
麻疯树主要传粉昆虫的传粉效率比较研究
2018-06-05 05:57:24罗长维
西南农业学报 2018年4期
罗长维
(1.重庆城市管理职业学院,重庆 401331;2.中国林业科学院资源昆虫研究所,云南 昆明 650224)
【研究意义】繁育系统依赖传粉昆虫的异交植物,需要有传粉者的活动才能完成受精结实。传粉昆虫对显花植物繁殖的贡献,即传粉效率(pollination effectiveness),依赖于传粉昆虫的传粉数量与传粉质量[1-2]。传粉者的传粉数量与其访问植物的频次直接相关,而传粉质量则依赖于其携粉情况、花粉沉降能力、觅食模式等[1]。传粉者携带花粉量越高,暗示着花粉转移效率更高,常代表更高的传粉效率[3]。如果传粉者携带多种植物花粉,则能造成植物柱头被异源花粉堵塞,降低传粉效率[1]。传粉者单次访花后在柱头沉降的花粉数目是衡量昆虫传粉效率高低的重要标准[3]。此外,传粉者的一些访花模式(例如对单花性别的偏好)则可能促进植物的自交或异交结实,降低或增加传粉效率[1]。因此,传粉昆虫的传粉效率与传粉数量及传粉质量密切相关,同时还受访花模式等因素的影响,但往往并不呈正相关[1]。【前人研究进展】麻疯树Jatrophacurcas为大戟科Euphorbiaceae麻疯树属灌木,亦称小桐子、膏桐(云南),原产中美洲,在我国主要分布于云南、贵州、四川等地。麻疯树种仁富含干性油,为生物质燃料的理想来源。麻疯树花单性,雌雄同株,必须依靠传粉者授粉方能结实[4]。由于受传粉限制等因素影响,自然条件下大部分麻疯树的结实率与种子产量均较低下[4]。麻疯树单花小,绿白色,花冠浅,适合蜂类、蝇类与蝶蛾类等不同种类昆虫访问[4-6]。目前有关麻疯树传粉生物学的研究,主要集中于传粉者种类、数量及其传粉行为等方面的研究[4-6]。有关麻疯树主要传粉者的传粉效率的研究,目前尚未见报道。【本研究切入点】本研究主要在对麻疯树的主要传粉者的种类、访花频率、携粉情况、花粉沉降量及访花偏好进行调查与检测的基础上,对传粉者种类及其传粉效率进行分析。【拟解决的关键问题】旨在为增加麻疯树结果率,提高种子产量提供传粉生物学方面的参考。
1 材料与方法
1.1 研究地点
试验点位于云南省永胜县热河镇(26°05.922 N,100°33.598 E,海拔1254 m),年均温23.8 ℃,年均降雨量559.4 mm。麻疯树林为15年生,株高5 m,植株间距3 m。试验点附近为面积较大且平缓的河谷谷底,同花期植物番红花Carthamustinctorius、辣椒Capsicumannuum、小白菊Chrysanthemumparthenium等。试验地附近山地为典型的萨王纳植被,散生有少量赤桉Eucalyptuscamaldulensis、剑麻Agavesisalana等植物。
1.2 研究方法
1.2.1 传粉昆虫组成 在麻疯树花期内,在传粉者的访问高峰期(早上7:00-13:00),每隔1 h对访花昆虫的种类与数量进行调查。调查中,以1个花序为观测对象,在花序2 m外对其访花昆虫进行观察,观察时间为6 min,记录访花昆虫种类与数量。并将访花者用捕虫网捕捉带回室内鉴定。每天累计观察10个花序(1序/株),连续观察6 d[6]。对麻疯树的夜间传粉昆虫也进行调查,连续6晚,仅发现1只夜蛾访问麻疯树,因此夜蛾未计入传粉昆虫种类里。最后统计麻疯树访花者每小时的访花频率,确定优势传粉昆虫种类。
