
贵州三叠纪海生爬行动物群的世界遗产价值
2018-06-04 06:31向廷杰熊康宁肖时珍袁周伟
四川师范大学学报(自然科学版) 2018年3期
向廷杰, 熊康宁, 肖时珍, 袁周伟
(贵州师范大学 喀斯特研究院/贵州省喀斯特山地生态环境国家重点实验室培育基地, 贵州 贵阳 550001)
世界遗产是指被联合国教科文组织和世界遗产委员会确认的人类史上罕见的、目前无法替代的具有突出普遍价值的文物古迹和自然景观,代表了人类共同的资源与财富,需要全人类的共同保护.世界遗产包括了世界文化遗产、世界自然遗产、世界自然与文化双遗产以及文化景观,其中世界自然遗产是大自然赋予人类的宝贵财富.截止到2016年9月,全球共有世界遗产1 052处,其中文化遗产814处,自然遗产203处,自然文化混合遗产35处.由于古生物化石的形成条件特殊,地壳运动、海陆变迁、气候变化以及人类活动都会影响古生物化石的形成与储存.世界上基于世界遗产标准(viii)具有重要古生物价值的遗产地相对稀少,仅17个.我国三叠纪化石研究起步虽然较晚,但是三叠纪化石资源却是十分丰富的,研究价值也很高,尤其在我国西南地区,自1957年发现贵州龙化石以来便吸引众多国内外专家到此研究.其中,汪啸风[1]认为关岭生物群是世界上极为罕见的珍稀古生物组合,堪称为晚三叠世“化石库”.郝维城等[2]认为贵州盘县动物群是世界上中三叠世最古老的海生爬行动物类群,唯一可以与世界自然遗产地圣乔治山动物群相媲美的三叠纪动物群.马乐天[3]认为贵州兴义动物群是三叠纪生物圈海洋生态系统辐射发展阶段晚期的代表.魏家庸[4]认为贵州三叠纪景观拥有6个世界之最,如全球保存最系统全面的三叠纪海陆变迁遗迹景观,全球最丰富多彩的三叠纪海生爬行类-海百合化石群等,为贵州三叠纪自然遗产及世界地质公园申报提供理论依据.目前,贵州三叠纪海生爬行动物群申报世界自然遗产工作已经启动,已有相关专家进行考察.本文旨在运用对比分析方法,探讨贵州三叠纪海生爬行动物群的古生物多样性方面的世界遗产价值,为贵州三叠纪海生爬行动物群的申遗工作提供理论依据.
1 贵州三叠纪海生爬行动物群物种多样性
贵州三叠纪海生爬行动物群提名地位于贵州西南部,是一个隆起于四川盆地和广西丘陵之间的亚热带喀斯特高原山地分水岭.动物群形成于印支造山运动和三叠纪古特提斯洋全球海侵、海退共同作用下.其中,盘县动物群提名地位于E104°53′03.01″~E104°54′43.71″,N25°30′49.93″~N25°32′26.19″间,面积23.75 km2,为新民乡羊圈村及其邻区发现的保存在中三叠统关岭组岩层中含海生爬行类、鱼类和无脊椎动物化石的组合.兴义动物群提名地包括乌沙和顶效两个片区,其中乌沙片区位于E104°45′44.48″~E104°47′49.45″,N25°09′09.88″~N25°11′53.36″间,面积18.99 km2;顶效片区位于E105°00′13.88″~E105°01′01.04″,N25°08′42.32″~N25°09′15.15″间,面积3.56 km2,是以保存于中三叠统法郎组竹竿坡段的以贵州龙为特色的海生动物群.关岭动物群提名地位于E105°24′05.32″~E105°26′51.27″,N25°52′27.48″~N25°54′03.62″间,36.13 km2,是以新铺乡上三叠统瓦窑组灰岩中发现的以海生爬行类和海百合为主,多门类无脊椎动物共存的海生动物群.提名地总面积82.43km2,提名地核心区面积19.97km2、缓冲区面积62.46km2(图1).

