
国家标本资源共享平台兰科植物标本记录采集地理偏差及其环境因子解释
2018-05-20 06:56尹朝露刘雨晴肖翠
数据与计算发展前沿 2018年5期
尹朝露,刘雨晴,肖翠
1.安徽农业大学,资源与环境学院,安徽 合肥,230031
2.中国科学院植物研究所,北京 100093
引言
兰科植物 (Orchidaceae) 有 800 属,约 25,000~ 30,000 种,在全球广泛分布。兰科植物多为地生、附生或较少为腐生草本,极罕为攀援藤本,该类群集中分布在湿润的热带和亚热带地区。我国有兰科植物 194 属 (11 个特有属,1 个引入属) 1,388 种 (491 个特有种,1 个引入种)[1]。分子生物学证据表明,兰科植物为单系类群,起源于古冈瓦纳大陆,由于其在植物系统演化上的特殊地位,成为进化生物学研究的热点类群之一[2]。兰科植物也是保护生物学领域的“旗舰”类群,本类群中的物种多为珍稀濒危植物,全世界所有的野生兰科植物均被列为《野生动植物濒危物种国际贸易公约》(CITES) 的保护范围,占该公约保护植物的 90% 以上[3],其中 651 种兰科植物被列入中国高等植物受威胁物种名录[4]。兰科植物在我国热带和亚热带地区分布广泛,基于物种分布记录,在区域或全国尺度已经有相关的研究对兰科植物的地理分布格局与保护进行了相关的研究。黄启堂等 (2009) 分析了福建省兰科植物的区系组成、生活型、分布区类型及其与邻近地区的关系,为福建省兰科植物种质资源的科学保护和合理开发提供依据[5];张晓龙 (2014) 分析探讨了我国野生兰科植物地理分布格局,并结合环境因子和人文因子对该格局形成的主要驱动力进行了分析,结果表明:在 100km×100km 尺度上,中国野生兰科植物物种丰富度格局和被子植物总体物种丰富度分布格局基本一致,呈现出明显的纬度梯度格局[6]。秦卫华等 (2012) 分析了中国 543 个自然保护区内兰科植物物种名录,对中国 1,334 种兰科植物就地保护状况进行了评价[7]。兰科植物标本记录在生物地理学特别是保护生物学中具有重要地位。系统评估兰科植物标本记录的采集完整性,避免出现由采集不完整导致的地理偏差,对制定兰科植物的保育策略、物种濒危等级的评估以及对兰科植物的空间分布格局研究都有着重要意义。标本采集偏差指在植物标本采集过程中受地理条件,天气,交通,采集人的主观意识等条件的影响导致采集时采集地区分布不均匀,部分地区采集遗漏,或采集数量过少导致数据不完整。加强数据质量评估是提高统计数据质量的有效手段[8]。
国家标本资源共享平台 (简称 NSII,http://www.nsii.org.cn/) 是国家科技部认定并资助的国家科技基础条件平台之一,其下属包括植物、动物、教学、自然保护区、岩矿化石、极地等六个标本子平台;汇集了植物、动物、岩矿化石和极地资源等标本的记录、照片、名录、文献等资源。NSII 的核心和重点是标本数字化,在近十多年的建设和积累中, NSII 成为拥有 1400 多万份数字化标本的大数据平台,其中植物标本已超过 1000 万份。植物标本在进行数字化的同时,标本信息包括采集信息 (采集人、采集时间、采集地、采集号、经纬度坐标)、分类信息 (科属种的学名、标本状态:花果)、馆藏信息 (条形码、存放的标本馆) 等均整合在数据库中,特别是学名和分布地字段的规范和完善,为分类学和生态学的研究提供了坚实的数据基础。将标本数据地标化后,为一些生态学的关键问题的解决起到了很好的助推作用。
