
基于Δ13C的不同水氮管理对水稻水分利用效率的影响
2018-05-31 03:16张忠学郑恩楠聂堂哲陈帅宏李伟彤
农业机械学报 2018年5期
张忠学 陈 鹏 郑恩楠 聂堂哲 陈帅宏 李伟彤
(1.东北农业大学水利与土木工程学院, 哈尔滨 150030; 2.农业部农业水资源高效利用重点实验室, 哈尔滨 150030;3.东北农业大学资源与环境学院, 哈尔滨 150030)
0 引言
水分和氮素是调节作物生长发育的主要环境因子,也是决定作物产量的重要影响因素,水分状况会限制肥效的发挥,氮素营养的好坏会通过影响作物叶片光合和水分生理过程最终影响其水分利用效率[1],合理的水氮管理对提高作物的养分及水分利用效率有着显著的协同促进作用,即达到“以肥调水”、“以水调肥”的目的。世界范围内的缺水以及连年的干旱促使节水农业的持续发展,作为节水农业的最终目标,提高作物水分利用效率是实现高效用水增产的核心所在。叶片尺度上水分利用效率的研究可以揭示作物内在的耗水机制,一般表示为瞬时水分利用效率(LWUEins)和内在水分利用效率(LWUEint),LWUEins和LWUEint可以反映作物水分利用过程对环境因素的动态响应[2],但通常LWUEins和LWUEint都是采用气体交换技术(Gas exchange technique)测定某特定时间下作物部分叶片的气体交换参数来确定的,只能代表作物对环境因素的短期响应,在作物的较长生长期内,LWUEins和LWUEint在表达作物对环境因素响应的生理过程具有局限性。
近年来,稳定同位素技术在农业、生态、环境科学等研究领域得到广泛应用[3-7]。研究表明,植物叶片碳同位素比率(δ13C)和碳同位素分辨率(Δ13C)不仅可以反映植物叶片胞间与大气CO2分压比(Pi/Pa)和浓度比(Ci/Ca)[8],还反映了净光合同化速率和气孔导度的相对大小[9]。同时许多相关研究均表明叶片的δ13C和Δ13C可以反映出植物在一段时间内对水分的利用以及对水分胁迫的适应状况,可以用来指示植物长期水分利用效率[10]。因此与气体交换测定相比,叶片δ13C和Δ13C可以更有效地作为不同环境因素下整株植物功能的综合反映指标,还可以用其来评估由气体交换技术确定的LWUEins和LWUEint的准确度。已经有大量研究结果表明,影响植物气体交换代谢过程的环境因子对植物叶片碳同位素分馏程度也产生影响,包括水分条件[11-12]、氮素形态[13]、土壤含水率与大气湿度[14]、降雨量[15]等。但对于不同水氮管理对水稻气体交换参数与碳同位素分辨率的影响系统分析报道较少。
本文以不同水氮管理下种植的水稻作为研究对象,在分析不同水氮管理对水稻叶片气体交换参数的影响基础上,探讨水稻叶片碳同位素分辨率对不同水氮管理的响应情况及其与产量与气体交换参数的关系,并评估采用水稻叶片碳同位素分辨率表征不同水氮管理下水稻不同水平水分利用效率的可靠性和实用性,以揭示不同水氮管理下水稻叶片气体交换代谢调节规律,为叶片尺度的水分高效利用提供理论。
1 材料与方法
1.1 试验区概况

图1 水稻生长期内空气温度和降雨量的日变化Fig.1 Changes in daily air temperature and rainfall during rice growth period
试验在黑龙江省水稻灌溉试验站进行,该站(127°40′45″E、46°57′28″N)位于庆安县和平镇,是典型的寒地黑土分布区。从水稻移栽到成熟,该地区水稻生长期内日气温和降雨量变化如图1所示,多年平均水面蒸发量750 mm,作物水热生长期为156~171 d,全年无霜期128 d。气候特征属寒温带大陆性季风气候。供试土壤为黑土型水稻土,种植水稻20 a以上,土壤耕层厚度11.3 cm,犁底层厚度10.5 cm。在移栽和施肥前,对试验小区0~20 cm土层进行5点对角取样后混合,并对其主要土壤理化性质进行分析,土壤粒径0.02~2.0 mm的颗粒占37.3%、0.002~ 0.02 mm的颗粒占32.3%、粒径小于0.002 mm的颗粒占30.4%,土壤容重1.01 g/cm3,孔隙度61.8%。土壤基本理化性质为:pH值6.45,耕层土壤(0~20 cm)基础肥力(均为质量比)为:有机质41.8 g/kg、全氮15.06 g/kg、全磷15.23 g/kg、全钾20.11 g/kg、碱解氮198.29 mg/kg、有效磷36.22 mg/kg和速效钾112.06 mg/kg。
1.2 试验设计
