指数随生长时期的变化趋势同样表现为抽雄期>拔节期>乳熟期,但 3个生长时期间均无显著差异(P>0.05)。根际土壤细菌和真菌群落的 4种多样性指数变化情况均与AWCD值相似,说明随着生长时期的推进,根际土壤细菌和真菌群落的多样性指数均呈现先升高后降低的变化趋势。
2.3 根际土壤细菌和真菌群落碳源相对利用率变化
由图 2A可知,3个生长时期根际土壤细菌群落对各类碳源相对利用率最高的是糖类,占总碳源利用率的 35.90%~38.06%,氨基酸类次之,占21.39%~25.77%。对聚合物类、羧酸类、胺类和酚酸类的相对利用率较低,分别占 13.87~18.91%、8.01~9.29%、5.63~10.15%和4.39~6.88%。3个生长时期根际土壤细菌群落对同一碳源的相对利用率间均无显著差异(P>0.05),但其变化随生长时期有所不同,对糖类的相对利用率表现为逐渐降低,对羧酸类和氨基酸类的相对利用率表现为先升高后降低,而对聚合物类、酚酸类和胺类的相对利用率表现为先降低后升高。羧酸类为根际土壤真菌群落相对利用率最高的碳源,占总碳源利用率的31.17%~34.90%,糖类次之,占 25.34%~27.77%,而对氨基酸、聚合物类、胺类和其他类的相对利用率分别仅占 18.23%~18.80%、8.07%~10.15%、3.32%~5.85%和6.39%~9.96%(图2B)。3个生长时期根际土壤真菌群落对糖类、羧酸类和聚合物类的相对利用率表现为先降低后升高,对氨基酸、胺类和其他类的相对利用率表现为先升高后降低。对糖类的相对利用率随生长时期无显著变化,氨基酸类和聚合物类的分析结果与糖类相同。抽雄期对胺类和其他类的相对利用率均显著高于拔节期和乳熟期(P<0.05),拔节期和乳熟期间差异不显著(P>0.05)。而拔节期对羧酸类的相对利用率最高,显著高于抽雄期(P<0.05),但与乳熟期无显著差异(P>0.05)。
2.4 根际土壤细菌和真菌群落代谢特征的主成分分析
采用主成分分析方法分析不同时期根际土壤细菌和真菌群落在碳源利用上的差异。由图3A可知,拔节期位于第三象限,乳熟期位于第二和第三象限,而抽雄期位于第一和第四象限,说明拔节期与乳熟期根际土壤细菌群落代谢特征存在交集,而抽雄期根际土壤细菌群落代谢特征较拔节期与乳熟期发生了明显改变。根际土壤真菌群落代谢特征随生长时期的变化与细菌略有不同,但整体上趋于一致(图3B)。
3 讨论

图2 不同生长时期根际土壤细菌(A)和真菌(B)群落碳源相对利用率Fig. 2 Relative utilization rate of carbon substrate by rhizosphere soil bacterial and fungal community in different growth stages
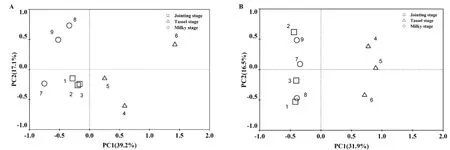
图3 根际土壤细菌(A)和真菌(B)群落代谢特征的主成分分析Fig. 3 Principal component analysis of metabolic characteristics of rhizosphere soil bacterial and fungal community
AWCD值和多样性指数能够反映土壤细菌和真菌群落代谢活性、生理状态与功能(邹春娇等,2016;唐海明等,2016)。本研究中,根际土壤细菌的 AWCD值随生长期的推进而先升高后降低,抽雄期显著高于拔节期和乳熟期(P<0.05)。分析认为,根际土壤是联系植物和微生物的纽带,是植物和微生物相互作用最为活跃的区域(Reinhold-Hurek et al.,2015),植物将多达40%的光合固定碳源以根系分泌物等形式释放到土壤环境中,为微生物提供了重要的能源物质(Hütsch et al.,2002)。拔节期、抽雄期和乳熟期是玉米的 3个代表性生长时期,玉米根系活动在此期间随植株生长不断增强,而进入生殖生长后,植株生长缓慢,根系活动减弱,这种变化导致了细菌活性的相应改变。根际土壤真菌的AWCD值变化与细菌基本一致,说明真菌活性同样受到生长时期的影响,但其变化并不显著。Harms et al.(2011)研究认为相比于细菌,真菌对生存环境变化的表现比较稳定,受环境扰动的影响较小。