
子午岭典型植被凋落叶-土壤养分与酶活性特征
2018-05-16 08:42张海鑫曾全超安韶山王宝荣白雪娟
生态学报 2018年7期
张海鑫,曾全超,安韶山,王宝荣,白雪娟
1 西北农林科技大学资源环境学院,杨凌 712100 2 西北农林科技大学水土保持研究所,杨凌 712100
黄土高原子午岭林区属于落叶阔叶林带,地带性植被为辽东栎(Quercuswutaishanica)林[1]。在清同治年间(1842—1866)陕甘交界地区植被开始自然恢复,目前子午岭已经形成较大范围而且连片的次生森林景观[2]。有研究表明,子午岭天然林与人工林地土壤水分状况存在明显的差异,辽东栎林地土壤水分基本没有形成亏缺,而刺槐林地土壤水分不管是深层还是浅层,都形成了较严重的亏缺状况[3]。该区乔木能够显著地促进土壤中氮素的富集,有效改善土壤氮素养分状况且养分具有明显的表聚作用,刺槐对土壤氮和磷的利用效率比油松高[4- 6],这些学者的工作主要集中在不同植被恢复方式下土壤养分及水分的研究,而对不同植被恢复方式下土壤酶活性的研究较少。土壤酶是土壤中具有生物活性的蛋白质,在土壤物质循环和能量转化过程中起着重要作用[7- 8],它的活性强弱可以较直接反映出土壤生化反应的方向和强度。对土壤酶活性的研究有助于了解土壤肥力状况和演变规律[9]。土壤脲酶、蔗糖酶及磷酸酶为水解酶;脲酶活性高低可以反映土壤氮素营养状况[10- 11],蔗糖酶对增加土壤中易溶性营养物质起重要作用,磷酸酶活性的高低可以评价土壤磷酸生物转化方向与强度[12]。
凋落叶是森林生态系统养分循环的基础,在维持土壤肥力方面起特别重要的作用[13]。凋落叶淋溶、分解后,将化学元素释放出来,归还到土壤养分库中。碳是构成植物体内干物质的最主要成分,氮和磷是陆地生态系统植物生长的主要限制因子[14],在植物养分方面一直倍受关注[15- 16]。凋落物的碳氮比值与其分解速率成反比关系,碳氮比值越小,其分解速率越快[17]。另有研究表明,土壤碳氮比值的变化将影响微生物的组成及活性,进而影响有机碳的分解速率[18]。目前在子午岭林区结合五种典型植被的凋落叶、土壤理化性质及酶活性三者的研究较少,鉴于此,本文分析子午岭林区不同植被凋落叶中各元素与土壤中各元素相关关系及碳氮比,并对土壤理化性质与土壤酶活性进行进一步研究,找出影响森林土壤酶活性的因子,以期为子午岭林区人工林种植及植被恢复效果评价提供科学依据。