试验采用灌水方式和施氮量2因素全面试验,试验处理设计详见表1。设置2种灌水方式(表1):控制灌溉(C)、淹水灌溉(F)。水稻控制灌溉模式除水稻返青期田面保持0~30 mm浅薄水层和黄熟期自然落干外,其余各生育阶段均不建立水层,以根层的土壤含水率为控制指标确定灌水时间和灌水定额,灌水上限为土壤饱和含水率,各生育阶段土壤含水率下限分别取饱和含水率的百分比,用TPIME- PICO64/32型土壤水分测定仪于每天07:00和18:00分别测定各试验小区土壤含水率,当土壤含水率接近或低于处理灌水下限时,人工灌水至灌水上限,维持土壤含水率处于相应生育阶段的灌水上限和灌水下限之间,并记录各处理相应灌水量;淹水灌溉处理的试验小区于每天08:00通过预先埋设的竖尺读取田面水层深度,以确定是否需要灌水。施氮量设6个水平(纯氮),即N0(0 kg/hm2)、N1(60 kg/hm2)、N2(85 kg/hm2)、N3(110 kg/hm2)、N4(135 kg/hm2)、N5(160 kg/hm2)。每个处理设3次重复,共36个试验小区,每个小区面积100 m2(10 m×10 m),各小区之间田埂向地下内嵌40 cm深的塑料板,防止各小区间的水氮交换。氮肥按照基肥∶蘖肥∶促花肥∶保花肥比例为4.5∶2∶1.5∶2分施,各处理磷、钾肥用量均一致,施用P2O545 kg/hm2,K2O 80 kg/hm2,磷肥在移栽前一次性施用,钾肥于移栽前和水稻8.5叶龄分2次施用,前后比例为1∶1。试验选用当地的水稻品种“龙庆稻3号”,在充满土壤的育秧盘中将预发芽的种子培育成幼苗,并于2017年5月17日选取长势相同的水稻幼苗进行移栽,株距16.67 cm、行距30 cm,每穴定3株,9月20日收割,生育期为126 d,在水稻各生长阶段及时除草,防治病虫害,以免影响水稻养分吸收。

表1 稻田生育期内各处理土壤水分管理方式Tab.1 Water management of different irrigation management pattern in rice growth stages
注:θs为土壤饱和含水率。
1.3 观测内容与方法
1.3.1土壤含水率
在各试验小区内预先埋设1根长1 m的测管,采用TPIME- PICO64/32型土壤水分测定仪测定各试验小区0~100 cm土层的土壤含水率,每10 cm采集1个数据,间隔3~5 d测定一次,灌溉或降雨后加测。
不同水氮调控下水稻全生育期耗水量采用水量平衡法计算,为
ET=P+I+K+W1-R-D-W2
(1)
式中ET——耗水量,mm
W1、W2——移栽、收获时土壤储水量,mm
P——降雨量,mmI——灌水量,mm
K——地下水补给量,mm
R——地表径流量,mm
D——耕层土壤水渗漏量,mm
试验区地势比较平坦,降水量远不及蒸发量且地下水位的埋深较深,因此R、K的取值为零,水稻全生育期土壤水渗漏量取试验站连续13 a实测渗漏量的平均值[16]。
1.3.2气体交换参数
每个小区随机选取代表性水稻3穴,于水稻各生育期内天气晴朗无云的09:00—11:00,用LI- 6400XT型便携式光合测定仪自带的6400- 02 LED红蓝光源叶室(Li- COR 6400型,USA)测定每小区已选取水稻植株最新完全展开叶片的净光合速率(Pn)、气孔导度(Gs)、细胞间二氧化碳浓度(Ci)和蒸腾速率(Tr)等气体交换参数,每片叶片连续读3次数取平均值。测定时设置叶室内光照强度为1 500 μmol/(m2·s),CO2浓度400 μmol/(m2·s),待叶片适应叶室内环境后进行测定。
1.3.3水分利用效率
作物水分利用效率(WUE)的计算公式为
WUE=Y/ET
(2)
式中Y——水稻产量,kg/hm2
叶片水分利用效率可以表示为叶片的瞬时水分利用效率(LWUEins)和内在水分利用效率(LWUEint)。
叶片尺度的瞬时水分利用效率(LWUEins)用叶片通过蒸腾消耗的一定量的水同化的二氧化碳的量来表示,计算公式为
LWUEins=Pn/Tr
(3)
叶片的内在水分利用效率(LWUEint)用叶片通过蒸腾消耗一定量的水所通化的水量表示,计算公式为
LWUEint=Pn/Gn
(4)
1.3.4碳同位素比率及碳同位素分辨率
于水稻分蘖期、拔节孕穗期和抽穗开花期对不同水氮处理的水稻植株健康叶片进行取样,每个处理3次重复,将水稻叶片在70℃干燥至质量恒定,使用球磨机对水稻叶片样品进行粉碎处理,过80目筛后密封保存,稳定同位素测试在东北农业大学农业部水资源高效利用重点实验室完成,采用元素分析仪(Flash 2000 HT)和同位素质谱仪(DELTA V Advantage,Thermo Fisher Scientific,USA)联用的方法测定水稻叶片稳定同位素组成(δ13C),每隔10个样品加入一个已知δ13C值的标准样品进行测试质量控制,测量误差小于0.2‰,碳同位素值采用国际V- PDB标准,千分比(‰)表示,根据文献[6-7]的公式计算水稻叶片的δ13C,为