1.2.2 携粉能力 为了确定传粉者携粉能力,笔者对主要传粉昆虫的携粉部位及其携粉量进行观察记录。在麻疯树林里,通过捕虫网捕捉正在访花的传粉昆虫,将其置于毒瓶内(浸有乙酸乙酯的棉球)毒杀。在扫描电子显微镜(SEM)与解剖镜下对携粉部位进行观察。在解剖镜下对不同部位的携粉量进行观察计录。昆虫体表被花蜜粘结一起的花粉块(包括蜜蜂花粉篮里)不计入携粉量,观察重复10次,统计平均值。
1.2.3 柱头花粉沉降数目 为了检测传粉昆虫的柱头花粉沉降数目,随机选择尚未开放的麻疯树花序,套上硫酸纸袋。待雌花开放后,去除纸袋。当雌花接受中华蜜蜂、迷蚜蝇、大头金蝇与紫绿蝇中的任何一种昆虫的1次访问后,摘取雌花放入1.5 mL的离心管带回实验室。麻疯树每朵雌花有3个柱头,每个柱头连接1个心皮,授粉后能否每个柱头都充足授粉会直接影响到麻疯树结籽数量。解剖镜下统计每个柱头上沉降的花粉量,并计算出该访问者1次访问给每朵雌花带来的花粉量。观察重复10次。同时,对雌花3个柱头上的花粉沉降数目由多到少进行排序,分析4种传粉者单次授粉后给单朵雌花以及雌花的3个柱头分别沉降的花粉数量。
1.2.4 访花偏好 为了检测传粉昆虫对麻疯树雌花与雄花是否有访花偏好,对中华蜜蜂、迷蚜蝇、大头金蝇与紫绿蝇对麻疯树单花的访问情况进行观察统计。在麻疯树林里,选择20株长势一致的植株,每株选1个东向的、中部的、雌花与雄花均在开放的麻疯树花序,记录其正在开放的雌花与雄花的数量。同时在传粉昆虫访花高峰期,以1 h为时间单位,观察并记录不同种类昆虫对雌雄花的访问。最后统计分析传粉昆虫对麻疯树雌花与雄花的访问是否存在性别偏好。
1.3 统计分析
中华蜜蜂、迷蚜蝇、大头金蝇与紫绿蝇的携粉量与柱头花粉沉降数目及不同柱头间的花粉沉降用单因素方差分析(one-way analysis of variance,ANOVA) 与多重比较(LSD)进行比较。携粉量与柱头花粉沉降数目进行log (x+1)的数据转换,以满足正态分布(normal distribution)与同质性(homoscedasticity)并进行单因素方差分析。所有的分析均用SPSS 22.0。

表1 麻疯树传粉昆虫种类、访花酬物及丰富度
2 结果与分析
2.1 传粉昆虫组成
在麻疯树上采集到的传粉者种类经整理鉴定,现已初步查明主要为膜翅目的蜂类,双翅目的蝇类与鞘翅目类昆虫,共3目、10种。其中膜翅目的种类最少,有2种;双翅目4种;鞘翅目4种(表1)。这些访花昆虫与印度的麻疯树访花者种类相比,除大头金蝇外,其他种类均不相同[4]。种群数量较大的昆虫为中华蜜蜂、紫绿蝇、大头金蝇与迷蚜蝇,其次为小蜜蜂与小青花金龟,数量最少的访花昆虫为家蝇、白星花金龟、赤翅甲与花萤。
中华蜜蜂与蝇类的访花酬物均为花蜜,中华蜜蜂偶尔会采集花粉(表1)。中华蜜蜂与大头金蝇访问麻疯树雌花时,喙穿过柱头裂片缝隙伸到花盘底部吸食花蜜,传粉部位主要为喙基部。其它甲虫类访花昆虫(如小青花金龟)由于咀嚼式口器无法取食花盘底部的花蜜,只能取食雄花花粉。甲虫爬过雌花时,腹部携带花粉有粘附到雌花上从而完成授粉。由于麻疯树花粉比较粘重,中华蜜蜂、蝇类与甲虫类都有经常清除体表花粉的行为。除访问麻疯树外,中华蜜蜂、大头金蝇、紫绿蝇、迷蚜蝇也访问附近的小白菊、辣椒等同花期植物。

