
HIV相关脑病患者中枢神经系统来源的HIV-1 Tat蛋白对HUVECs活性的影响
2018-05-25 08:13郑文慧秦泽明李昕曹馨月单晓宇温红玲王志玉黄涛赵丽
中华实验和临床病毒学杂志 2018年6期
郑文慧 秦泽明 李昕 曹馨月 单晓宇 温红玲 王志玉 黄涛 赵丽
250012济南,山东大学公共卫生学院卫生微生物检验学系山东省“十三五”高校重点实验室 感染性疾病防控重点实验室(郑文慧、李昕、曹馨月、单晓宇、温红玲、王志玉、赵丽);250012济南,山东大学公共卫生学院预防医学实验中心(秦泽明);250014济南,山东省疾病预防控制中心(黄涛)
HIV相关脑病(HIV-associated dementia,HAD)是艾滋病患者晚期出现的神经系统并发症,患者血脑屏障(blood brain barrier,BBB)出现不同程度的结构损伤和功能减退,但其机制尚未阐释清楚。HIV-1 Tat是HIV-1基因编码的重要调控蛋白,可借助被HIV-1感染的巨噬细胞等多种细胞分泌、释放到细胞外,借助自身的穿膜功能到达机体多个部位。内皮细胞是组成BBB的重要细胞,其活性对BBB功能尤为重要,本研究表达并纯化了HAD患者大脑基底核来源的Tat蛋白,研究了其对人脐静脉内皮细胞(human umbilical vein endothelial cells,HUVECs)活性的影响,以期为HIV-1神经致病机制提供科学依据。
1 材料与方法
HAD患者基底核(basal ganglia,BG)来源的HIV-1基因(pGEX-KG-tat,216 bp)由本室克隆并保存。镍亲和层析柱、凝胶过滤预装柱(Column Superdex 75 Increase 10/300 GL)和蛋白纯化仪(AKTA explorer)为GE公司产品,His-Tag抗体为proteintech公司产品,BCA蛋白浓度测定试剂盒为碧云天生物技术有限公司产品,cck-8为biosharp公司产品,酶标仪(Synergy2)为美国BioTek公司产品,HUVECs为本室保存。

PCR产物纯化后和pET-32a载体酶切用T4连接酶22℃连接过夜。将连接产物转化大肠杆菌DH5-α,用氨苄青霉素筛选,阳性菌落送上海生工生物工程有限公司测序。
1.2.2 Tat蛋白的原核表达及鉴定:构建成功的重组质粒pET-32a-tat转化BL21(DE3),挑取单菌落接种到含氨苄青霉素的LB培养基中,37℃震荡培养至吸光度值在0.6~0.8之间,用IPTG 37℃进行诱导。菌液用PBS清洗,3次后用Tris-NaCl(Tris浓度为50 mmol/L,NaCl浓度为300 mmol/L,PH=8.0)重悬,冰上超声破碎并离心,上清和沉淀经Western blot(WB)鉴定,同时设pET-32a质粒对照和细菌对照。
1.2.3 Tat蛋白的纯化、鉴定及定量:冰上超声破碎,4℃,9 000×离心30 min收集上清,蛋白纯化仪进行纯化。按照产品说明书处理镍亲和层析柱后上样,用含 10 mmol/L、200 mmol/L、500 mmol/L 咪唑的Tris-NaCl溶液洗脱,根据吸收峰的位置,收集洗脱液并经SDS-PAGE和WB鉴定。蛋白纯度高的洗脱液经超滤离心管浓缩,再经Column Superdex 75 Increase 10/300 GL进一步纯化,洗脱液为含1 mmol/L EDTA的Tris-NaCl,SDS-PAGE和WB进行鉴定,浓缩后BCA试剂盒进行蛋白定量。
1.2.4 Tat蛋白对内皮细胞活性的影响:HUVECs常规培养,待细胞铺满至90%时,将Tat蛋白加入96 孔板中,浓度依次为 100 ng/ml、200 ng/ml、300 ng/ml、400 ng/ml、500 ng/ml、1 000 ng/ml,每个浓度设5个平行孔,以正常细胞为阴性对照,培养基作为空白对照。36 h后,按cck-8产品说明书操作检测细胞活性,酶标仪450 nm检测吸光度()。
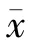
2 结果
PCR扩增产物经琼脂糖凝胶电泳,在250 bp处出现与预计的第一外显子基因(216 bp)片段大小相符的条带。重组质粒pET-32a-tat转化后的阳性菌经PCR鉴定和测序证实为HIV-1基因。
WB结果显示,重组质粒 pET-32a-tat转化 BL21(DE3)后,经IPTG诱导后在相对分子质量15×10处有一条明显蛋白条带(见图1箭头所示),与目的蛋白大小一致,还有小部分蛋白以多聚体存在,大部分可溶性蛋白在上清中,还有较少部分在包涵体内。上清经镍亲和层析柱纯化,500 mmol/L洗脱液中Tat蛋白含量较高,加入DTT后对蛋白多聚体影响较小(见图2所示)。该峰洗脱液浓缩后经Column Superdex 75 Increase 10/300 GL进一步纯化可见单个吸收峰,根据出峰位置可知为一分子量较大的多聚体,纯化浓缩后的Tat蛋白在DTT作用下,分解成以单体为主的多种形式(见图3)。该单峰洗脱液浓缩,测得浓度为0.47 mg/ml。
HUVECs吸光度检测结果及统计学分析见表1和表2。结果显示,不同浓度Tat蛋白的实验组中,HUVECs细胞的均数在组间的差异具有统计学意义(<0.05);阴性对照组(0 ng/ml)、100 ng/ml和200 ng/ml实验组之间均数的差异无统计学意义(>0.05);与阴性对照组相比,300 ng/ml、400 ng/ml、500 ng/ml和1 000 ng/ml组显著降低,差异有统计学意义(<0.05),300 ng/ml、400 ng/ml、500 ng/ml和1 000 ng/ml组之间值的差异无统计学意义(=19.607,=0.000)。随着Tat蛋白浓度的增加,细胞活性逐渐降低,300 ng/ml组细胞活性下降更明显(图4)。