(5)
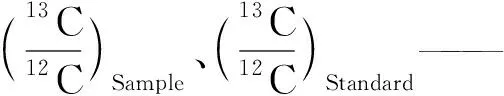
水稻叶片Δ13C值计算公式为

(6)
式中δair——大气CO2的Δ13C值,取-8‰[15]
δplant——测定的水稻各部位样品的Δ13C值
1.3.5产量
齐穗期各处理小区随机调查20穴水稻的有效穗数,并计算平均有效穗数。于成熟期各处理按平均有效穗数选取10穴代表性水稻进行考种,考查穗数、穗粒数、结实率和千粒质量,收获时各试验小区单打单收,并按照稻粒标准含水率14.5%折合计算水稻单位面积产量。
1.4 数据统计分析
采用SPSS 13.0对叶片碳同位素分辨率与水分利用效率进行相关性分析,采用单因素方差分析及Duncan多重比较方法来进行各处理水稻气体交换参数显著差异性分析及均值比较,并采用Origin 9.0软件进行作图。
2 结果与分析
2.1 不同水氮处理对叶片气体交换参数的影响
叶片气孔(Leaf stomatal)是植物体与外界气体交换的通道,植物体光合、呼吸及蒸腾作用均受到气孔导度的影响,一般情况下,气孔部分关闭有利于保护植物过度的水分流失,同时也限制了叶片光合作用,但过低的Gs不利于叶片尺度的水分高效利用[1],因此应综合考虑气孔导度变化对植物叶片光合及蒸腾的影响。由图2(图中不同小写字母表示处理在5%水平上差异显著,ns表示在5%水平上不显著。下同。)可知,不同生育期相同施氮量下淹水灌溉处理下水稻叶片Gs高于控制灌溉下水稻叶片Gs,较高的Gs有利于外界CO2进入叶片胞间,使CO2供应充分,加快光合暗反应速率,相同施氮量下淹水灌溉可以获得较控制灌溉更高的净光合速率(Pn),单位时间内叶片光合消耗CO2的量超过了外界通过气孔进入胞间的CO2的量,因此两种灌溉方式不同施氮水平下Ci随施氮量的增加呈逐渐减少趋势,这与徐俊增等[17]的研究结果相一致。淹水灌溉模式下水稻获得更高Pn的同时也具有较高的Tr,使得大量水分通过蒸腾散失,不利于叶片水分的高效利用。
不同生育期淹水灌溉模式下水稻叶片Gs随施氮量的增加而增大,但增幅相对较小,拔节孕穗期FN5处理水稻的Gs最大,较FN0、FN1、FN2、FN3、FN4处理分别增加了27.63%、6.81%、6.04%、3.56%、2.44%;控制灌溉模式下水稻叶片Gs随施氮量的增加呈先增加后减少趋势,施氮量为135 kg/hm2时叶片Gs最大,较CN0、CN1、CN2、CN3、CN5处理分别增加了37.69%、18.62%、17.99%、7.41%、1.60%,与淹水灌溉下不同施氮水平相比控制灌溉下施氮对Gs的增幅效果较优,,这是由于施氮可以有效降低植物由于受到水分亏缺时合成的脱落酸(ABA)水平,从而降低ABA对叶片气体交换的影响,维持一定的气孔导度[19]。适宜施氮提高一定的气孔导度有利于提高叶片的Pn,拔节孕穗期控制灌溉下与不施氮CN0相比CN1、CN2、CN3、CN4、CN5处理水稻叶片Pn分别提高了16.36%、17.99%、29.97%、41.47%、41.30%;叶片Tr提高了4.67%、6.38%、12.39%、17.24%、17.40%,不同施氮水平较不施氮水稻叶片Pn的增幅大于Tr的增幅,从而相对的提高了叶片尺度的水分利用效率。

图2 不同水氮管理对水稻叶片气体交换参数的影响Fig.2 Effect of different water and nitrogen managements on leaf gas exchange parameters of rice

图3 不同水氮管理对水稻各生育时期耗水量的影响Fig.3 Effect of different water and nitrogen managements on water consumption of rice at different growth stages
2.2 不同水氮处理对水稻耗水量、水分利用效率和产量的影响
如图3所示,不同水氮处理下水稻不同生育期耗水量的变化趋势相同,以水稻抽穗开花期为例,2种灌溉方式下水稻耗水量均随施氮量的增加而增大,相同施氮量下淹水灌溉处理水稻耗水量较稻作控制灌溉模式增加了57.44%、54.61%、52.01%、51.96%、48.33%、45.27%,这是由于传统淹水灌溉田面蒸腾蒸发量较大,而稻作控制灌溉模式在满足水稻生长发育需水量的同时还降低了无效蒸发,从而实现了节水减耗的目标。