A.喙上刚毛; B.喙基部刚毛及所携花粉; C.腹部腹板羽状毛携带花粉; D.中足刚毛携带花粉; E.蜜蜂花粉篮(椭圆所示); F.花粉篮里的花粉A. Setas on proboscis; B. PG adhered to setas on proboscis base; C. PG attached to branched hair on abdominal region; D.PG attached to setas on mid-leg; E. PG collected in corbiculas (marked with ellipse); F.PG from corbiculas图1 中华蜜蜂不同部位体毛及携粉情况 Fig.1 Setas on A. cerena and pollen grains (PG) attached to it
2.2 携粉种类
中华蜜蜂全身密被细毛,其中喙部与足部的毛为刚毛(图1A,D),可粘附少量花粉粒(图1B,D);胸部与腹部的毛为分叉绒毛,能粘附较多花粉粒(图1C)。中华蜜蜂的访花酬物通常为花蜜,偶尔采集花粉(图1E)。中华蜜蜂体表粘附的花粉(图1B~D)与花粉篮里的花粉(图1E~F)均为麻疯树花粉,没有异源花粉。
大头金蝇、紫绿蝇与迷蚜蝇均为麻疯树数量最丰富的蝇类访花昆虫。大头金蝇喙(图2 A)、足(图2 B)与腹部(图2 C)稀被短刚毛,能粘附少量花粉。喙基部深陷于头部,携带较多花粉 (图2A)。紫绿蝇的喙部(图2 D,E)与腹部(图2 F)稀被短小刚毛,喙基部容易粘附大量花粉(图2 D)。迷蚜蝇为一种食蚜蝇,喙部(图2 G)、胸部(图2 H)与腹部(图2 I)均密被长刚毛,易粘附大量花粉。3种蝇类体表粘附花粉均为麻疯树花粉,无异源花粉(图2 A,C~I)。
2.3 携粉量
中华蜜蜂、迷蚜蝇、大头金蝇与紫绿蝇的携粉量差异显著(ANOVA,F=5.951,df=3,P<0.01; 图3)。其中,中华蜜蜂与迷蚜蝇携粉量相当(P=0.34,LSD; 图3),中华蜜蜂携粉量显著大于大头金蝇(P<0.05,LSD; 图3),大头金蝇携粉量显著高于紫绿蝇(P<0.05,LSD; 图3)。可以看出,昆虫携粉量与其体型尺寸、体毛是否分叉及密集程度有关。随着其体型的增大,昆虫携粉数目相应增加;体毛分叉越多,越易携带花粉;体毛越密集,携粉量也越大。
2.4 柱头花粉沉降数目
中华蜜蜂、迷蚜蝇、大头金蝇与紫绿蝇类单次授粉后单朵雌花的麻疯树柱头沉降的花粉量差异显著(ANOVA,F=6.892,df=3,P<0.01; 图4)。其中,中华蜜蜂与迷蚜蝇在柱头沉降的花粉数量相当(P=0.52,LSD; 图4),迷蚜蝇柱头花粉沉降量显著大于大头金蝇(P<0.05,LSD; 图4),大头金蝇柱头花粉沉降量显著高于紫绿蝇(P<0.05,LSD; 图4)。可以看出,传粉昆虫沉降在柱头的花粉量与其携粉量基本上呈正相关。