(A) 盘县动物群提名地详图
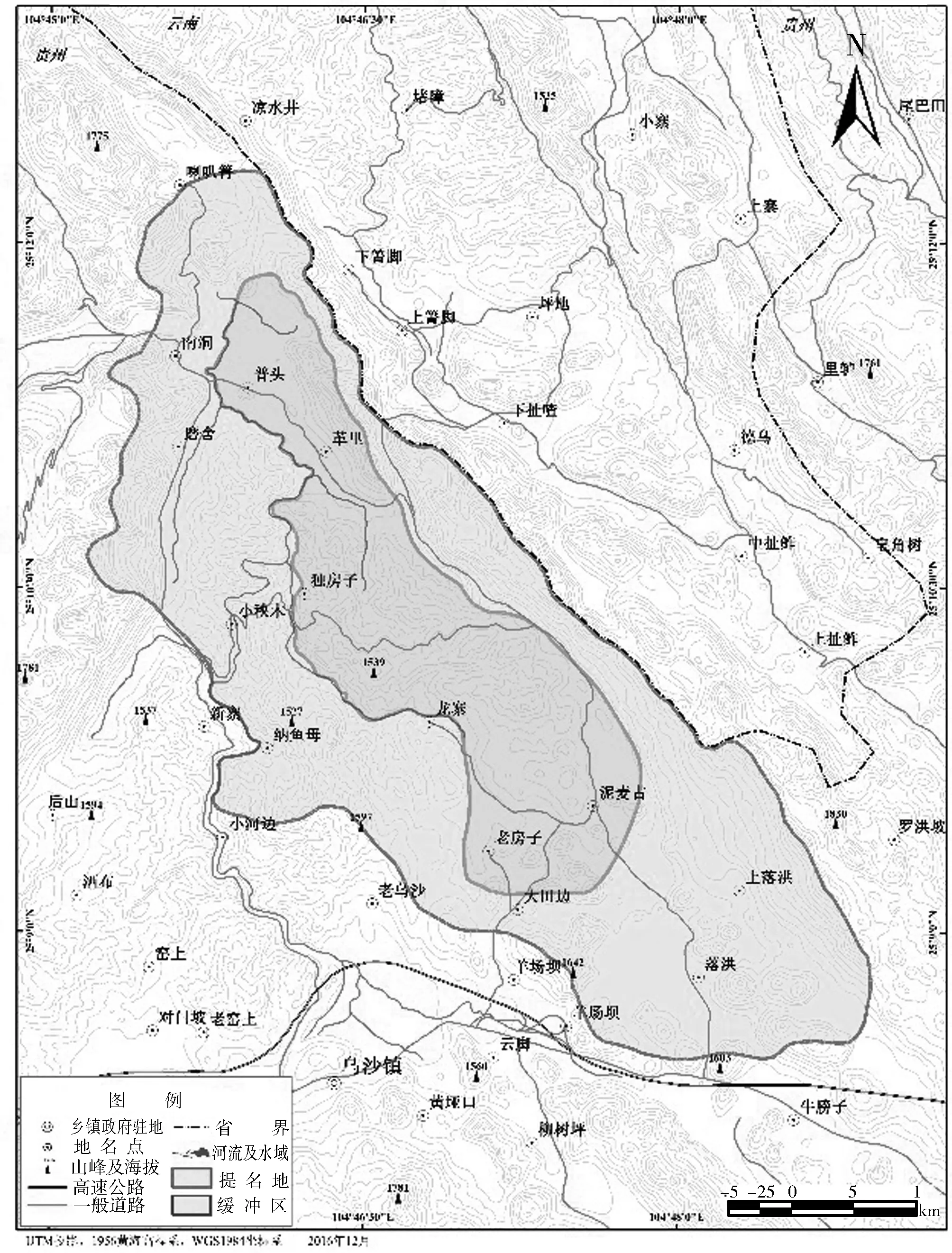
(B) 兴义动物群提名地乌沙片区详图

(C) 兴义动物群提名地顶效片区详图

(D) 关岭动物群提名地详图
图1贵州三叠纪海生爬行动物群提名地详图
Fig.1DetailedMapofthenominatedGuizhouTriassicmarinereptilefauna
贵州三叠纪海生爬行动物群的地质时代始于中三叠世安尼期中期,至晚三叠世卡尼期早期,属三叠纪东特提斯生物区(或西太平洋生物区).截止到2016年9月,贵州三叠纪海生爬行动物群共发现爬行动物40种,包括了鱼龙类、楯齿龙类、始鳍龙类、海龙类、原龙类、初龙类、龟类共7大类,基本覆盖了三叠纪时期海生爬行类群,如图2和3.除海生爬行类外,鱼类以及海百合、菊石、双壳类、腕足动物等无脊椎动物的种类繁多、数量丰富.动物群中发现了具有重要古生物学意义的特有属种.

图2 盘县中三叠统关岭组二段灰岩含化石层[5]

图3 体长1米的盘县混鱼龙化石[6]
1.1中三叠世安尼期盘县动物群盘县动物群时代早于兴义、圣乔治山动物群,处于牙形石Nicoraellakockeli带,时代为中三叠世安尼期中期,是世界上中三叠世最古老的海生爬行动物群[2].二叠纪末期生物大灭绝后,生物开始复苏,三叠纪早期发展较为缓慢,中三叠世海洋生态系统开始迅速恢复,此时的盘县动物群就是这一事件的一个自然标志和记录.动物群研究始于20世纪90年代末,在盘县新民乡羊圈村及其邻区发现了大量海生爬行类及鱼类化石,化石主要赋存在盘县新民乡关岭组上段,岩层由泥质灰岩、泥质白云岩、白云质灰岩和瘤状灰岩组成[2,7].截止到2016年9月,已经发现并报道的海生爬行类共8属10种,包括原始鱼龙类卡塔克新民龙Xinminosauruscatactes[8]、最原始的混鱼龙属——盘县混鱼龙Mixosauruspanxianensis[9]、混鱼龙属未定种[10]、始鳍龙类中最原始的鸥龙属——红果鸥龙Lariosaurushongguoensis[11]、羊圈幻龙Nothosaurusyangjuanensis[12]、乌蒙纤颌龙Wumengosaurusdelicatomandibularis[13]、小吻幻龙Nothosaurusrostellatus[14]、东特提斯首次发现的楯齿龙亚目代表意外楯齿龙Placodusinexpectatus[15]、真正适应水生生活的原龙类——东方恐头龙Dinocephalosaurusorientalis[16]和具有水生特征的初龙类——混形黔鳄Qianosuchusmixtus[17].混鱼龙和原龙类的发现表明盘县动物群具有强烈的西特提斯亲缘性,对研究三叠纪时期古海洋生物演化具有重要意义.除海生爬行动物外,还有鱼类7种以及大量海生无脊椎动物化石,其中,海生爬行类保存完整、精美,鱼类、无脊椎动物数量丰富,是一套完整的的海洋生物群落系统.