如表2所示,不同水氮管理下水稻分蘖期LWUEins变化范围为1.007~1.488 μmol/mmol,LWUEint变化范围为15.682~20.190 μmol/mol,不同生育期两种灌溉方式下水稻的LWUEins和LWUEint随施氮量的增加总体呈抛物线趋势,但因水稻返青期后稻田土壤含水率较高,导致控制灌溉模式土壤含水率未达控制下限,水分亏缺对水稻影响并不明显,故不能完全说明不同水氮处理对水稻叶片水分利用效率的影响。拔节孕穗期、抽穗开花期不同水氮处理LWUEins和LWUEint变化表现一致,拔节孕穗期同一灌水方式下与CN0、FN0相比,CN1、CN2、CN3、CN4、CN5处理水稻的LWUEins分别增加了10.44%、10.91%、15.62%、20.65%、20.33%;LWUEint分别增加了0.23%、1.10%、1.38%、2.74%、4.26%;FN1、FN2、FN3、FN4、FN5处理水稻的LWUEins分别增加了9.94%、11.30%、17.59%、20.65%、14.14%,LWUEint分别增加了3.72%、3.80%、9.18%、13.82%、12.27%。2种灌溉方式下施氮量135 kg/hm2增幅效果高于其他处理,显著提高了水稻LWUEins和LWUEint。不同灌溉方式处理间对比显示,控制灌溉模式水稻LWUEint在抽穗开花期相同施氮量下较淹水灌溉分别高出10.12%、4.44%、3.44%、3.64%、10.82%、8.77%,LWUEins分别高出6.89%、4.48%、4.17%、3.48%、8.00%、0.26%。表明控制灌溉下叶片水分利用效率优于淹水灌溉,有利于叶片水分高效利用。
2种灌溉方式下产量均随施氮量的增加而增大,但控制灌溉下施氮量超过一定阈值产量有下降趋势;WUE随施氮量的变化呈先增加后减少趋势,控制灌溉下施氮量为135 kg/hm2产量最高且对应作物及叶片水平水分利用效率最大,淹水灌溉下施氮量为160 kg/hm2产量最高,但其产量水平的水分利用效率较低,不同灌溉方式处理间对比显示,施氮量为60、85、110、135 kg/hm2下控制灌溉模式水稻产量较淹水灌溉分别高出14.06%、24.14%、22.01%、6.46%;WUE高出76.46%、90.83%、84.58%、59.21%。研究结果表明,控制灌溉下施氮量为135 kg/hm2时产量与作物水分利用效率均达到最优,为寒地黑土水稻节水高产提供理论依据。
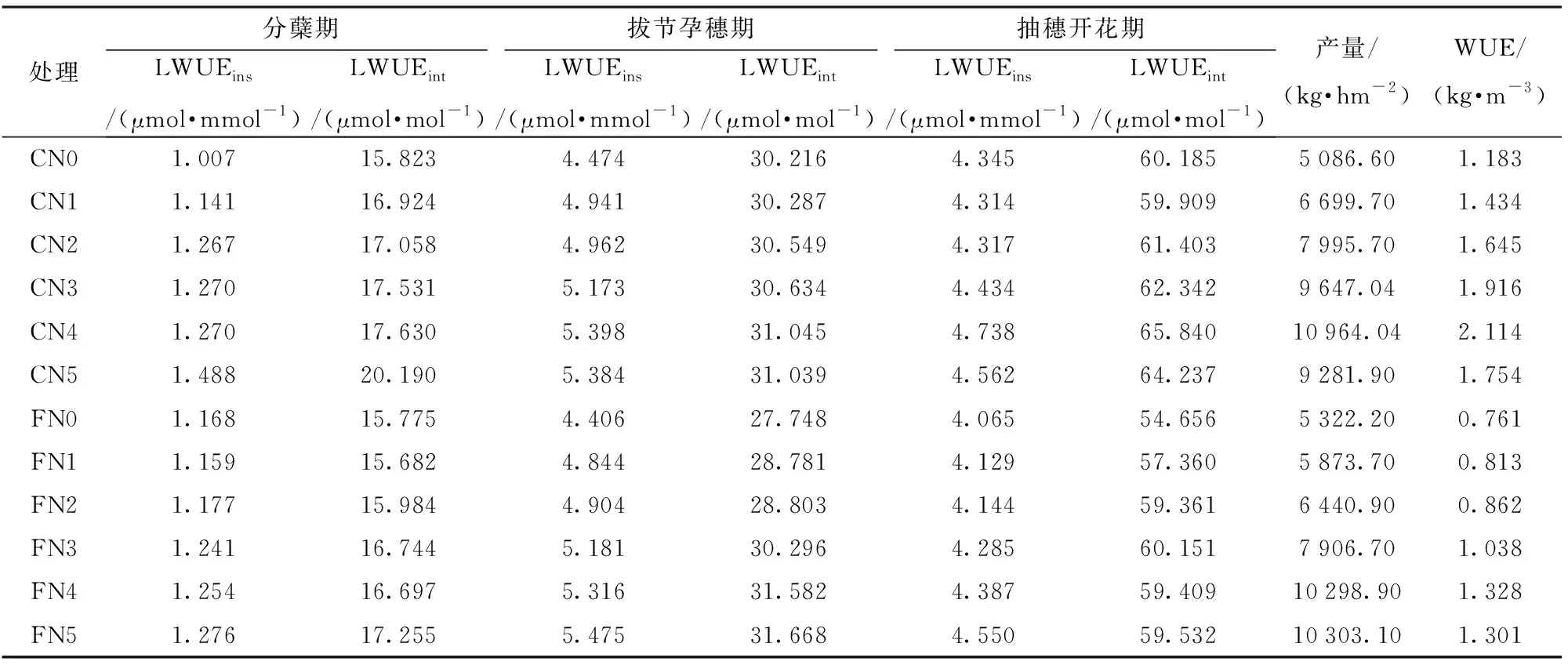
表2 不同水氮管理下水稻叶片LWUEins、LWUEint、WUE和YTab.2 LWUEins, LWUEint, WUE and yield under different water and nitrogen managements
2.3 不同水氮处理对水稻叶片碳同位素分辨率的影响
由表3可知,不同施氮量下控制灌溉模式水稻相同抽穗开花期叶片Δ13C变化范围为20.20‰~20.79‰,淹水灌溉下抽穗开花期叶片Δ13C变化范围为20.42‰~21.03‰,淹水灌溉模式水稻叶片Δ13C在抽穗开花期相同施氮量下较控制灌溉分别高出0.96%、1.45%、1.51%、1.22%、1.68%、0.59%。结果表明,相同施氮量下控制灌溉水稻叶片Δ13C明显小于淹水灌溉下水稻叶片Δ13C,这是由于水稻优先从胞间空气吸收12CO2,使之融于细胞质,使残余的溶解CO2富集13C,且控制灌溉下土壤含水率较低,使水稻叶片气孔导度较淹水灌溉低,不利于外界12CO2进入胞间,从而导致叶片Δ13C较小。
控制灌溉下水稻抽穗开花期叶片Δ13C随施氮量的增加呈先减小后增加的趋势,淹水灌溉下水稻叶片Δ13C随施氮量的增加整体呈逐渐减少趋势,2种灌溉方式下处理CN4和FN5的叶片Δ13C相应为最小,分别为20.20‰、20.42‰。表明在充足的水分条件下,提高施氮水平可以增加光合羧化酶的含量及活性,使溶解在细胞质中的12CO2通过光合羧化酶的作用优先结合到磷酸甘油酸中,生成的糖类物质富集12C而贫化13C,从而使叶片Δ13C减小;当在水分匮缺的情况下,水稻对氮素水平更加敏感,过量的氮素会抑制光合羧化酶的活性,使叶片Δ13C有所增加。

表3 不同水氮管理下水稻各生育期叶片Δ13CTab.3 Leaf Δ13C value at different growth stages under different water and nitrogen managements ‰
2.4 不同水氮管理下水稻叶片Δ13C与气体交换参数、LWUEins、LWUEint、WUE、Y的关系
2.4.1水稻叶片Δ13C与气体交换参数、LWUEins、LWUEint、Y的关系