A~C大头金蝇 (A喙基所携花粉, B足部, C腹部所携花粉); D~F紫绿蝇(D喙部所携花粉,E喙基所携花粉; F腹部携带花粉);G~I 迷蚜蝇(G为喙部所携花粉; H胸部腹板所携花粉; I腹末所携花粉)A-C. C. megacephala (A.PG attached to proboscis base, B.mid-leg, C. PG attached to thorax ) ; D-F L. porphyrina (D. PG attached with proboscis; E.PG attached to proboscis base; F.PG attached to thorax);G-I Milesiinae sp. (G.PG attached at proboscis base; H.PG attached with thorax; I.PG attached to abdominal end)图2 大头金蝇、紫绿蝇与迷蚜蝇不同部位体毛及携粉情况 Fig.2 Setas of three flies and PG attached to body parts
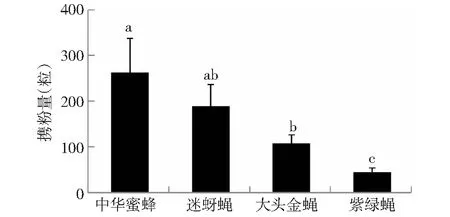
图3 主要传粉昆虫的携粉量Fig.3 Average quantity of pollen grains attached to body of A. cerena, Milesiinae sp., C.megacephala and L. porphyrina
中华蜜蜂、迷蚜蝇、大头金蝇与紫绿蝇单次访问麻疯树雌花后,沉降在单朵雌花的3个柱头的花粉量差异也非常显著(ANOVA,F=34.081vs 28.092 vs15.292 vs 39.252,df=2,表2)。可以看出,花粉量大部分沉降在1个柱头上,另外2个柱头沉降的花粉量非常少或没有。大头金蝇与紫绿蝇访问麻疯树雌花后,一些雌花的柱头、子房壁及蜜腺缝隙均粘附上黑色细屑,可能与这2种蝇食性较杂、以腐食为主相关。
2.5 访花偏好
从表3可以看出,中华蜜蜂对雄花的访问次数是雌花的6.5倍,开放雄花数量是开放雌花的5.0倍。那么中华蜜蜂对单朵雄花的访问频率为单里面是雌花的1.3倍(6.5/5.0 = 1.3)。虽然雌花位于麻疯树二歧聚伞花序的中心,在中华蜜蜂对聚伞花序的92次访问中,中华蜜蜂通常先访问雄花(N= 81次),偶尔也会先访问雌花(N=11次)。偶尔雌花比例非常高时(雄花数︰雌花数=29︰15),也会先访问雌花。而因此中华蜜蜂访花时具有一定的偏“雄性”访问。
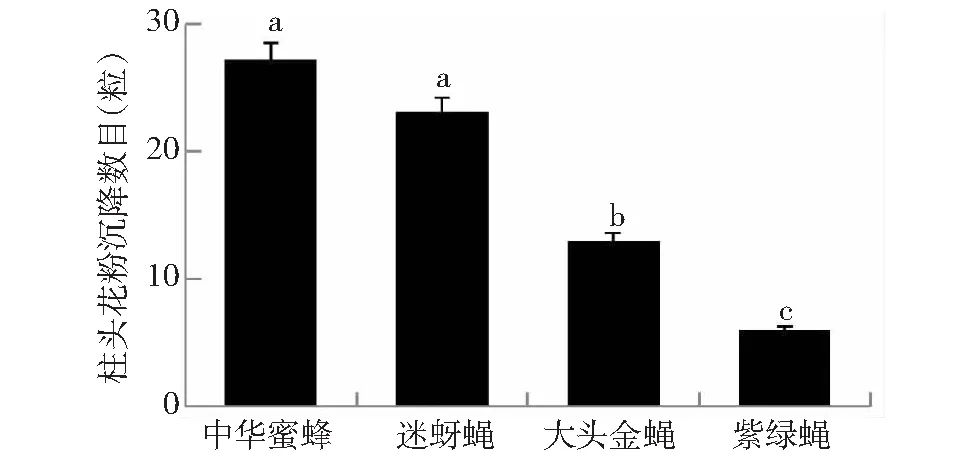
图4 传粉昆虫单次授粉后雌花柱头花粉沉降数目(平均值±标准误)Fig.4 Stigmatic pollen load of female flower after single visit of A. cerena, Milesiinae sp., C.megacephala and L. porphyrina (Mean±SE)