1 研究地区与研究方法
1.1 研究区概况
研究区位于陕西省子午岭任家台林区(36°03′10″—36°05′27″N,109°07′3″—109°10′36″E),海拔 1163—1305m,属于国家级自然保护区,是暖温带半湿润地区,年均气温7.4℃,年降水量587.6mm,年平均相对湿度63%—68%,无霜期140—160d。土壤为原生或次生黄土,厚度约为50—100m。 植被多分布于阴坡及半阴坡,主要人工林植被有刺槐(Robiniapseudoacacia)林、油松(Pinustabuliformis)林和侧柏(Platycladusorientalis)林等,天然次生林植被有桦树(Betulaplatyphylla)林、辽东栎(Quercuswutaishanica)林等(表1)。气候性的演替顶极为辽东栎林[19]。
1.2 样点布设与样品采集
2015年9月初,在子午岭林区选择海拔、盖度等较为接近的、林相整齐、林木分布均匀、平均林龄约30a的坡位及坡向较为一致的刺槐、油松、侧柏、桦树、辽东栎等5种植被林地作为采样点(70m×40m),每个植被在野外选取3个采样点,在每个样点内设置3个20m×20m的样方。
9月初,叶片开始凋落。在每个样方内沿对角线方向设置3个1m×1m小样方,其中在刺槐、桦树、辽东栎样地内随机地选择腐熟程度一致(颜色相近、且颜色较浅)的新近凋落叶,在油松、侧柏样地内选取刚刚掉下来的,且颜色为浅棕黄色的叶片,进行多点采集,混匀后装入牛皮纸袋,带回实验室于105℃下杀青约10min,在65℃下烘48h至恒重、样品经研磨过100目筛(孔径为 0.15mm)后进行元素的测定。在样方内按照“S”型路线取5个点除掉上层凋落物层,用土钻采集0—20cm土层土样,混合均匀后带回实验室,土样自然风干后过1mm和0.25mm筛备用。
1.3 样品测定
不同植被凋落叶中有机碳采用重铬酸钾-外加热法测定;全氮、全磷采用 H2SO4-H2O2消煮法,消解后的待测液供全氮、全磷的测定,其中全氮采用凯氏定氮仪测定(KDY- 9830,KETUO),全磷采用钒钼黄比色法测定(UV- 2450 紫外分光光度计)。土壤容重的测定采用环刀法。
土壤pH值:采用水浸提电位法测定,用无CO2去离子水调节土水比为1∶5,搅拌1min,静置30min后,用DMP- 2mV/pH计测定。土壤有机碳采用重铬酸钾-外加热法测定,土壤全氮测定采用硫酸-混合催化剂消煮,消煮液中的全氮测定采用凯氏定氮仪(KDY- 9380, KETUO)测定,全磷采用HClO4-H2SO4消煮后进行钼蓝比色法比色,利用UV- 2450紫外分光光度计测定[20]。