1.2中三叠世拉丁期兴义动物群兴义动物群牙形石地层控制存在争议,Zou等[18]对含兴义动物群化石地层进行了菊石层位划分,其低于或位于菊石Haocerasxingyiense带,推断时代为中三叠世拉丁期晚期.动物群主要是以产大量海生爬行类贵州龙为特色,鱼类和菊石、双壳类、海百合等无脊椎动物共存的古海洋生态遗迹,是三叠纪生物圈海洋生态系统辐射发展阶段晚期的代表[3],预示着辐射期的结束.动物群研究始于1957年贵州龙[19]的发现,是我国发现最早的三叠纪海生爬行动物群.化石主要赋存在兴义市乌沙镇、顶效镇中三叠统法郎组竹竿坡段,岩层主要由均质泥质微晶灰岩、含少量、丰富破碎生物碎屑泥质微晶灰岩、含丰富完整生物碎屑泥质微晶灰岩和水平纹层状泥晶灰岩组成[20].截止到2016年9月,已经发现并报道海生爬行类共11属12种,包括了鱼龙类黔鱼龙属兴义黔鱼龙Qianichthyosaurusxingyiensis[21]、海龙类安顺龙属乌纱安顺龙Anshunsauruswushaensis[22]、黄泥河安顺龙Aushunsaurushuangnihensis[23]、云贵地区均有发现的鳍龙类楯齿龙目康氏雕甲龟龙Glyphodermakangi[24]、始鳍龙目胡氏贵州龙Keichousaurushui[19]和岔江黔西龙Qianxisauruschajiangensis[25]、原龙类长颈龙属未定种[26]、真鳍龙亚目幻龙科杨氏幻龙Nothosaurusyoungi[27]、绿荫顶效龙Dingxiaosaurusluyinensis[28]、意外兴义龙Shingyisaurusunexpectus[29]、纯信龙次亚目李氏云贵龙Yunguisaurusliae[30]和兴义鸥龙Lariosaurusxingyiensis[31],其中兴义欧龙与欧洲的Lairosaurus头骨特征具有一致性[32],反应了东西特提斯生物群之间具有一定联系.除海生爬行类外,兴义动物群还产大量鱼类和海生无脊椎动物化石.已记述的鱼类化石约10种.其中,兴义飞翼鱼Potanichthysxingyiensis[33]是迄今发现最早的飞鱼化石,时代为中三叠世中期,优美贵州弓鳍鱼Guizhouamiabellula[34]为发现的最原始的弓鳍鱼,时代为晚三叠世早期.
1.3晚三叠世卡尼期关岭动物群关岭动物群是以保存完整、精美、属种丰富的三叠纪海生爬行动物与海百合化石为主,多门类脊椎和菊石、双壳类、腹足类等无脊椎动物共同繁盛,以及来自附近陆地的古植物化石伴生的珍稀三叠纪古生物群落组合,处于牙形石Paragandalellapolygnathiformis-Paragondolellatadpale组合代,时代为晚三叠世卡尼期早期[35],是晚三叠世古生物向海洋稳定发展演化的最重要记录.20世纪中叶发现的海百合化石是关岭古生物研究的最早记录[36],但对关岭动物群的系统研究则始于20世纪末大量三叠纪海生爬行动物化石的发现[37-38].化石主要赋存于关岭县新铺乡上三叠统竹竿坡组和小凹组,岩层主要由晶粒方解石和生物屑组成的含生物屑泥晶灰岩、微含泥质含生物屑泥晶灰岩和含泥质含粉砂质不等晶灰岩组成[39].截止到2016年9月,已经发现并报道海生爬行类共17属18种,均产自上三叠统瓦窑组,包括了尹恭正等[40]报道的鱼龙类亚洲杯锥鱼龙Cymbospondylusasiaticus、邓氏贵州鱼龙Guizhouicthyosaurustangae、关岭混鱼龙Mixosaurusguanlingensis、蔡胡氏典型鱼龙Typicusichthyosaurustsaihuae、梁氏关岭鱼龙Guanlingsaurusliangae以及海龙类孙氏新铺龙Xinpusaurussuni、黄氏新中国龙Neosinasaurushoangi、朱氏瓦窑龙Wayaosaurusgeei、美丽瓦窑龙Wayaosaurusbellus.还发现了鱼龙类周氏黔鱼龙Qianichthyosauruszhoui[37]、美丽盘江龙Panjiangsaurusepicharis[41]、卧龙岗卡洛维龙Callawayiawolonggangense[42].楯齿龙类新铺中国豆齿龙Sinocyamodusxinpuensis[43]、多板砾甲龟龙Psephochelyspolyosteoderma[44].海龙类黄果树安顺龙Anshunsaurushuangguoshuensis[38]、短吻贫齿龙Miodentosaurusbrevis[45]和双列齿凹棘龙Concavispinabiseridens[46].王立亭等[47]发现的原始龟类半甲齿龟Odontochelyssemitestacea,距今约2.2亿年前,为已知第二老的,也是唯一具有一系列过渡特征的原始龟类化石.关岭动物群也显现出了较为明显的东、西特提斯洋生物群以及东太平洋生物群特征.多板砾甲龟龙[44]、黄果树安顺龙[38]与欧洲发现的西特提斯生物区属种在结构上具有相似性,孙氏新铺龙和周氏黔鱼龙与北美的Nectosaurus、Toretocnmus具有较高亲缘性[38,48-49],反应关岭动物群与西特提斯动物群和东太平洋动物群可能均具有一定联系.除海生爬行类外,大量附着在木化石上的创口海百合化石的发现首次证明了这类海百合是营假浮游生活方式[50].