对蝇类对麻疯树雌雄花的访花偏好的调查结果表明:迷蚜蝇对雄花的访问次数是雌花的6.7倍,所访花序中开放雄花数量是开放雌花的6.5倍,那么迷蚜蝇对单朵雄花的访问频率是单朵雌花的1.0倍(6.7/6.5 =1.0)。大头金蝇对雄花的访问次数是雌花的5.1倍,开放雄花数量是开放雌花的5.0倍,那么大头金蝇对单朵雄花的访问频率是单朵雌花的1.0倍(5.1/5.0=1.0)。紫绿蝇对雄花的访问次数是雌花的6.4倍,所访花序中开放雄花是雌花数量的6.2倍,那么紫绿蝇对单朵雄花的访问频率是单朵雌花的1.0倍(6.4/6.2=1.0)。因此大头金蝇、迷蚜蝇与紫绿蝇3种蝇类访问麻疯树雌雄花时没有性别偏好。

表2 传粉昆虫单次授粉后雌花3个柱头花粉沉降数目
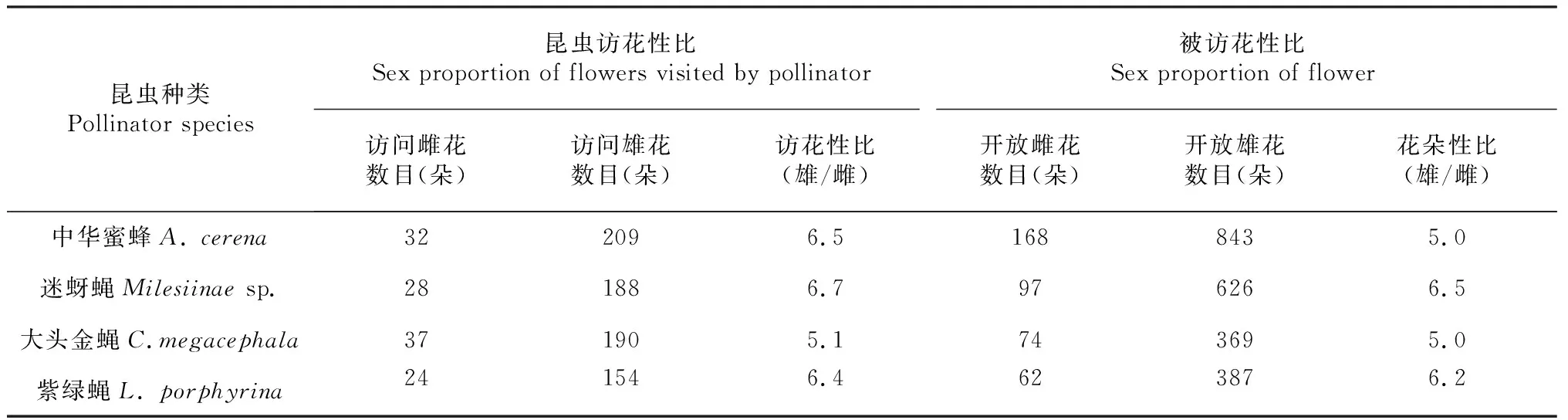
表3 传粉昆虫对麻疯树雌雄花的访花偏好
3 讨 论
3.1 主要传粉昆虫
麻疯树花冠小,花盘浅,雌雄花均能泌蜜,能接受多种短喙昆虫的访花[7]。在笔者研究里,麻疯树的访花昆虫主要为膜翅目、鞘翅目与双翅目昆虫。在印度与我国云南双柏等地,麻疯树访花昆虫还出现了蝶类与蛾类,可能暗示附近有蔬菜类植物小生境[4,6]。研究结果表明,中华蜜蜂、大头金蝇、紫绿蝇与迷蚜蝇为麻疯树的优势传粉昆虫,可能与中华蜜蜂与蝇类与人伴生的生活习性相关。中华蜜蜂比较耐热,是干热河谷地区生存相对较易的一种蜜蜂。3种传粉蝇类中,紫绿蝇与大头金蝇为粪蝇,迷蚜蝇为食蚜蝇。前者是由于干热河谷地区牛羊放牧较多,导致粪蝇数量较大。后者则由于当地大量种植油料作物番红花,蚜虫危害严重因而食蚜蝇的数量较丰富。迷蚜蝇的种群数量在番红花收获后急剧减少,因此其种群数量不太稳定。中华蜜蜂与蝇类除了访问麻疯树,也访问附近的开花作物,因此都属于泛化传粉者。
3.2 携粉种类与携粉量
中华蜜蜂与蝇类访问麻疯树与其它多种植物,可携带其它植物花粉,会导致麻疯树雌花柱头被异源花粉堵塞。笔者研究结果表明,中华蜜蜂与3种蝇类携带花粉均为麻疯树花粉,没有携带异源花粉,可能与中华蜜蜂与3种蝇类经常清除体表花粉有关。此外,中华蜜蜂还具有访花连续性flower constancy,即在一定时间内的访问行为具有一定的恒定性。中华蜜蜂这种访花连续性促进了麻疯树花粉源的同质,避免异源花粉堵塞柱头。迷蚜蝇也会经常清理体表花粉且仅访问蜜源植物,因此体表花粉组成单一。尽管大头金蝇与紫绿蝇没有携带异源花粉,但由于腐食习性会转移腐败杂质至雌花柱头,导致柱头阻塞而降低其传粉效率。
传粉昆虫体表携粉量与昆虫体型大小、体毛数量及体毛形状相关。总体而言,同一类昆虫,体型越大,携粉量越大[8],但有时不成正比[9]。传粉者体表分叉毛数目对携粉量影响最为关键,体表分叉毛越丰富,粘附花粉量越多。在本研究中,中华蜜蜂腹部独有的羽状分枝毛能粘附丰富花粉。尽管中华蜜蜂经常清除体表花粉,仍能携带相当数目的花粉。迷蚜蝇体表具大量的刚毛,能携带相当数量的花粉。大头金蝇与紫绿蝇体表毛为稀疏的刚毛,难以粘附较多花粉。
3.3 柱头花粉沉降数目
携粉量高的传粉昆虫通常可以带给雌花柱头更多的花粉,即意味着更高的传粉效率[1]。在本研究中,中华蜜蜂单次访花后单朵雌花沉降的花粉数目最高,其次为迷蚜蝇,最少的依次为大头金蝇与紫绿蝇。但是,传粉者的携粉量与其传粉效率也可能并不一致。例如,卡罗来纳黄素馨Gelsemiumsempervirens的传粉者木蜂Xylocopavirginica尽管携带大量花粉,由于其访花行为是盗蜜,因此其传粉效率显著低于另一种传粉者熊蜂Bombusbimaculatus[10]。4种传粉者均为正常访花,其中大头金蝇与紫绿蝇的柱头花粉沉降数目显著低于中华蜜蜂与迷蚜蝇,且容易携带腐败杂质导致柱头堵塞,因此二者的传粉效率明显低于中华蜜蜂与迷蚜蝇。