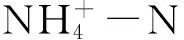
1.4 数据处理
试验数据采用 Microsoft Excel 2010和SPSS 21.0 统计分析软件进行整理。采用One-Way ANOVA (Scheffe) 对不同植被凋落叶、土壤相应的碳、氮、磷含量以及土壤酶活性进行差异性检验。利用OriginPro 8.0软件作图。图表中数据为平均值±标准差。
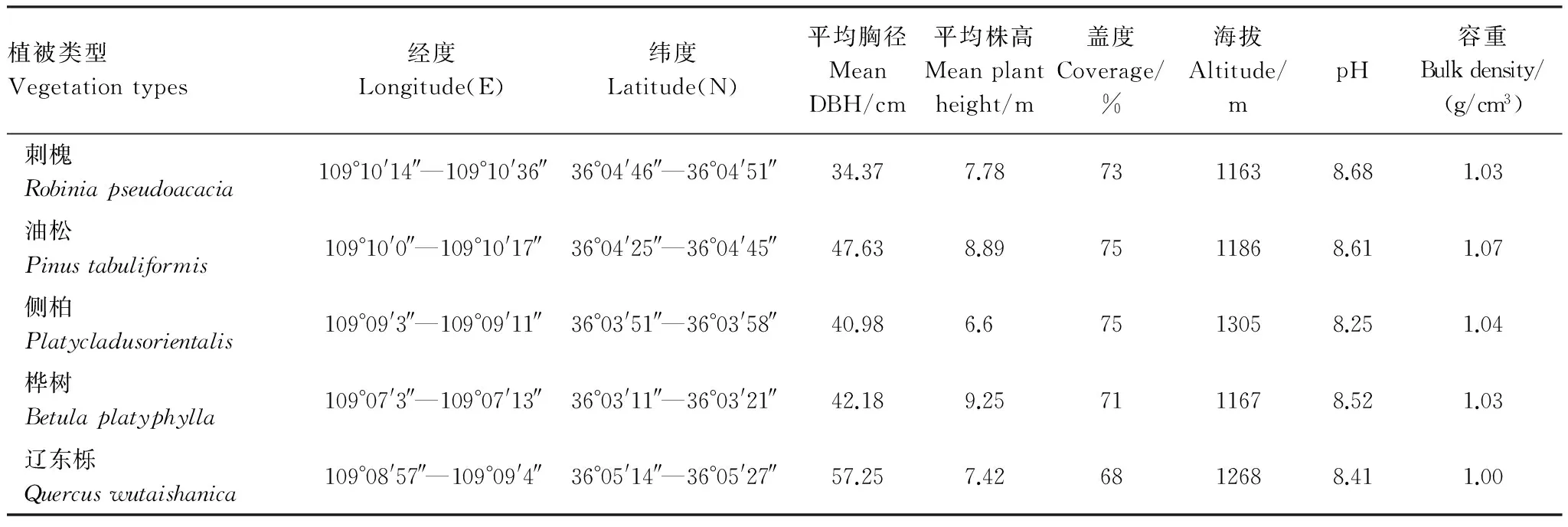
表1 样方基本信息
2 结果与分析
2.1 不同植被凋落叶碳、氮、磷含量变化及碳氮比
由图1可以看出,不同植被凋落叶碳氮磷含量变化范围分别为398.22—501.02、5.34—17.10、1.41—1.83g/kg,其中油松凋落叶全碳含量最高,刺槐凋落叶全氮含量最高,辽东栎凋落叶全磷含量最高。油松、侧柏、桦树凋落叶全碳含量显著高于刺槐和辽东栎(P<0.05)。辽东栎、刺槐凋落叶全氮含量显著高于侧柏和桦树(P<0.05),油松凋落叶全氮含量最低。辽东栎凋落叶全磷含量显著高于其他4个植被(P<0.05),刺槐、油松、侧柏和桦树凋落叶全磷含量无显著性差异。刺槐、油松、侧柏、桦树和辽东栎凋落叶碳氮比值分别为23.75、95.06、54.36、59.35和26.87,其中油松凋落叶碳氮比值显著高于其他4个植被(P<0.05)。
2.2 不同植被凋落叶和土壤碳、氮、磷含量相关关系及碳氮比
不同植被土壤碳氮磷含量变化范围6.76—19.18、0.70—1.60、0.55-0.61g/kg,其中辽东栎土壤有机碳、氮含量最高,刺槐土壤全磷含量最高。辽东栎土壤有机碳含量分别是侧柏和桦树的1.18倍和1.19倍,是油松和刺槐1.60倍和2.84倍。辽东栎和侧柏土壤全氮含量显著高于油松和桦树(P<0.05),刺槐土壤全氮含量最低。刺槐土壤全磷含量显著高于侧柏、桦树、辽东栎,与油松差异性不显著(P<0.05)。桦树土壤碳氮比值显著高于刺槐、油松(P<0.05),辽东栎土壤碳氮比值显著高于油松(P<0.05),而刺槐、油松、侧柏土壤碳氮比值差异性不显著。
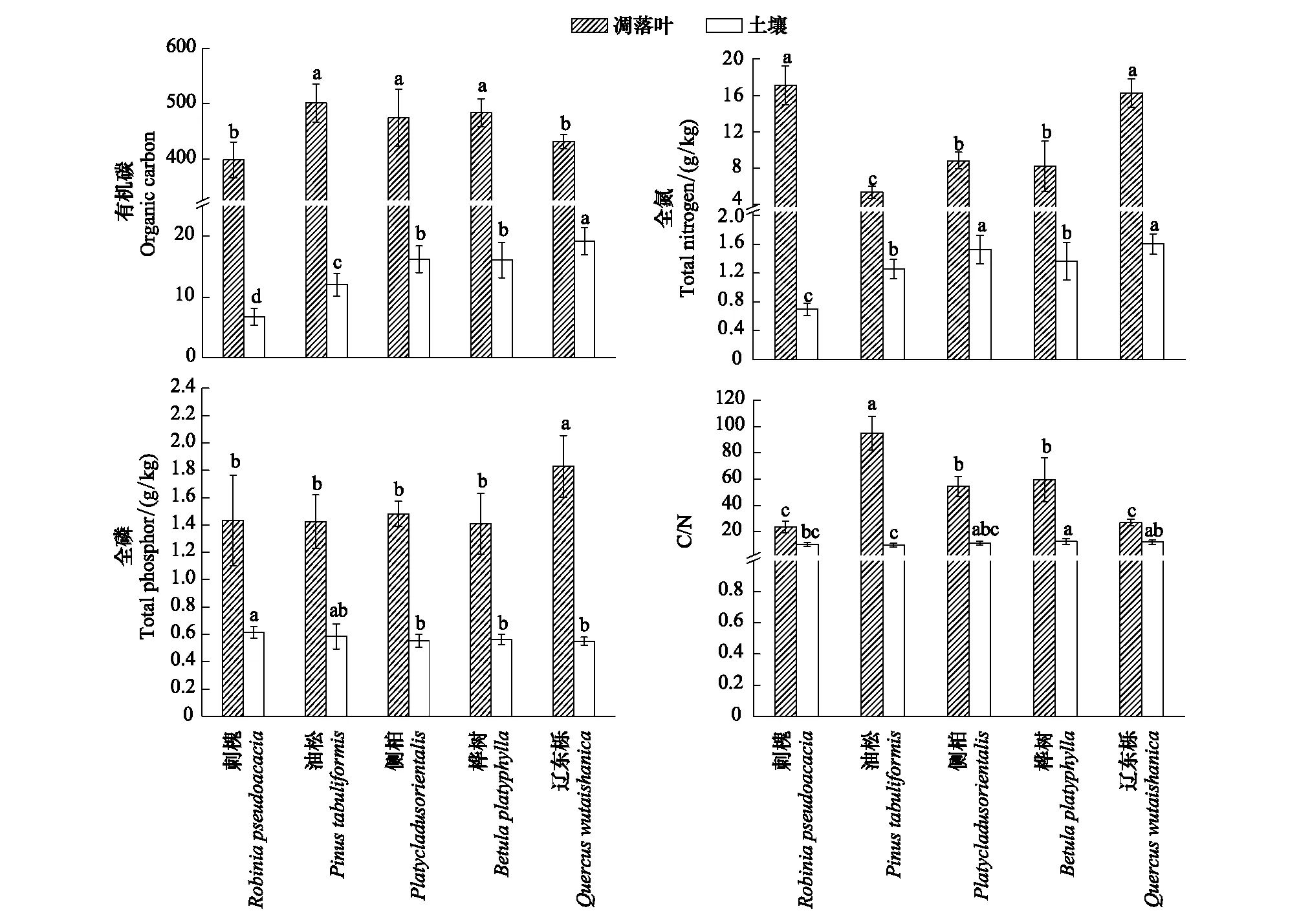