对抽穗开花期2种灌溉方式下不同氮素水平处理水稻叶片Δ13C和LWUEins、LWUEint的统计分析表明(表4,线性回归方程为Z=ax+y0),2种灌溉方式下不同施氮水平水稻叶片Δ13C与LWUEins、LWUEint均呈负相关,2种灌溉方式下同一施氮水平水稻叶片Δ13C与LWUEint的相关性均明显优于其与LWUEins的相关性。这与WEI等[12]对不同灌溉定额下沟灌番茄的研究结果一致。如表5所示,将同一灌溉方式下不同施氮处理抽穗开花期所有测量数据进行汇总得出,除淹水灌溉Gs外,2种灌溉方式下水稻叶片Δ13C与Pn、Gs、Ci、Tr均呈显著负相关(p<0.05),控制灌溉下水稻叶片Δ13C与LWUEint、Y呈显著负相关(p<0.05),相关系数分别为-0.891、-0.903,同时Δ13C与LWUEins呈负相关,R为-0.749,未达显著水平。淹水灌溉下Δ13C与Y呈显著负相关p<0.05),R为-0.822,Δ13C与LWUEins、LWUEint呈负相关,R为-0.606、-0.779。结果表明,控制灌溉下水稻叶片Δ13C与Pn、Gs、Ci、LWUEins、LWUEint的相关性优于其在淹水灌溉下的相关性,且同一水氮管理下水稻叶片Δ13C对LWUEint的指示性优于对LWUEins的指示性,水稻叶片Δ13C可作为量化表征作物叶片气体交换参数和叶片水平水分利用效率的重要指标。
2.4.2不同生育时期水稻叶片Δ13C与WUE的相关性
如图4所示,不同水氮调控下水稻拔节孕穗期和抽穗开花期的叶片Δ13C均与WUE呈极显著负相关(p<0.01),显著优于分蘖期水稻叶片Δ13C与WUE的相关性;抽穗开花期的水稻叶片Δ13C较拔节孕穗期可以更好地表征水稻WUE。结果表明,通过测定水稻不同时期叶片Δ13C能够为预测不同水氮调控下水稻产量水平水分利用效率提高参考。
3 讨论
水氮管理是调控作物生长和发育的重要途径,通过合理的水氮管理调节作物叶片气体交换能力进而改善叶片光合特性,是实现作物节水高产的重要措施之一[18]。研究证明,施氮可以有效降低植物内源脱落酸(ABA)浓度,对ABA信号运输和传导也有明显影响,从而改善植物叶片气孔开度[19]。本研究中,淹水灌溉下不同生育期水稻叶片Gs随施氮量的增加而增大,但控制灌溉下水稻叶片Gs随施氮量的增加呈抛物线变化,且相同施氮量下控制灌溉的Gs小于淹水灌溉,这可能是由于施氮能提高作物叶片对水分亏缺的敏感性,使得气孔部分关闭,从而减少水分蒸腾。气孔导度的变化直接影响着植物叶片的光合与蒸腾作用,试验结果证明,不同水氮管理下水稻叶片Pn的变化趋势与Gs一致,与庞桂斌等[1]就节水灌溉下水稻光合特性的研究结果不同,原因可能在于本试验中Gs的最大值未超过阈值,故Pn与Gs未呈二次曲线关系。并且水分亏缺会抑制叶片中叶绿素的生物合成,提高叶绿素酶活性并加速叶绿素分解,导致叶片光合能力下降[20]。因此,控制灌溉下水稻叶片Pn小于淹水灌溉。由于施氮使水稻叶片光合能力增强,使单位时间内叶片光合消耗CO2的量超过了外界通过气孔进入胞间的CO2的量,从而导致Ci随施氮量的增加而减小。不同水氮管理主要通过对叶片Gs的调控从而影响叶片Tr。本试验中,拔节孕穗期水稻叶片较高的Gs却具有较低的Tr,这主要是因为拔节孕穗期时当地降雨较多,叶片外部蒸气压较大,而气孔下腔蒸气压相对不变,使得叶片蒸腾变慢。叶片与外界气体与水分的交换情况势必会影响叶片水分利用效率。本研究中,控制灌溉下各施氮水平叶片LWUEins、LWUEint均高于淹水灌溉,LWUEins、LWUEint随施氮量的增加先增大后减少,均在施氮量135 kg/hm2下取得最大值,可见控制灌溉通过较少的水量投入就可获得较优的叶片水分利用效率,这与徐俊增等[17]的研究结果一致。本试验结果证明,通过适量施氮从而使叶片水分利用效率达到最优。

表4 不同水氮管理下抽穗开花期水稻叶片Δ13C与LWUEins、LWUEint之间的相关性与线性回归方程Tab.4 Parameters of liner regression between Δ13C andLWUEins and LWUEint in leaf of rice at heading stageunder different water and nitrogen managements
注:*表示变量之间在p<0.05水平差异显著,下同。

表5 不同灌溉方式下水稻叶片Δ13C与叶片气体交换参数、水分利用效率及产量的相关关系Tab.5 Correlation between Δ13C and gas exchange parameters, water use efficiency and yield
注:** 表示变量之间在p<0.01水平差异显著。