虫媒植物是否获得充足授粉,一方面取决于昆虫传粉效率[11],另一方面取决于植物本身,如植物柱头位置、柱头数量等[11-12]。尽管麻疯树单朵雌花的柱头仅有3个,中华蜜蜂与大头金蝇单次访花后在不同柱头间沉降的花粉数量极不均匀,部分柱头只被沉降数粒甚至没有花粉。这解释了部分麻疯树果实仅有1粒或2粒种子,而非3粒[6,13]。因此,雌花需要2次以上访问方能完成充足授粉。笔者以往的研究表明,麻疯树授粉后的雌花尽管泌蜜速率非常小,仍能继续泌蜜2 d[6]。这种授粉后继续泌蜜的特征能增加授粉不足的雌花对传粉昆虫的吸引力,促进雌花多次授粉,保证充足授粉。对自然授粉麻疯树柱头的检测也发现柱头沉降的花粉部分新鲜,部分却已变黄,表明雌花被传粉昆虫多次授粉。
3.4 访花偏好
同一植株上存在不同性别的花在显花植物中较为常见,不同性别的花对传粉昆虫的吸引并不相同。很多研究表明雄花与两性花比雌花更受传粉昆虫偏爱[12,14]。影响昆虫偏爱某种性别的花的因素很多,例如雄花花粉可作为额外的访花酬物,雌雄花的花蜜分泌量差异、雌雄花开放数量差异等[12]。在研究中,中华蜜蜂不采集麻疯树雄花花粉,但偏好访问雄花,而3种蝇类对麻疯树雌雄花没有偏好。推测可能与中华蜜蜂与蝇类对麻疯树雌雄花的访问速率有关。以往研究表明,麻疯树雄花与雌花的泌蜜速率无显著差异[6],但由于麻疯树雌花的上位子房,中华蜜蜂访问麻疯树雄花的速率是雌花的2倍,而大头金蝇对麻疯树雌花与雄花的访花速率相近[15]。此外,麻疯树单花序中开放雄花的数量明显高于开放雌花数量[4],严重偏雄花的开放式样是否会引起蜜蜂由于访花的连续性而偏雄访花也需要更多的试验进行验证。总体看来,中华蜜蜂对麻疯树雄花的偏好,无缝会导致同一花序中雌花接受自花花粉的机率增加。但是由于麻疯树的繁育系统为兼性异交,自交与异交结实率差异不大[16],因此中华蜜蜂的偏雄访花对麻疯树结实率和种子产量影响不大。
4 小 结
中华蜜蜂在4种优势传粉昆虫中具有较高的访花频率、稳定的种群数量、最高的携粉量与柱头花粉沉降数目。尽管中华蜜蜂的偏雄访花会降低麻疯树的异交结实率,由于麻疯树自交与异交结实率差异不大,对麻疯树的结实率和种子产量影响不大。因此,中华蜜蜂为麻疯树最高效的传粉昆虫,在麻疯树丰产栽培管理中,可以通过中华蜜蜂的引进与管理,增加麻疯树的传粉强度,促进麻疯树雌花授粉结实,提高麻疯树的结实率和种子产量。
致谢:双翅目昆虫由沈阳师范大学薛万琦教授鉴定;云南省林业职业技术学院周莉丽同学协助调查。
参考文献:
[1]Traveset A. Ecology of plant reproduction: mating systems and pollination[A]. In: pugnaire F I, Valladares F. Handbook of functional plant ecology[M]. New York:Marcel Dekker, 1999:545-588.