因此,随玉米生长而产生的根际环境变化对真菌活性没有造成显著的影响。土壤微生物多样性指数反映了微生物群落中物种的丰度及其各类型间的分布,多样性指数越高,土壤中微生物多样性越丰富,土壤微生态系统越复杂,功能越稳定(赵兰凤等,2017)。本研究中,根际土壤细菌和真菌群落的Shannon指数、Simpson指数、McIntosh指数和丰富度指数变化与AWCD值相同,说明抽雄期根际土壤细菌和真菌活性增强的同时,群落更加稳定。土壤细菌的均匀度显著升高,而土壤细菌的丰富度、优势度和利用碳源的数量无显著变化,土壤真菌的 4项多样性指标均无显著变化,这也反映了,虽然根际环境随生长时期发生改变,但细菌和真菌群落的多样性整体上未发生明显变化。
Biolog-Eco微平板上共有31种单一碳源,分为糖类(12种)、氨基酸类(6种)、羧酸类(5种)、聚合物类(4种)、胺类(2种)和酚酸类(2种)等6类(Kong et al.,2006)。Biolog-FF微平板共有95种碳源,分为糖类(44种)、氨基酸类(13种)、羧酸类(17种)、聚合物类(6种)、胺类(6种)和其他类(9种)等6类(Preston-Mafham et al.,2002)。细菌和真菌的代谢功能类群能够通过对Biolog-Eco和Biolog-FF微平板的6种类型碳源的相对利用程度来反映(张红等,2014)。本研究中,根际土壤细菌群落主要利用的碳源是糖类和氨基酸类,占总碳源利用的55%以上,这2类碳源属于易于代谢的高能量底物,最容易被农田土壤细菌所利用(Brackin et al.,2013)。而根际土壤真菌对碳源的利用与细菌有明显的差别,对羧酸类利用率最高,糖类次之。推测认为玉米根系分泌物组分及微生物对碳源利用的偏好决定了细菌和真菌碳源相对利用率的不同,但具体机理还有待深入研究。同时,本研究结果与胡国彬等(2015)发现小麦和蚕豆间作系统下红壤根际真菌对糖类的相对利用率最高的结果不同,说明土壤类型、作物种类可能是影响真菌对碳源利用的选择性的关键因素。本研究试验地白浆土具有典型的酸性强和易干旱等特点,这样的环境背景决定了真菌利用碳源的种类。根际土壤细菌和真菌群落对不同碳源的相对利用率随生长时期表现不同,分析认为这一现象与玉米根系环境变化及其与土壤微生物的相互作用有关,土壤微生物群落在适应环境变化的过程中,其对碳源的选择和利用也发生了相应的改变。同时Biolog微平板中所提供的碳源种类仍然有限,加之土壤细菌和真菌群落对碳源种类利用偏好的不同,也是造成本试验中不同生长时期根际土壤细菌和真菌群落碳源相对利用率差异的原因之一,但具体情况还有待进一步的研究。
同一种植物在不同生长时期根际微生物群落也会发生变化(Na et al.,2017)。PCA分析发现,抽雄期根际土壤细菌和真菌群落较拔节期与乳熟期发生了明显改变,而拔节期与乳熟期比较相似。抽雄期是玉米由营养生长向生殖生长过渡的时期,叶面积指数等生理指标均达到最高值,新陈代谢旺盛,根系活动及与微生物的相互作用也最为强烈,推测认为这些变化直接导致了根际土壤微生物群落结构的改变,而拔节期和乳熟期时,对根际微生物的影响相对较小,这2个时期土壤微生物群落结构虽有所不同,但差异并不明显。
本研究采用 Biolog微平板技术同时对玉米根际土壤细菌和真菌代谢特征随生长时期的变化进行了分析,为掌握白浆土微生物群落情况提供了基础数据,但仅从微生物代谢特征信息得到群落结构变化的详细信息仍比较局限,今后将采用更先进的分析方法,如宏基因组技术、宏转录组技术等,并结合更为丰富的试验设计,为中国白浆土资源的可持续利用提供技术支撑。
4 结论
(1)根际土壤细菌和真菌活性呈先升高后降低的变化趋势。抽雄期细菌活性分别比拔节期和乳熟期高26.44%和53.37%,差异显著,真菌活性无显著变化。
(2)根际土壤细菌和真菌的多样性指数变化均表现为抽雄期>拔节期>乳熟期,仅抽雄期细菌的McIntosh指数显著高于拔节期和乳熟期。
(3)糖类(35.90%~38.06%)和氨基酸类(21.39%~25.77%)是细菌群落相对利用率最高的碳源,而真菌群落相对利用率最高的碳源为羧酸类(31.17%~34.90%)和糖类(25.34%~27.77%)。根际土壤细菌和真菌对不同碳源的相对利用率随生长时期变化而不同。