2 贵州三叠纪海生爬行动物群物种多样性的全球对比分析
2.1与古生物自然遗产地的对比截止到2016年9月,全球共有世界遗产1 052处,其中文化遗产814处,自然遗产203处,自然文化混合遗产35处.其中,基于世界遗产标准(viii)的古生物自然遗产17处.这些古生物自然遗产按时间尺度包括了古生代海洋生物和陆生动植物化石遗址、中生代海生、陆生脊椎动物和无脊椎动物、植物化石遗址以及新生代哺乳动物、植物化石遗址,共三大类(表1)[51-67].17处古生物自然遗产中,由于长远的地质历史和科技水平限制,几乎所有遗产地发掘的古生物化石在种类、数量都较为稀少,保存均不完整,多为组织和骨骼碎片,无法清晰地再现古生物及其群落特征.遗产地所显现的时间尺度或远早于三叠纪,如中国澄江化石群和加拿大米瓜莎公园,分别为寒武纪和泥盆纪,或远晚于三叠纪,如澳大利亚哺乳动物化石地和肯尼亚图尔卡纳湖国家公园,分别为第三纪和第四纪.仅有阿根廷伊莎瓜拉斯托-塔拉姆佩雅自然公园、英国多塞特和东德文海岸、意大利和瑞士联合申报的圣乔治山具有可比性(表2)[57-59].但阿根廷伊莎瓜拉斯托-塔拉姆佩雅自然公园和英国多塞特和东德文海岸的生物化石主要限于陆地生物方面,其海洋生物化石的种类、数量和质量都无法与意大利与瑞士的圣乔治山相比.因此,本文仅选取意大利和瑞士联合申报的圣乔治山作为贵州三叠纪海生爬行动物群的世界遗产对比地.

表1 基于世界遗产评价标准(viii)的古生物自然遗产地
注:参考联合国教科文组织世界遗产清单资料http://whc.unesco.org/en/list.

表2 基于世界遗产评价标准(viii)与贵州三叠纪海生爬行动物群具有可比性的世界遗产地
注:参考联合国教科文组织世界遗产清单资料http://whc.unesco.org/en/list.
意大利与瑞士联合申报的圣乔治山动物群被认为是研究三叠纪海洋生物最好的记录,地质时代为中三叠世安尼期晚期(处于安尼阶,近拉丁阶界线),5个完整、明显、重叠和连续的沉积地层记录了约1 500万a(2.45~2.30亿a前)的地质历史.在约14 km2范围内发现了超过10 000件的化石标本,产出了中三叠世约30种海生及陆生爬行动物化石,成百上千的鱼类以及双壳类、菊石、甲壳类等无脊椎动物化石,大部分保存完好,骨骼和其它形态特征得到较完整的保留,最大的化石尺寸超过7 m[59].
与圣乔治山相比,贵州三叠纪海生爬行动物群记录了从中三叠世安尼期中期至晚三叠世卡尼期早期的生物演化史,时间跨度(0.15~0.20亿a)远长于圣乔治山(0.10~0.15亿a),起始时间也早于圣乔治山(中三叠世安尼期晚期).动物群82.43 km2的总面积也广于圣乔治山,至少12个化石埋藏层的数量也远多于圣乔治山的5层.截止到2016年9月,贵州三叠纪海生爬行动物群共发现海生爬行动物40种,共7大类(鱼龙类、楯齿龙类、始鳍龙类、海龙类、原龙类、初龙类、龟类,其中初龙类为海生),基本覆盖了三叠纪时期海生爬行类群,远多于圣乔治山的30种,共5大类(鱼龙类、楯齿龙类、始鳍龙类、海龙类、原龙类,其中初龙类为陆生),且每类都不比圣乔治山少,其中有大型鱼龙化石体长超过12 m,已经超过了圣乔治山最长7 m的化石尺寸.并且发现了最原始的混鱼龙属——盘县混鱼龙Mixosauruspanxianensis、最原始的欧龙属——红果欧龙Lariosaurushongguoensis、真正适应水生生活的原龙类——东方恐头龙Dinocephalosaurusorientolis、具有水生特征的初龙类——混形黔鳄Qianosuchusmixtus以及迄今为止第二老的龟类——半甲齿龟Odontochelyssemitestacea,相比圣乔治山在研究三叠纪生物演化具有重要意义(表3).除了海生爬行类外,脊椎动物中的鱼类,无脊椎动物中的海百合、菊石、双壳类、腕足类等种类多样、数量丰富,还有不少陆生植物化石,是圣乔治山以外的低纬度地区唯一集脊椎动物、无脊椎动物和古植物化石的三叠纪化石群.其中包括了迄今最早的飞鱼化石——兴义飞翼鱼Potanichthysxingyiensis以及最原始的弓鳍鱼——优美贵州弓鳍鱼Guizhouamiabellula.关岭动物群创口海百合化石的发现证明了这类海百合是营假浮游生活方式.这些贵州三叠纪海生爬行动物群特有种的发现相较于圣乔治山具有更重要的古生物与古环境学意义.动物群不仅物种多样性和储量十分丰富,且化石大部分保存完整(达95%)、形态优美,是全球分布密集,保存完好的化石遗迹.