本研究拟以国家标本资源共享平台收录的中国兰科植物的植物标本采集记录为研究对象,地理单元选取县级行政单位进行如下分析:(1) 兰科植物标本记录质量评估,包括:标本记录数量,物种丰富度,物种丰富度估计值,物种采集完整性评估。(2) 利用与采集地点相对应的环境因子对物种丰富度格局和标本采集完整性进行解释。相关结果对指导野外标本的采集、生物多样性空间格局研究以及兰科植物保护策略制定等领域有着重要的借鉴意义。
1 材料与方法
1.1 数字化标本记录的获取与筛选
本研究所用的数字化标本记录全部来源于国家标本资源共享平台 (National Specimen Information Infrastructure,NSII)。经过初步筛选,共获取到兰科植物标本采集记录 71,826 份。由于植物标本的采集经历了很长的历史时期,部分数据年代久远且数据量大,记录在标签上的部分信息存在缺失或字迹模糊等情况,在数字化过程中导致出现错误信息。为保证数据质量评估的准确性,本研究采用如下步骤对初步筛选的数据进行校对:(1) 植物拉丁名校对。利用 (http://tnrs.iplantcollaborative.org/TNRSapp.html) 网站将标本拉丁名进行剔除错误数据与标准化,将查询标本拉丁名准确率得分在 0.9 以下的标本数据剔除,准确率得分在 0.9 以上的接收为正确拉丁名。(2) 剔除采集地点缺失以及在中国行政区以外的采集记录。 (3) 对于采集地点在自然保护区内的标本记录,根据电子地图查询自然保护区所在地,若完整保护区在某个县级地区内,将保护区名改为该县名。(4) 县级行政单位名称校对。根据中国县级行政区划地图确定县名。部分旧县名经历行政区划的合并和重新划分,经查找其历史沿革后,确定最新的县名。经过整理后共获取了约 62,175 条地标化至县级的兰科植物标本数据,共包含有兰科植物 1,583 种。
1.2 环境因子的获取
从世界气候数据库中获取 19 个生物气候因子 (http://www.worldclim.org/),并将其进一步归类为能量因子和水分因子。为了消除因子之间的多重共线性,采用 Spearman 秩相关法计算因子之间的相关系数,对于同类型的因子若二者相关系数 >0.75,则依据其生态学意义选择其一。经过筛选后共保留四个能量因子 (AMT:年平均温度;MDR:气温日较差;ISO:等温性 (气温日较差/气温年较差) ;TAR:气温年较差;) 和两个水分因子 (AP:年平均降水;PS:降雨量季节性变化)。
1.3 物种丰富度与 Chao1 估计
由于稀有种的存在和野外调查限制,植物群落中被记录的物种数量一般要少于真实的物种数量。Chao (1984) 提出了一种基于稀有种数量的群落丰富度估计方法,通常被称为 Chao1 估计量[9]。Shen 等 (2003) 和 Chao 等 (2006b) 证实,对于许多数据集来说,它是一个群落总体物种丰富度的相对较好的点估计量。Chao1 指数是指在一个群体中随机抽样,若稀有种(Singletons)不断的被发现,则表明还有其他稀有种未被发现;直到所有物种至少被抽到两次 (Doubletons) 时,则表明不会再有新的物种被发现。Chao1 是度量物种丰富度的指标,它和多度、均匀度无关,但是它对稀有种很敏感。因此,本研究选取物种丰富度 Chao1 估计量来估计中国每个县真实的兰科植物物种丰富度。Chao1 的估计是基于已有取样中物种个体数量的分布频率来进行的 (Chao 1984; Colwell and Coddington 1994)[9,10],当取样中每个物种都是两个或两个以上的个体时,Chao1 认为该地区的取样比较完全;而当取样中有物种只有一个个体时,Chao1 认为该地区仍有新物种有待发现。其计算方程为:
其中,Sobs是观测到的物种数量,a 是取样中只有一个个体的物种数量,b 是只有两个个体的物种数量, S*是 Chao1 估计的物种数量。也就是每个县中稀有种数量越多,根据 Chao1 我们认为该地区新物种未被发现的概率越大;而每个县中稀有种数量越少,根据 Chao1 我们认为该地区新物种未被发现的概率越小。在计算过程中我们将稀有种个数定为 10。其它物种丰富度估计模型,如 Jackknife 和 Bootstrap,这些模型得出的估计结果和 Chao1 的结果基本相似。
1.4 标本采集记录完整性评估
本研究用两种方法来评估每个县兰科植物物种在该地的采集数据的不完整程度。第一种方法计算县级行政单位的观测的物种数量和 Chao1 估计量之间的参差来表示某一个县物种在该地的采集数据的不完整程度。第二种方法是利用物种累积曲线 (species accumulation curve, SAC),物种累积曲线用于描述即在已有采集记录随机抽样,随着抽样强度增加,物种丰富度积累多少亦增加,是理解调查样地物种组成和预测物种丰富度的有效工具,在生物多样性和群落调查中,被广泛用于抽样量充分性的判断以及物种丰富度估计[11]。
用物种累积曲线的弯曲程度来表示物种的不完整程度[12]。本研究采用的假设为:对于每一个县级行政单位,取样越不完整则对应的物种累积曲线越近于直线,而取样越完整则与之对应的物种累积曲线的弯曲度越高,物种积累曲线的弯曲程度可以用物种积累曲线尾端 10% 的斜率表示。物种累积曲线用R程序包“vegan”中的“specaccum”函数构建,方法为“exact”[13],该方法计算的是当对县的样本进行重抽样时,抽取 n 个个体时,这 n 个个体包含的平均 (或期望) 物种数量。构建光滑的物种累积曲线的假说前提是标本采集和物种的分布在时间和空间上都是随机的[10,14]。我们计算物种累积曲线尾端 10% 部分的平均斜率来表示物种各积累曲线的弯曲程度,代表某个县采集数据的不完整程度。当斜率值近于 0 时,说明取样较为完整,而当斜率近于 1 时,说明取样十分不完整。该斜率值还意味着如果采集强度变化,有新物种被发现的概率。将物种累积曲线尾端 10% 斜率域值定义为 0.05,当斜率值小于或等于 0.05 (SAC slope≤0.05) 时,认为该县的采集数据较为完整,斜率值大于 0.05 (SAC slope > 0.05) 时,则认为该县的采集数据不完整。
1.5 兰科植物标本采集完整性的环境因子解释
将标本采集数量、原始丰富度、丰富度估计值、丰富度残差和物种积累曲线尾端 10% 斜率五个参数分别作为因变量,环境因子作为自变量,采用逐步回归的方法对其进行拟合。逐步回归的基本思路是将环境因子逐个引入回归模型每引入一个解释变量后都要进行 F 检验,并对已经选入的解释变量逐个进行t检验,如果原来引入的解释变量由于后面解释变量的引入变得不再显著时,则将其删除。以确保每次引入新的变量之前回归方程中只包含显著性变量。逐步回归在 SPSS 15.0 软件中进行。
2 结果
从采集数量来看,兰科植物标本的采集集中在四川省和云南省,其中四川省南川市和峨眉山市的采集数量最为丰富,兰科植物标本数量分别为 2,277 份和 2,022 份。在云南省中部和东南沿海地区兰科植物标本的采集强度较低,有标本采集记录的县级行政单位呈零星分布 (图 1,A)。兰科植物物种丰富度记录数和 Chao1 估计量最高的地区为云南省勐腊县、贡山独龙族怒族自治县和勐海县,物种丰富度较高地点多集中在热带和亚热带地区,呈现出一定的纬度梯度,与我国被子植物丰富度的空间格局有一致性。