图1 HIV-1 Tat的表达与鉴定A: SDS-PAGE;B:Western blotM:Protein marker;1:Bacterium control;2:pET-32a plasmid control;3:pET-32a-tat positive bacterium;4:Supernatant;5:PrecipitationFig.1 Expression and identification of HIV-1 Tat
3 讨论

图2 HIV-1 Tat经镍亲和层析柱纯化和鉴定A:UV absorption of AKTA explorer;B:SDS-PAGE;C:Western blotM:protein marker;1:Waste liquor;2:Waste liquor with DTT;3:200 mmol/L imidazole eluent;4:200 mmol/L imidazole eluent with DTT;5:500 mmol/L imidazole eluent;6:500 mmol/L imidazole eluent with DTTFig.2 Purification of HIV-1 Tat protein by Ni-chelating chromatography column and identification

表1 不同浓度HUVECs细胞吸光度检测结果Tab.1 Absorbance of HUVECs in different concentration groups

表2 不同组HUVECs吸光度多重比较方法的结果Tab.2 Absorbance of HUVECs in different groups results of multiple comparisons
Tat蛋白由调控基因tat的两个独立外显子编码,可划分为N末端激活区、半胱氨酸富集区、核心区、碱性氨基酸富集区、谷氨酰胺富集区、C-末端区六个功能区。Tat蛋白对BBB相关细胞的作用方式可分为两种:①直接作用:Tat可与细胞表面受体结合,发挥生物学作用,如碱性氨基酸富集区可介导Tat蛋白与硫酸类肝素蛋白多糖(heparan sulfate proteoglycans,HSPG)结合,使Tat蛋白在细胞外基质富集,Tat蛋白与碱性成纤维细胞生长因子(basic fibroblast growth factor,bFGF)竞争结合 HSPG,使bFGF浓度增加,从而促进卡波氏肉瘤和内皮细胞增长;也可借助碱性氨基酸富集区的穿膜特性进入细胞,可调控HIV-1基因组表达,并为病毒复制创造适宜环境。还可通过损伤细胞器、上调促凋亡蛋白表达等途径损伤细胞,影响细胞发挥正常生理作用。②间接作用:通过激活信号通路等途径,诱导星形胶质细胞产生ROS和促炎症介质,参与炎症反应,进而损伤BBB。近些年的研究表明,HIV-1 Tat蛋白可通过损伤BBB相关细胞、调节紧密连接蛋白的表达等途径损伤BBB的结构,进而影响BBB发挥正常功能。

A:AKTA explorer UV 吸收图;B:SDS-PAGE;C:Western blot;图3 Column Superdex 75 Increase 10/300 GL纯化的HIV-1 Tat及鉴定A:UV absorption of AKTA explorer;B:SDS-PAGE;C:Western blot;Fig.3 Purification of HIV-1 Tat by Column Superdex 75 Increase 10/300 GL and determination