表3 贵州三叠纪海生爬行动物群与圣乔治山动物群海生爬行类分布表
从地层序列和反映的生物演化完整性上对比,圣乔治山反应的仅是中三叠世短时间海生动物演化过程.贵州三叠纪化石群反映的是三叠世中、晚期海生动物连续的演化史,最重要的是反映了海生爬行动物从近岸浅水区域向远洋发展的重大演化转折,从不善于游泳的盘县混鱼龙逐渐演化到具有远洋能力的真鱼龙类周氏黔鱼龙,体现了从盘县动物群到兴义动物群再到关岭动物群的海生爬行类演变过程,构成了一个沉寂-复苏-辐射-稳定的古海洋生物完整演变序列,与前者相比更具优势.从古地理上讲,圣乔治山代表的是典型的西特提斯生物区特征,贵州三叠纪海生爬行动物群3个动物群中均发现了与欧洲西特提斯生物区具有一定联系的物种[9,16,32,38,44],孙氏新铺龙和周氏黔鱼龙与北美东太平洋生物区的Nectosaurus、Toretocnmus具有较高亲缘性[38,48-49],可见其与西特提斯生物区和东太平洋生物区之间存在一定演化关系,古地理位置特殊,这也是圣乔治山无法比拟的.
在世界自然保护联盟(IUCN)对圣乔治山的评估报告中[59]明确指出中国贵州也是三叠纪海洋生命记录的重要证据,申报文本提名理由部分明确指出圣乔治山与中国贵州省发现的与瑞士相似的海生爬行动物呈现了新的、特殊的重要性.表明贵州三叠纪海生爬行动物群的重要性在世界上也是被认可的.
2.2与古生物自然遗产提名地的对比2016年度世界遗产预备名录清单中共有世界遗产提名地1 650处,其中与古生物化石相关的提名地68处,与化石相关且基于世界遗产标准(viii)的提名地共38处.这些遗产提名地展示了从早古生代寒武纪至新生代第四纪以来几乎不间断的地质与生物演化史.如爱沙尼亚的波罗的海陡崖,出露丰富的、保存完好的寒武系与奥陶系化石;美国的硅化木国家公园保存有晚三叠纪重要的古生物资源及完好的木化石.赞比亚的奇龙杜化石森林含有许多侏罗纪的超级化石,沉积层可揭示当时气候变化.韩国的恐龙化石群是世界上最大的白垩纪恐龙蛋化石和恐龙足迹化石产地;美国哥伦比亚的Tatacoa沙漠是美国最大的研究新热带动物群的脊椎动物化石产地.中国的土林-古格风景名胜区反映了喜马拉雅隆升,是地球演化重要证据,沉积层中含有丰富的哺乳动物化石.38处提名地所显现的时间尺度或远早于三叠纪,如爱沙尼亚波罗的海陡崖和澳大利亚世界遗产区的冈瓦纳雨林,分别为寒武纪和石炭纪,或远晚于三叠纪,如巴西的Serra do Divisor 国家公园,为全新世.仅有提名地美国硅化木国家公园和阿根廷Sierra de las Quijadas国家公园2处提名地的地质时间与贵州三叠纪海生爬行动物群最为接近,具有可比性(表4)[68-69].

表 4 基于世界遗产评价标准(viii)与贵州三叠纪海生爬行动物群具有可比性的世界遗产提名地
注:参考联合国教科文组织世界遗产清单资料http://whc.unesco.org/en/tentative lists
美国硅化木国家公园保存有晚三叠世重要价值的古生物资源,完好的木化石.公园地层中大范围出露精美的、保存完好的晚三叠世木化石,同时有78种动物化石被发现.但公园是以木化石为特色,动物化石保存并不完整,化石材料破碎,化石类群难以区分,难以深入研究.对比前者,贵州三叠纪海生爬行动物群中的关岭动物群晚三叠世海生动物化石数量十分丰富,保存非常完整、精美,极具美学价值,类群分类清楚,研究程度高,相比前者更具优势.与盘县、兴义动物群一起构成了一个三叠纪古海洋生物完整的演变序列,且具有众多具有重要古生物价值的特有种,在研究三叠纪古生物演化方面更具优势.
阿根廷Sierra de las Quijadas国家公园出露了三叠纪到白垩纪,从干旱到半干旱气候条件下由于沉积作用形成的连续的沉积盆地序列.地层中发现了少量的脊椎、无脊椎动物、古植物和不同类型的脊椎动物足迹化石,动物化石数量稀少,多为残破的、孤立的部分骨骼化石,植物化石则为茎、花瓣等部分组织的碎片,不能完整的反映生物演化过程. 对比前者,贵州三叠纪海生爬行动物群出露中三叠世安尼期至晚三叠世卡尼期地层,地层序列连续、完整,海生爬行类物种多样性丰富,种类齐全,完整的反映了从盘县动物群近岸不善游泳的盘县混鱼龙到拥有远洋能力的关岭动物群周氏黔鱼龙的古海洋生物演化进程,在研究物种多样性及生物演化过程上更具优势.