图1 A.标本采集数量;B.原始丰富度 (物种数);C.丰富度估计值;D.残差 (丰富度估计值-实际值);E.物种积累曲线尾端 10% 斜率。Fig.1 A: Number of collected specimens; B: Original richness; C: Richness estimates; D: Richness residue (estimated value minus actual value); E: Gradient of the last 10% section of the accumulated species curve
物种丰富度的 Chao1 估计量和记录值残差最高的地区为元江哈尼族彝族傣族自治县、彭水苗族土家族自治县和新平彝族傣族自治县 (图 1,D),残差值>100 种,代表区域内的稀有种数量较多。物种积累曲线尾端 10% 斜率代表标本采集的完整程度,以0.05为阈值,在有标本采集记录的县级行政单位中,仅有 38 个县兰科植物标本采集较为完整,占总数的 9.8%。采集最不完整的县 (物种积累曲线尾端 10% 斜率最大) 与物种丰富度残差值最高的县级行政单位一致,分别为:元江哈尼族彝族傣族自治县、彭水苗族土家族自治县和新平彝族傣族自治县 (图1,E)。表明 Chao1 估计量和物种积累曲线都能够对标本采集完整性进行评估,两种方法的评估结果具有相关性。
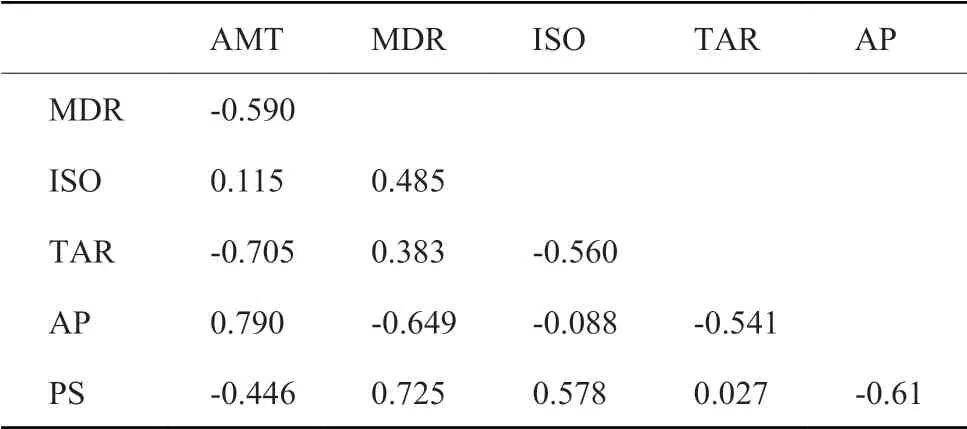
表1 环境因子(能量因子和水分因子)之间的Spearman秩相关系数。AMT:年平均温度;MDR:气温日较差;ISO:等温性(气温日较差/气温年较差);TAR:气温年较差;AP:年平均降水;PS:降雨量季节性变化。Table 1 Spearman coefficients of the environmental factors (between energy and moisture).AMT: average annual temperature; MDR: daily temperature variance; ISO: isothermality (daily/annual temperature variance); TAR: annual temperature variance; AP: annual average precipitation; PS: seasonal precipitation variance
环境因子对标本采集数量、丰富度残差和物种积累曲线尾端 10% 斜率解释能力较弱,对丰富度记录数和 Chao1 估计量解释能力较强 (表 2)。在逐步回归中,环境因子等温性与标本采集数量、原始丰富度、估计丰富度以及丰富度残差之间呈显著关系,且均为正相关。水分因子对各自变量的解释能力较弱。对原始丰富度和丰富度估计值的逐步回归中发现,等温性的解释能力最强 (正相关),在引入气温日较差因子后 (负相关),回归模型的解释能力分别可达 15.6% 和 16.9%。
3 讨论
植物标本的采集是一个长期的历史过程。我国国土面积广阔,植被类型多样,生境类型复杂,地形地貌复杂多变,系统化的进行标本采集几乎很难实现,广东省、广西省、福建省和浙江省等省份都有丰富的兰科植物文献记载,但是标本采集地点呈零星分布 (图 1,A)。兰科植物标本从采集数量来看多集中在我国西南地区 (四川省和云南省) ,该地区有着丰富多样的小生境类型,是我国著名的植物特有种中心,稀有种数量种类丰富[15],有分子生物学证据表明,该地区地形的复杂性造成了植物种间隔离,加速了物种的分化[16]。从采集地点来看,西南地区多各县级行政单位并没有兰科植物标本采集记录,尤其是四川省和云南省交界的三江源地区和云南省中部地区,我们认为造成这种空缺的原因可能是缺乏专门针对兰科植物的标本采集,或者是兰科植物分类学家在此地区的采集活动较少。