∗与阴性对照组比较,P<0.05图4 HIV-1 Tat蛋白对HUVECs细胞活性的影响∗P<0.05 VS negative controlFig.4 The effects of HIV-1 Tat on viability of HUVECs
本课题组前期研究发现,该HAD患者感染的HIV-1为B亚型,其BG来源tat序列与HXB2 tat标准序列间的基因距离差异有统计学意义(<0.05),而非HAD患者BG来源tat序列与HXB2 tat标准序列间的基因距离差异无统计学意义(>0.05),HAD患者BG来源的Tat诱导U87细胞分泌的TNF-α和IL-1β比非HAD患者BG来源组高,表明二者生物学活性存在差异。本研究表达了HAD患者BG来源的HIV-1 Tat,并进行纯化,研究发现,经镍亲和层析柱纯化的Tat蛋白在浓度较低时主要是以单体和二聚体存在,蛋白浓缩后经凝胶过滤预装柱进一步纯化后,得到的UV吸收峰为单峰。在DTT的作用下,多聚体逐渐分解成以单体为主、多种聚合体共同存在,说明Tat多聚体的形成并不完全依靠二硫键,其他机制有待进一步研究。
HIV-1进入中枢神经系统首先要通过血脑屏障,血脑屏障维持脑内各种离子、递质等的动态平衡与BBB的细胞和结构密不可分。血管内皮细胞是血脑屏障最表面的细胞,对维持血脑屏障的通透性具有非常重要的作用,同时,内皮细胞之间形成的紧密连接还可控制细胞旁路的物质转运。HIV-1感染细胞后,在细胞内增殖,合成病毒蛋白,通过直接感染内皮细胞和破坏紧密连接蛋白损伤血脑屏障。血脑屏障的通透性增强,感染了HIV-1的单核细胞等有害物质可进入中枢神经系统,损伤神经元,进而引起一系列的神经行为障碍。基于内皮细胞在HIV入侵中枢神经系统和HIV神经致病性过程中的重要地位,本文使用内皮细胞进行研究。先前研究血管内皮细胞通常采用原代HUVECs,但其增殖能力低且生命周期有限,本研究采用的HUVECs可稳定传代,生长良好,易于培养,且保留了原代内皮细胞的特征,可较好的替代原代内皮细胞进行研究。不同浓度的Tat蛋白对HUVECs活性的影响结果显示,与阴性对照组相比,100 ng/ml和200 ng/ml Tat使HUVECs细胞活性降低,但是差异无统计学意义;随着Tat蛋白浓度的增加,细胞活性逐渐降低,300 ng/ml组细胞活性下降明显。与阴性对照组相比,300 ng/ml、400 ng/ml、500 ng/ml和 1 000 ng/ml四组细胞活性差异均有统计学意义,但是组间差异无统计学意义。不同Tat浓度对HBMECs活性影响的结果可能与 Tat蛋白的作用方式有关,Tat对HUVECs的作用是通过与其表面受体的结合还是进入细胞发挥作用有待于进一步研究。Ma等报道,200 ng/ml Tat蛋白可使人脑微血管内皮细胞(human brain microvascular endothelial cells,HBMECs)活性显著降低,与本研究的结果略有不同,原因之一可能是HUVECs与HBMECs都有内皮细胞的共性,但在结构和功能仍存在差异,使得不同的内皮细胞对外源性物质的敏感性存在差异。此外,不同亚型的HIV-1 Tat蛋白在与 LTR(long terminal repeat sequence,LTR)结合能力、促进病毒增殖和上调细胞因子、趋化因子辅助受体表达等生物学活性方面存在差异,Tat蛋白的氨基酸位点的变异使其功能和致病机制发生转变,不同来源的Tat,其氨基酸位点变异不同,这也可能是造成研究结果差异的原因。
本研究优化了Tat蛋白的纯化条件、且纯化的蛋白具有一定的生物学活性,为进一步研究细胞水平和动物水平Tat蛋白的神经致病机制奠定了基础。
无
猜你喜欢
体育科技文献通报(2022年4期)2022-10-21
河北科技师范学院学报(2022年2期)2022-08-26
世界科学技术-中医药现代化(2022年3期)2022-08-22
世界科学技术-中医药现代化(2021年8期)2021-12-21
载人航天(2021年5期)2021-11-20
现代临床医学(2021年5期)2021-11-02
中成药(2019年12期)2020-01-04
中国眼镜科技杂志(2019年9期)2019-11-11
中成药(2017年8期)2017-11-22
CHINESE JOURNAL OF AERONAUTICS(2016年3期)2016-11-23