3 结论与讨论
1) 贵州三叠纪海生爬行动物群是世界上海生爬行动物多样性最丰富、时间跨度较长、分布范围较广的三叠纪海生爬行动物群,具有唯一性.截止到2016年9月,贵州三叠纪海生爬行动物群已发现并报道海生爬行动物7大类,40种,基本覆盖了三叠纪海生爬行类群,物种多样性优于圣乔治山.其中,还发现许多具有重要古生物学意义的海生爬行类、鱼类和无脊椎动物化石等,如盘县动物群发现的最原始的混鱼龙属、欧龙属等.兴义动物群发现的迄今最早的飞鱼和最原始的弓鳍鱼.关岭动物群发现的半甲齿龟为目前已知第二老的,唯一具有一系列过渡阶段特征的原始龟类化石,也是研究龟类甲壳形成阶段的重要化石证据,对探究龟类起源与演化具有突出价值,创口海百合化石的发现证明了这类海百合是营假浮游生活方式.贵州三叠纪海生爬行动物群记录了中三叠世安尼期中期至晚三叠世卡尼期早期(0.15~0.20亿a)的海洋生物演化进程,时间尺度上长于圣乔治山(0.10~0.15亿a),起始时间也早于世界遗产地圣乔治山(中三叠世安尼期晚期).提名地范围82.43 km2,远大于圣乔治山的14 km2.随着研究的深入,更多的海生爬行类新种、鱼类和无脊椎动物化石的发现将进一步丰富贵州三叠纪海生爬行动物群乃至世界三叠纪海生动物多样性,其古生物价值也将更为突出.
2) 贵州三叠纪海生爬行动物群物种多样性组成构成了一个沉寂-复苏-辐射-稳定的古海洋生物完整演变序列,在研究三叠纪古海洋生物演化方面具有突出普遍价值,是研究三叠纪中、晚期海洋生物发展演化的最好的记录.盘县动物群是世界已知的三叠纪最古老的动物群[2],反映了二叠纪末期生物大绝灭后生物沉寂期结束,海生爬行类和鱼类开始辐射并已有较高的分异度,代表了海生爬行类和鱼类处于演化辐射高潮期.兴义动物群是三叠纪古生物从陆地向海洋发展辐射阶段晚期的演化的代表地[3],预示着辐射期结束.关岭动物群是晚三叠世古生物向海洋洋稳定发展演化的最重要的记录,填补了世界上晚三叠世无重要海洋生物演化记录的空白.此外,贵州三叠纪海生爬行动物群多属种具西特提斯动物群特征甚至东太平洋特征[9,16,32,38,44,48-49],未来对于这些重要物种的研究极有可能揭开三叠纪三大生物区海洋生物演化之谜.
3) 贵州三叠纪海生爬行动物群物种多样性具有世界遗产的突出普遍价值,满足世界遗产标准 (viii):是地球演化史中重要阶段的突出例证,包括生命记录和地貌演变中的重要的地质过程或显著的地质或地貌特征.通过与世界上重要的古生物遗产地和提名地进行对比分析得出:贵州三叠纪海生爬行动物群是世界上海生爬行动物多样性最丰富、时间跨度较长、分布范围较广的三叠纪海生爬行动物群,具有唯一性.贵州三叠纪海生爬行动物群物种多样性组成构成了一个沉寂-复苏-辐射-稳定的古海洋生物完整演变序列,具有唯一性.为研究三叠纪古海洋生物演化提供了重要证据,具有突出普遍价值,是研究三叠纪中、晚期海洋生物发展演化的最好的记录.此前,汪啸风[1]、魏家庸[4]和王砚耕等[70]已探讨过贵州三叠纪动物群的古生物学价值,主要涉及其物种多样性和古地理环境方面,本文结合前人研究成果探讨贵州三叠纪海生爬行动物群的突出普遍价值,为其申报世界自然遗产提供理论依据.
[1] 汪啸风. 贵州关岭生物群研究的进展和存在问题(代序)[J]. 地质通报,2003,22(4):221-227.
[2] 郝维城,孙园林,江大勇,等. 盘县动物群研究进展[J]. 北京大学学报(自然科学版),2006,42(6):817-823.
[3] 马乐天,季承,孙作玉,等. 贵州兴义三叠纪海生爬行动物群的地层分布及其生物多样性[J]. 地层学杂志,2013,37(2):178-185.
[4] 魏家庸. 贵州三叠纪自然遗产及世界地质公园申报[J]. 贵州地质,2008,25(4):241-246.
[6] 李锦玲,金帆. 畅游在两亿年前的海洋:华南三叠纪海生爬行类和环境巡礼[M]. 北京:科学出版社,2009:64-65.
[7] 万大学. 贵州盘县羊圈地区安尼锡克中期凝灰岩的发现及其意义[J]. 贵州地质,2002,19(2):77-81.
[8] JIANG D Y, MOTANI R, HAO W C, et al. New primitive ichthyosaurian (Reptilia, Diapsida) from the Middle Triassic of Panxian, Guizhou, southwestern China and its position in the Triassic biotic recovery[J]. Progress in Natural Science,2008,18(10):1315-1319.
[9] JIANG D Y, SCHMITZ L, HAO W C, et al. A new mixosaurid ichthyosaur from the Middle Triassic of China[J]. J Vertebrate Paleontology,2006,26(1):60-69.
[10] 陈孝红,程龙. 贵州普安中三叠统混鱼龙(鱼龙类:爬行动物)一新种[J]. 古生物学报,2010,34(6):251-260.
[11] JIANG D Y, MAISCH M W, SUN Z Y, et al. A new species ofLariosaurus(Reptilia, Sauropterygia) from the Middle Anisian (Middle Triassic) of southwestern China[J]. Neues Jahrbuch für Geologie und Paläontologie-Abhandlungen,2006,242(1):19-42.