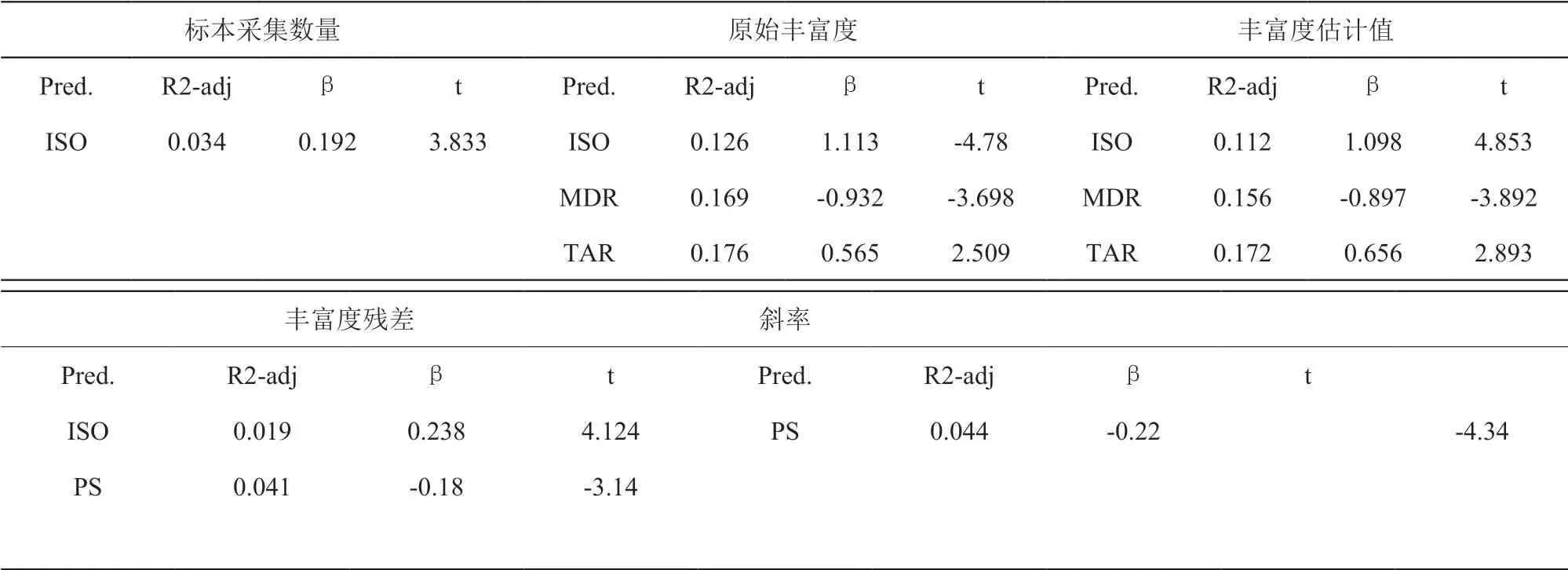
表2 环境因子与标本采集数量、原始丰富度、丰富度估计值、丰富度残差和物种积累曲线尾端 10% 斜率之间逐步回归结果Table 2 Results of gradual regression among environmental factors, collected specimen quantity, original richness, richness estimates, richness residue, and gradient of the last 10% section of the species accumulated curve
兰科植物丰富度和丰富度估计值最高的区域出现在云南省的西双版纳地区和三江并流地区,与云南省木本种子植物丰富度分布格局相一致[17]。在西双版纳等热带地区,兰科植物多附生在热带雨林的树干、树枝或枯木上,根部吸收空气中的水分,附生兰科植物通常要求生境有着较高的温度和湿度。在三江并流地区,造成兰科植物丰富度较高的原因是由于种间隔离造成了物种的分化。现存的兰科植物都是由共同的祖先分化而来,兰科植物形态和习性变异多样,花部结构高度特化,是进化程度最高的类群之一[18-19],兰科植物的繁殖过程高度依赖虫媒传粉,因此地理隔离极易阻碍物种间的基因交流,促进了种间分化。
标本采集的完整性采用两种方法进行评估,本研究中两种评估方法得出的结果具有相似性 (图 1,D;图1,E)。仅有少于 10% 的县级行政单位被评估为采样完全,表明未来野外调查的任务仍然十分艰巨。兰科植物类群缺乏广布种,对其生境依赖性较强,且散布能力较弱,上述原因都造成了兰科植物稀有种数量较多,因此在野外调查的过程中遗漏率也较高,极易造成采样不完全现象的出现。因此,针对专科专属的普查十分必要。有限的分布范围和较多的稀有种数量也增加了本类群植物的濒危等级,对于兰科植物的保护生物学研究有着重要意义。
气温的稳定性 (等温性和气温年较差) 与兰科植物丰富度显著相关,即兰科植物种类最高的区域出现在温度稳定性较高的地区。Zhang 等 (2014) 分析了气候变化对云南省木本植物分布格局的影响,结果发现温度的稳定性是最重要的决定因子,当气温发生波动时,极易造成物种的本地灭绝。在亚马孙和婆罗洲的热带雨林群落中也发现,温度的稳定性对于热带和亚热带地区生物多样性空间格局的形成有着显著影响[20-21]。由于兰科植物类群分布的热带和亚热带地区温度较高并且湿润多雨,总能量输入和总降雨量并不是其限制因子。兰科植物通常是狭域分布[22],在野外易形成极小种群,濒危等级较高。上述因素造成了兰科植物的分布通常依赖小生境 (微环境),对外部环境的适应能力较弱,当气温发生波动或者受到干扰时,兰科植物种群较为敏感,在对兰科植物进行保护生物学实践中应综合考虑上述因素的影响。
猜你喜欢
物理之友(2020年12期)2020-07-16
科学大众(中学)(2019年3期)2019-05-17
陕西农业科学(2019年3期)2019-04-09
汽车观察(2018年10期)2018-11-06
科技知识动漫(2017年1期)2017-02-06
福建中学数学(2016年7期)2016-12-03
光学精密工程(2016年1期)2016-11-07
电测与仪表(2016年6期)2016-04-11
新疆农业科技(2015年2期)2015-08-01
少儿科学周刊·少年版(2015年1期)2015-07-07