图4 水稻不同生育时期叶片Δ13C与WUE的相关性Fig.4 Correlation between Δ13C and WUE in different growth stages
陆地植物固定大气CO2的过程中会发生碳同位素效应,其中植物的光合作用是产生碳同位素动力学分馏的最主要过程,不同光合途径(C3、C4、CAM)因其光合羧化酶(RuBP酶和PEP酶)和羧化的时空差异对13C有不同的判别和排斥,从而导致不同光合途径的植物具有明显不同的Δ13C,YASIR等[21]对中国49种基因型C3作物小麦品种研究发现,叶片Δ13C与Gs、Ci等气体交换参数呈显著负相关,本研究发现,不同水氮管理下水稻叶片Δ13C与Pn、Gs、Ci同样均呈负相关,这与ZHANG等[22]对于不同施氮水平下冬小麦的研究结果一致。EHLERINGER[23]发现C3作物菜豆叶片Δ13C与LWUEint呈负相关,同时还可对LWUEins起指示作用,本研究也表明,水稻作为C3作物在不同水氮管理下叶片Δ13C与LWUEint、LWUEins均呈负相关,这与CUI等[24]对梨枣树、ZHAO等[25]对旱稻的研究结果一致,本研究还发现,不同水氮管理下水稻叶片Δ13C与LWUEint的相关性明显优于其与LWUEins的相关性,这是由于LWUEint能够更直接地反映叶片气孔的开闭程度,影响大气CO2经气孔向叶内扩散的过程,进而反映在叶片Δ13C的变化上。并且在控制灌溉下能更好地体现叶片Δ13C对LWUEint的指示性,这印证了在土壤水分较低条件下,很多作物的Δ13C可以作为叶片水分利用效率的可靠量化表征指标,这与番茄[4]、硬质小麦[26]的研究结果一致。WUE作为反映作物生产性能的重要指标,已有对多种作物的研究均表明,利用作物不同部位的Δ13C可以有效地指示WUE[27-28],本研究中发现水稻不同生育时期叶片Δ13C与WUE呈负相关,且抽穗开花期水稻叶片Δ13C对WUE的指示性更优,可为估算不同水氮调控下水稻WUE提供参考。
植物组织Δ13C与产量(Y)的关系受到调控作物生理过程的环境因子的影响,可能呈现不同的相关关系,本研究中不同水氮管理下水稻叶片Δ13C与Y呈负相关,这与WEI等[12]的研究结果不完全一致。本试验仅研究了不同水氮调控下水稻叶片Δ13C与LWUEint、LWUEins、WUE和Y的相关性,而作物在不同环境因素下叶片Δ13C对作物不同水平的水分利用效率的表征作用是否稳定还存在争议,尚需进一步验证。
4 结论
(1)稻作控制灌溉模式水稻叶片Gs随施氮量的增加呈先增大后减少趋势。在施氮量为135 kg/hm2下水稻叶片Gs最大,较其余施氮水平处理水稻叶片Gs提高了1.6%~16.7%,适量施氮可以增强水分亏缺条件下水稻叶片的气体交换能力,有利于外界CO2通过气孔进入叶片内部,并扩散到相应的光合作用位点,从而提高了水稻叶片的Pn。淹水灌溉下不同施氮量水稻叶片具有较高的Gs,导致各生育期相同施氮量下水稻叶片Pn均高于控制灌溉,但同时具有较大的Tr,不利于叶片水分的高效利用。
(2)不同水氮管理下拔节孕穗期、抽穗开花期水稻叶片LWUEins、LWUEint及WUE随施氮量的增加呈抛物线变化,适量施氮可以提高水稻叶片LWUEins、LWUEint和WUE。控制灌溉下不同生育期各施氮水平水稻叶片LWUEins、LWUEint和WUE均高于淹水灌溉,本试验中稻作控制灌溉模式施氮量为135 kg/hm2时LWUEins、LWUEint和WUE值最大且产量最高,表明在控制灌溉下适量施氮可以起到节水高产的作用。
(3)控制灌溉下水稻叶片Δ13C随施氮量的增加呈先减小后增加的趋势,淹水灌溉下水稻叶片Δ13C随施氮量的增加整体呈逐渐减少趋势,不同生育期相同施氮量下控制灌溉水稻叶片Δ13C要明显小于淹水灌溉下水稻叶片Δ13C,且不同水氮管理下水稻叶片Δ13C与叶片水平瞬时水分利用效率、叶片水平内在水分利用效率分别呈负相关,拔节孕穗期和抽穗开花期水稻叶片Δ13C与WUE具有较优的负相关性。通过测定水稻不同生育时期叶片Δ13C能够为预测东北寒地黑土区不同水氮管理下水稻不同水平水分利用效率和产量提供参考。
1 庞桂斌, 徐征和, 杨士红,等. 控制灌溉水稻叶片水分利用效率影响因素分析[J/OL]. 农业机械学报, 2017, 48(4):233-241.http:∥www.j-csam.org/jcsam/ch/reader/view_abstract.aspx?flag=1&file_no=20170430&journal_id=jcsam.DOI:10.6041/j.issn.1000-1298.2017.04.030.