[12] JIANG D Y, MAISCH M W, HAO W C, et al.Nothosaurusyangyuanensissp(Reptlia, Sauropterygia, Nothosauridae) from the middle Anisian (Middle Triassic) of Guizhou, south-western China[J]. Neues Jahrbuch für Geologie und Paläontologie:Monatschefte,2006(5):314-317.
[13] JIANG D Y, RIEPPEL O, MOTANI R, et al. A new Middle Triassic eosauropterygian (Reptilia, Sauropterygia) from Southwestern China[J]. J Vertebrate Paleontology,2008,28(4):1055-1062.
[14] SHANG Q H. A new species ofNothosaurusfrom the early middle Triassic of Guizhou, China[J]. Vertebrata Palasiatica,2006,44(3):237-249.
[15] JIANG D Y, MOTANI R, HAO W C, et al. First record of placodontoidea (Reptilia, Sauropterygia, placodontia) from the eastern tethys[J]. J Vertebrate Paleontology,2008,28(3):904-908.
[16] LI C. First record of protorosaurid reptile (Order Protorosauria) from the middle Triassic of China[J]. Acta Geologica Sinica,2003,77(4):419-424.
[17] LI C, WU X C, CHENG Y N. An unusual archosaurian from the marine Triassic of China[J]. Naturwissenschaften,2006,93:200-206.
[18] ZOU X D, BALINI M, JIANG D Y, et al. Ammonoids from the Zhuganpo member of the Falang formation at Nimaigu and their relevance for dating the Xingyi fossil:lagerstatte(late ladinian, Guizhou, China)[J]. Rivista Italiana di Paleontologia e Stratigrafia,2015,121(2):135-161.
[19] 杨钟健. 贵州新发现的肿肋龙化石胡氏贵州龙[J]. 古脊椎动物学报,1958,2(2/3):315-335.
[20] 邹晓东,郭文,江大勇,等. 贵州省兴义地区中三叠世兴义动物群化石储集层古环境初步分析[J]. 北京大学学报(自然科学版),2015,51(3):472-484.
[21] 杨鹏飞,季承,江大勇. 贵州兴义中三叠世黔鱼龙属(爬行纲:鱼龙目)一新种[J]. 北京大学学报(自然科学版),2013,49(6):1002-1008.
[22] RIEPPEL O, LIU J, LI C. A new species of the thalattosaur genusAnshunsaurus(Reptilia:Thalattosauria) from the Middle Triassic of Guizhou Province, Southwestern China[J]. Vertebrata Pal Asiatica,2006,44(4):285-296.
[23] 程龙,陈孝红,王传尚. 贵州晚三叠世安顺龙(爬行纲:海龙目)一新种[J]. 地质学报,2007,81(10):1345-1351.
[24] 赵丽君,李淳,刘俊,等. 云南中三叠世豆齿龙类(爬行纲:楯齿龙目)一新属[J]. 古脊椎动物学报,2008,46(3):171-177.
[25] CHENG Y N, WU X C, SATO T, et al. A new eosauropterygian (Diapsida, Sauropterygia) from the Triassic of China[J]. J Vertebrate Paleontology,2012,32(6):1335-1349.
[26] 李淳. 贵州中三叠世长颈龙属(原龙目:长颈龙科)一幼年个体[J]. 古脊椎动物学报,2007,45(1):37-42.
[27] 李锦玲,奥利维尔·瑞柏尔. 贵州兴义中三叠世Nothosaurus一新种[J]. 古脊椎动物学报,2004,42(1):1-15.
[28] 刘冠邦,尹恭正,王雪华,等. 记贵州顶效中三叠世一新的海生爬行动物[J]. 高校地质学报,2002,8(2):220-226.
[29] 杨钟健. 中国湖北、贵州的幻龙[J]. 古脊椎动物与古人类,1965,9(4):69-82.
[30] CHENG Y N, SATO T, WU X C, et al. First complete pistosauroid from the Triassic of China[J]. J Vertebrate Paleontology,2006,26(2):501-504.
[31] 李锦玲,刘俊,奥利维尔·瑞柏尔. 贵州兴义中三叠统法郎组竹杆坡段Lariosaurus一新种[J]. 古脊椎动物学报,2002,40(2):114-126.
[32] 李锦玲,金帆. 贵州龙脊椎动物群研究新进展[J]. 自然科学进展,2003,13(8):796-800.
[33] XU G H, ZHAO L J, GAO K Q, et al. A new stem-neopterygian fish from the Middle Triassic of China shows the earliest over-water gliding strategy of the vertebrates[J]. Proceedings of the Royal Society B:Biological Sciences,2013,280(1750):867-907.
[34] 刘冠邦,尹恭正,王雪华. 贵州兴义发现最原始弓鳍鱼[J]. 古生物学报,2002,41(3):461-463.
[35] 孙作玉,郝维城,江大勇. 贵州关岭动物群时代的牙形石地层证据[J]. 北京大学学报(自然科学版),2003,39(1):118-125.
[36] MU A T. On the discovery of the crown of Trumatocrinus[J]. Bulletin of the Geological Society of China,1949,24:85-92.
[37] 李淳. 贵州三叠纪一新鱼龙的初步研究[J]. 科学通报,1999,44(12):1318-1321.
[38] RIEPPEL O, LIU J, BUCHER H. The first record of a thalattosaur reptile from the Late Triassic of South China[J]. J Vertebrate Paleontology,2000,20(3):507-514.