(4)拔节期和乳熟期根际土壤细菌群落代谢特征相似,且均与抽雄期明显不同,真菌群落代谢特征变化与细菌相似。
致谢:感谢北京大北农生物技术有限公司吉林省试验示范基地员工在实验中给予的帮助。
BODDINGTON C L, DODD J C. 2000. The effect of agricultural practices on the development of indigenous arbuscular mycorrhizal fungi. II.Studies in experimental microcosms [J]. Plant and Soil, 218(1-2):145-157.
BRACKIN R, ROBINSON N, LAKASHMANAN P, et al. 2013. Microbial function in adjacent subtropical forest and agricultural soil [J]. Soil Biology and Biochemistry, 57(3): 68-77.
FREY-KLETT P, BURLINSON P, DEVEAU A, et al. 2011.Bacterial-fungal interactions: hyphens between agricultural, clinical,environmental, and food microbiologists [J]. Microbiology and Molecular Biology Reviews, 75(4): 583-609.
HARMS H, SCHLOSSER D, WICK L Y. 2011. Untapped potential:exploiting fungi in bioremediation of hazardous chemicals [J]. Nature Reviews Microbiology, 9(3): 177-192.
HÜTSCH B W, AUGUSTIN J, MERBACH W. 2002. Plant rhizodepositionan important source for carbon turn over in soils [J]. Journal of Plant Nutrition and Soil Science, 165(4): 397-407.
KONG W D, ZHU Y G, FU B J, et al. 2006. The veterinary antibiotic oxytetracycline and Cu influence functional diversity of the soil microbial community [J]. Environmental Pollution, 143(1): 129-137.
MGANGA K Z, RAZAVI B S, KUZYAKOV Y. 2016. Land use affects soil biochemical properties in Mt. Kilimanjaro region [J]. Catena, 141:22-29.
NA X F, XU T T, LI M, et al. 2017. Bacterial diversity in the rhizosphere of two phylogenetically closely related plant species across environmental gradients [J]. Journal of Soils and Sediments, 17(1): 122-132.
PRESTON-MAFHAM J, BODDY L, RANDERSON P F. 2002. Analysis of microbial community functional diversity using sole-carbon-source utilization profiles-a critique [J]. FEMS Microbiology Ecology, 42(1):1-14.