PANG Guibin, XU Zhenghe, YANG Shihong, et al. Influence factors analysis of rice leaf water use efficiency under controlled irrigation[J/OL]. Transactions of the Chinese Society for Agricultural Machinery, 2017, 48(4):233-241. (in Chinese)
2 PASCUAL M, LORDAN J, VILLAR J M, et al. Stable carbon and nitrogen isotope ratios as indicators of water status and nitrogen effects on peach trees[J]. Scientia Horticulturae, 2013, 157(3):99-107.
3 王振昌, 郭相平, 黄双双,等. 水稻天然稳定碳、氮同位素组成与产量及品质的关系[J/OL]. 农业机械学报, 2016, 47(12):160-171.http:∥www.j-csam.org/jcsam/ch/reader/view_abstract.aspx?flag=1&file_no=20161220&journal_id=jcsam.DOI:10.6041/j.issn.1000-1298.2016.12.020.
WANG Zhenchang, GUO Xiangping, HUANG Shuangshuang,et al. Relationships between stable C and N isotope compositions of rice plants, rice yield and quality[J/OL]. Transactions of the Chinese Society for Agricultural Machinery, 2016, 47(12):160-171.(in Chinese)
4 魏镇华, 杜太生, 张娟,等. 利用碳同位素分辨率表征沟灌番茄水分利用效率[J]. 农业工程学报, 2013, 29(7):58-65.
WEI Zhenhua, DU Taisheng, ZHANG Juan,et al. Stable carbon isotope discrimination on water use efficiency of field tomato under furrow irrigation[J]. Transactions of the CSAE, 2013, 29(7):58-65.(in Chinese)
5 刘慧, 王为木, 谈建康,等. 稻田水体碳氮稳定同位素组成与时空变化研究[J/OL]. 农业机械学报, 2015, 46(2):132-136.http:∥www.j-csam.org/jcsam/ch/reader/view_abstract.aspx?flag=1&file_no=20150220&journal_id=jcsam. DOI:10.6041/j.issn.1000-1298.2015.02.020.
LIU Hui, WANG Weimu, TAN Jiankang,et al. Temporal and spatial variation in stable carbon and nitrogen isotope in paddy field water[J/OL]. Transactions of the Chinese Society for Agricultural Machinery,2015, 46(2):132-136.(in Chinese)
6 FARQUHAR G D, O’LEARY M H, BERRY J A. On the relationship between carbon isotope discrimination and the intercellular carbon dioxide concentration in leaves[J]. Australian Journal of Plant Physiology,1982,9(2):121-137.
7 郑利剑, 马娟娟, 郭飞,等. 基于水稳定同位素技术的生物炭对土壤持水性影响分析[J/OL]. 农业机械学报, 2016, 47(6):193-198.http:∥www.j-csam.org/jcsam/ch/reader/view_abstract.aspx?flag=1&file_no=20160625&journal_id=jcsam. DOI:10.6041/j.issn.1000-1298.2016.06.025.
ZHENG Lijian, MA Juanjuan, GUO Fei,et al. Impact of biochar addition on soil water retention based on water stable isotopes[J/OL]. Transactions of the Chinese Society for Agricultural Machinery, 2016, 47(6):193-198.(in Chinese)
8 FARQUHAR G D, EHLERINGER J R, HUBICK K T. Carbon isotope discrimination and photosynthesis[J]. Annual Review of Plant Physiology & Plant Molecular Biology, 1989, 40:503-537.
9 MONNEVEUX P, REKIKA D, ACEVEDO E, et al. Effect of drought on leaf gas exchange, carbon isotope discrimination, transpiration efficiency and productivity in field grown durum wheat genotypes[J]. Plant Science, 2006, 170(4):867-872.
10 SULLIVAN P F, WELKER J M. Variation in leaf physiology of Salix arctica, within and across ecosystems in the high arctic: test of a dual isotope (Δ13C and Δ18O) conceptual model[J]. Oecologia, 2007, 151(3):372-386.
11 CUI N, DU T, KANG S, et al. Relationship between stable carbon isotope discrimination and water use efficiency under regulated deficit irrigation of pear-jujube tree[J]. Agricultural Water Management, 2009, 96(11):1615-1622.