[39] 王尚彦,王敏,刘家仁. 关岭生物群产出地层的岩石微相和环境意义[J]. 贵州地质,2004,22(1):43-45.
[40] 尹恭正,周修高,曹泽田,等. 贵州关岭晚三叠纪早期海生爬行动物的初步研究[J]. 地质地球化学,2000,28(3):1-23.
[41] 陈孝红,程龙. 贵州晚三叠世关岭动物群大型鱼龙化石一新种[J]. 地质通报,2003,2(4):228-235.
[42] 陈孝红,程龙,SANDER P M. 贵州关岭上三叠统卡洛维龙(爬行动物:鱼龙类)一新种[J]. 中国地质,2007,49(2):974-982.
[43] 李淳. 贵州关岭上三叠统的楯齿龙类化石[J]. 古脊椎动物学报,2000,38(4):37-42.
[44] 李淳,RIEPPEL O. 贵州关岭晚三叠世一奇特的楯齿龙类[J]. 科学通报,2002,47(2):156-159.
[45] CHENG Y N, WU X C, SATO T. A new thalattosaurian (Reptilia: Diapsida) from the Upper Triassic of Guizhou, China[J]. Vert Pal Asiat,2007,45(3):246-260.
[46] 赵丽君,刘俊,李淳,等. 贵州关岭海龙类一新属种[J]. 古脊椎动物学报,2013,51(1):24-28.
[47] 王立亭. 半甲齿龟的发现及其意义[J]. 贵州地质,2009,26(2):160-161.
[48] LIU J, RIEPPEL O. The second thalattosaur from the Triassic of Guizhou, China[J]. Vertebrata Pal Asiatica,2001,39(2):77-87.
[49] LI J L. Abstact for the international symposium on the global strstotype of the permian-triassic boundary and the Paleozoic-Mesozoic event, Changxin, China[C]//Triassic marine reotiles from Guizhou of China,2001:59-60.
[50] 喻羑艺,罗永明,尹恭正. 贵州关岭动物群中的海百合[J]. 贵州地质,2000,17(40):40-44.
[51] China State. Chengjiang Fossil Site[EB/OL]. http://whc.unesco.org/en/list/1388,2016-09-20.
[52] Russian Federation State. Lena Pillars Nature Park[EB/OL]. http://whc.unesco.org/en/list/1299,2016-09-20.
[53] Viet Nam State. Phong Nha-Ke Bang National Park[EB/OL]. http://whc.unesco.org/en/list/951,2016-09-20.
[54] Canada State. Miguasha National Park[EB/OL]. http://whc.unesco.org/en/list/686,2016-09-20.
[55] Canada State. Joggins Fossil Cliffs[EB/OL]. http://whc.unesco.org/en/list/1285,2016-09-20.
[56] America State. Grand Canyon National Park[EB/OL]. http://whc.unesco.org/en/list/75,2016-09-20.
[57] Argentina State. Ischigualasto / Talampaya Natural Parks[EB/OL]. http://whc.unesco.org/en/list/966,2016-09-20.
[58] United Kingdom of Great Britain and Northern Ireland State. Dorset and East Devon Coast[EB/OL]. http://whc.unesco.org/en/list/1029,2016-09-20.
[59] Italy and Switzerland State. Monte San Giorgio[EB/OL]. http://whc.unesco.org/en/list/1090,2016-09-20.
[60] Canada State. Dinosaur Provincial Park[EB/OL]. http://whc.unesco.org/en/list/71,2016-09-20.
[61] Denmark State. Stevns Klint[EB/OL]. http://whc.unesco.org/en/list/1416,2016-09-20.
[62] America State. Great Smoky Mountains National Park[EB/OL]. http://whc.unesco.org/en/list/259,2016-09-20.
[63] Egypt State. Wadi Al-Hitan (Whale Valley)[EB/OL]. http://whc.unesco.org/en/list/1186,2016-09-20.
[64] Germany State. Messel Pit Fossil Site[EB/OL]. http://whc.unesco.org/en/list/720,2010.
[65] Australia State. Australian Fossil Mammal Sites(Riversleigh/Naracoorte)[EB/OL]. http://whc.unesco.org/en/list/698,2016-09-20.
[66] Kenya State. Lake Turkana National Parks[EB/OL]. http://whc.unesco.org/en/list/801,2016-09-20.
[67] America State. Yellowstone National Park[EB/OL]. http://whc.unesco.org/en/list/28,2016-09-20.
[68] America State. Petrified Forest National Park[EB/OL]. http://whc.unesco.org/en/tentative lists/5253,2016-09-20.
[69] Argentina State. Sierra de las Quijadas National Park[EB/OL]. http://whc.unesco.org/en/tentative lists/2021,2016-09-20.
[70] 王砚耕,王立亭,王尚彦. 论关岭生物群及其科学意义[J]. 贵州地质,2000,17(3):145-151.
猜你喜欢
学苑创造·B版(2022年8期)2022-06-21
科学大观园(2022年7期)2022-04-22
疯狂英语·新悦读(2021年5期)2021-06-08
华南地质(2020年1期)2020-07-07
——黑龙江省猛犸象动物群之奥妙
化石(2019年1期)2019-05-14
化石(2018年4期)2018-12-19
新生代(2018年14期)2018-11-13
小哥白尼(趣味科学)(2018年7期)2018-09-19
小学科学(2015年12期)2016-01-05
环球时报(2015-05-07)2015-05-07