REINHOLD-HUREK B, BUNGER W, BURBANO C S, et al. 2015. Roots shaping their microbiome: global hotspots for microbial activity [J].Annual Review of Phytopathology, 53: 403-424.
SCHIMANN H, BACH C, LENGELLE J, et al. 2017. Diversity and structure of fungal communities in neotropical rainforest soils: the effect of host recurrence [J]. Microbial Ecology, 73(2): 310-320.
SUN R B, DSOUZA M, GILBERT J A, et al. 2016. Fungal community composition in soils subjected to long-term chemical fertilization is most influenced by the type of organic matter [J]. Environmental Microbiology, 18(12): 5137-5150.
鲍士旦. 2008. 土壤农化分析[M]. 第三版. 北京: 中国农业出版社.
曹永昌, 谭向平, 和文祥, 等. 2016. 秦岭地区不同林分土壤微生物群落代谢特征[J]. 生态学报, 36(10): 2978-2986.
陈志豪, 梁雪, 李永春, 等. 2017. 不同施肥模式对雷竹林土壤真菌群落特征的影响[J]. 应用生态学报, 28(4): 1168-1176.
胡国彬, 董坤, 董艳, 等. 2015. 小麦与蚕豆间作对根际真菌代谢功能多样性的影响[J]. 土壤学报, 52(4): 926-933.
贾会彬, 刘峰, 赵德林, 等. 1997. 白浆土某些理化特性与改良的研究[J]. 土壤学报, 34(2): 130-137.
匡恩俊, 刘峰, 贾会彬, 等. 2008. 心土培肥改良白浆土的研究Ⅰ白浆土心土培肥的效果[J]. 土壤通报, 39(5): 1160-1109.
孟庆英, 张春峰, 贾会彬, 等. 2016. 不同机械改土方式对白浆土物理特性及酶活性的影响[J]. 土壤学报, 53(2): 552-559.
孙元宏, 高雪莹, 赵兴敏, 等. 2017. 添加玉米秸秆对白浆土重组有机碳及团聚体组成的影响[J]. 土壤学报, 54(4): 1009-1017.
唐海明, 肖小平, 李微艳, 等. 2016. 长期施肥对双季稻田根际土壤微生物群落功能多样性的影响[J]. 生态环境学报, 25(3): 402-408.
王文娟, 李景文, 王中斌, 等. 2017. 胡杨根际土壤真菌群落代谢特征及其土壤影响因子研究[J].. 西北植物学报, 37(7): 1429-1437.
于德清, 丁庆堂, 关熙铭, 等. 1993. 种植牧草在白浆土改良中的作用[J]. 应用生态学报, 4(1): 37-41.
张红, 吕家珑, 曹莹菲, 等. 2014. 不同植物秸秆腐解特性与土壤微生物功能多样性研究[J]. 土壤学报, 51(4): 743-752.
赵兰凤, 张新明, 程根, 等. 2017. 生物炭对菜园土壤微生物功能多样性的影响[J]. 生态学报, 37(14): 4754-4762.
中华人民共和国国家统计局. 2016. 中国统计年鉴2016[M]. 北京: 中国统计出版社.
邹春娇, 齐明芳, 马建, 等. 2016. Biolog-ECO解析黄瓜连作营养基质中微生物群落结构多样性特征[J]. 中国农业科学, 49(5): 942-951.
猜你喜欢
中国土壤与肥料(2021年5期)2021-12-02
今日农业(2021年6期)2021-11-27
内蒙古民族大学学报(自然科学版)(2021年4期)2021-09-02
中国种业(2021年2期)2021-03-01
草地学报(2019年3期)2019-07-25
中国奶牛(2019年1期)2019-02-15
天然产物研究与开发(2018年3期)2018-05-07
中国蔬菜(2016年8期)2017-01-15
动物营养学报(2015年10期)2015-12-01
植物营养与肥料学报(2012年1期)2012-10-26