[2]Young H.Diurnal and nocturnal pollination ofSilenealba(Caryophyllaceae)[J]. American Journal of Botany, 2002, 89: 433-440.
[3]Inouye D W, Gill D E, Dudash M R, et al. A model and lexicon for pollen fate[J]. American Journal of Botany, 1994, 81: 1517-1530.
[4]Raju A J S, Ezradanam V. Pollination ecology and fruiting behaviour in a monoecious species,JatrophacurcasL.(Euphorbiaceae)[J]. Current Science India, 2002, 83: 1395-1398.
[5]Bhattacharya A, Datta K, Datta S K. Floral biology, floral resource constraints and pollination limitation inJatrophacurcasL.[J].Pakistan Journal of Biological Sciences, 2005(8): 456-460.
[6]Luo C W, Huang Z Y, Chen X M, et al. Contribution of diurnal and nocturnal insects to the pollination ofJatrophacurcas(Euphorbiaceae) in Southwest China[J]. Journal of Economic Entomology, 2011, 104: 149-154.
[7]罗长维,陈 友. 麻疯树花部特征及其对传粉昆虫的影响[J]. 南京林业大学学报(自然科学版),2017, 41(1):55-60.
[8]Robertson A W. The relationship between floral display size, pollen carryover and geitonogamy inMyosotiscolensoi(Kirk) Macbride (Boraginaceae)[J]. Biological Journal of the Linnean Society, 1992, 46: 333-349.
[9]De Vega C, M Arista, P L Ortiz, et al. The ant-pollination system ofCytinushypocistis(Cytinaceae), a Mediterranean root holoparasite[J]. Annals of Botany London, 2009, 103: 1065-1075.
[10]Adler L S, Irwin R E. Comparison of pollen transfer dynamics by multiple floral visitors: experiments with pollen and fluorescent dye[J]. Annals of Botany, 2006, 97: 141-150.
[11]Zhu X, Wan J, Li Q. Nectar robbers pollinate flowers with sexual organs hidden within corollas in distylousPrimulasecundiflora(Primulaceae)[J]. Biology Letters, 2010(10): 1098.
[12]Huang S, Tang L, Sun J, et al. Pollinator response to female and male floral display in a monoecious species and its implications for the evolution of floral dimorphism[J]. New Phytologist, 2006, 171: 417-424.
[13]Pan B, Xu Z. Benzyladenine treatment significantly increases the seed yield of the biofuel plantJatrophacurcas[J]. Journal of Plant Growth Regulation, 2011, 30: 166-174.
[14]Ashman T, Bradburn M, Cole D H, et al. The scent of a male: the role of floral volatiles in pollination of a gender dimorphic plant[J]. Ecology, 2005, 86: 2099-2105.
[15]罗长维,李 昆,陈晓鸣,等. 干热河谷麻疯树访花昆虫及主要传粉昆虫[J]. 昆虫知识,2007,45(1):121-127.
[16]Luo C W, Li K, Chen Y, et al. Floral display and breeding system ofJatrophacurcasL.[J]. Forestry Study of China, 2007(9): 114-119.
猜你喜欢
中国科学探险(2022年8期)2022-12-31 07:27:54
疯狂英语·新悦读(2022年12期)2022-03-08 02:37:12
新疆大学学报(自然科学版)(中英文)(2020年2期)2020-07-25 01:40:26
阅读与作文(小学低年级版)(2018年9期)2018-10-26 10:53:24
阅读与作文(小学低年级版)(2018年10期)2018-10-25 12:26:00
铜仁学院学报(2018年7期)2018-09-08 08:50:40
现代园艺(2017年22期)2018-01-19 05:06:57
中国稻米(2017年6期)2017-12-28 02:13:01
作物研究(2015年4期)2015-03-24 00:37:07
生物进化(2010年3期)2010-09-27 01:31:12