12 WEI Z, DU T, ZHANG J, et al. Carbon isotope discrimination shows a higher water use efficiency under alternate partial root-zone irrigation of field-grown tomato[J]. Agricultural Water Management, 2016, 165:33-43.
13 MARTAS L, JOSEÉL A. Nitrogen source and water regime effects on durum wheat photosynthesis and stable carbon and nitrogen isotope composition.[J]. Physiologia Plantarum, 2006, 126(3):435-445.
15 ANDERSON J E, KRIEDEMANN P E, AUSTIN M P, et al. Eucalypts forming a canopy functional type in dry sclerophyll forests respond differentially to environment[J]. Australian Journal of Botany, 2000, 48(6):759-775.
16 郭龙珠. 水稻旱育稀植栽培模式下的优化灌溉制度研究[D].哈尔滨:东北农业大学, 2002.
GUO Longzhu. Optimized irrigation regime of rice under dry seeded and thinly populated cultivated pattern[D]. Harbin: Northeast Agricultural University, 2002.(in Chinese)
17 徐俊增, 彭世彰, 魏征,等. 节水灌溉水稻叶片胞间CO2浓度及气孔与非气孔限制[J]. 农业工程学报, 2010, 26(7):76-80.
XU Junzeng, PENG Shizhang, WEI Zheng,et al. Intercellular CO2concentration and stomatal or non-stomatal limitation of rice under water saving irrigation[J]. Transactions of the CSAE, 2010, 26(7):76-80.(in Chinese)
18 张彦群, 王建东, 龚时宏,等. 滴灌条件下冬小麦施氮增产的光合生理响应[J]. 农业工程学报, 2015, 31(6):170-177.
ZHANG Yanqun, WANG Jiandong, GONG Shihong,et al. Photosynthetic response of yield enhancement by nitrogen fertilization in winter wheat fields with drip irrigation[J]. Transactions of the CSAE, 2015, 31(6):170-177.(in Chinese)
19 CHEN Y, MURCHIE E H, HUBBART S, et al. Effects of season-dependent irradiance levels and nitrogen-deficiency on photosynthesis and photoinhibition in field-grown rice (Oryzasativa).[J]. Physiologia Plantarum, 2003, 117(3):343-351.
20 刘小刚,耿宏焯,程金焕,等. 保水剂和灌水对小粒咖啡苗木的节水调控效应[J/OL]. 农业机械学报, 2014, 45(3):134-139.http:∥www.j-csam.org/jcsam/ch/reader/view_abstract.aspx?flag=1&file_no=20140323&journal_id=jcsam.DOI:10.6041/j.issn.1000-1298.2014.03.023.
LIU Xiaogang, GENG Hongzhuo, CHENG Jinhuan, et al. Effect of super absorbent polymer and irrigation on improving water use efficiency of Arabica coffee seedling[J/OL]. Transactions of the Chinese Society for Agricultural Machinery, 2014, 45(3):134-139.(in Chinese)
21 YASIR T A, MIN D, CHEN X, et al. The association of carbon isotope discrimination (Δ) with gas exchange parameters and yield traits in Chinese bread wheat cultivars under two water regimes[J]. Agricultural Water Management, 2013, 119:111-120.
22 ZHANG Y, WANG J, GONG S, et al. Nitrogen fertigation effect on photosynthesis, grain yield and water use efficiency of winter wheat[J]. Agricultural Water Management, 2017, 179:277-287.
23 EHLERINGER J R. Correlations between carbon isotope discrimination and leaf conductance to water vapor in common beans[J]. Plant Physiology, 1990, 93(4):1422-1425.
24 CUI N B, DU T S, KANG S Z, et al. Relationship between stable carbon isotope discrimination and water use efficiency under regulated deficit irrigation of pear-jujube tree[J]. Agricultural Water Management, 2009, 96(11):1615-1622.
25 ZHAO B, KONDO M, MAEDA M, et al. Water-use efficiency and carbon isotope discrimination in two cultivars of upland rice during different developmental stages under three water regimes[J]. Plant & Soil, 2004, 261(1-2):61-75.
26 CABRERABOSQUET L, MOLERO G, BORT J, et al. The combined effect of constant water deficit and nitrogen supply on WUE, NUE and Δ13C in durum wheat potted plants[J]. Annals of Applied Biology, 2007, 151(3):277-289.
27 CHUN Yingyin, XIANG Wang, BAO Liduan, et al. Early growth, dry matter allocation and water use efficiency of two sympatric Populus, species as affected by water stress[J]. Environmental and Experimental Botany, 2005, 53(3):315-322.
28 RYTTER R M. Water use efficiency, carbon isotope discrimination and biomass production of two sugar beet varieties under well-watered and dry conditions[J]. Journal of Agronomy & Crop Science, 2005, 191(6):426-438.
猜你喜欢
作物学报(2022年2期)2022-11-06
灌溉排水学报(2022年9期)2022-09-28
中国农业科学(2022年17期)2022-09-19
作物学报(2022年9期)2022-07-18
中国土壤与肥料(2021年5期)2021-12-02
养殖与饲料(2021年4期)2021-04-17
河南农业·综合版(2020年10期)2020-10-20
河南农业(2020年10期)2020-10-15
安徽农学通报(2020年3期)2020-02-28
节水灌溉(2018年3期)2018-04-12