图1 不同植被凋落叶和土壤的碳氮磷含量和碳氮比Fig.1 Carbon, nitrogen, phosphorus contents and C/N of litter and soil in different vegetation types不同小写字母表示同一组分不同植被间差异显著(P<0.05)
由表2可以看出,土壤有机碳与凋落叶全碳、全磷之间呈显著的正相关(P<0.05),相关系数分别为0.228、0.255,与凋落叶全氮之间没有显著的负相关(P>0.05)。土壤全氮与凋落叶全碳之间呈极显著正相关(r=0.319,P<0.01),与凋落叶全氮之间呈显著负相关(r=-0.266,P<0.05)。土壤全磷与凋落叶全氮之间呈显著正相关(r=0.249,P<0.05),与凋落叶全碳没有显著的负相关(P>0.05)。

表2 凋落叶与土壤养分含量之间的相关关系
** 在 0.01 水平上显著相关;* 在 0.05 水平上显著相关(df=76);OC:Organic carbon;TN: Total nitrogen;TP:Total phosphorus
2.3 不同植被土壤酶活性的变化
试验样地内的土壤pH为8.25—8.68,均为碱性土(表1),因此本实验测定的磷酸酶为碱性磷酸酶。由图2可以看出,侧柏土壤碱性磷酸酶含量为 1.23mg g-1d-1,分别是刺槐、油松、桦树、辽东栎的1.89、1.17、1.26、1.33倍。辽东栎土壤脲酶含量为0.26mg g-1d-1,分别是刺槐、油松、侧柏、桦树的1.72、1.71、1.13、1.34倍。侧柏土壤蔗糖酶含量为64.50mg g-1d-1,分别是刺槐、油松、桦树、辽东栎3.81、1.30、2.95、1.27倍。
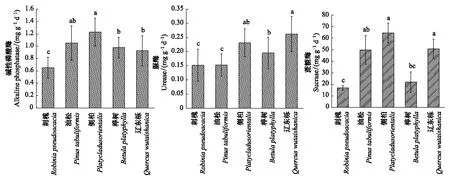
图2 不同植被土壤的酶活性Fig.2 Soil enzyme activities in different vegetation types
2.4 不同植被土壤酶活性与土壤理化性质之间的关系
表3表示土壤酶活性与土壤理化性质之间的相关关系。碱性磷酸酶与土壤有机碳、全氮呈极显著正相关(r=0.300和r=0.420,P<0.01),与土壤容重呈正相关,与土壤pH呈显著负相关(P<0.05)。脲酶与土壤有机碳、全氮呈极显著正相关(r=0.528 和r=0.534,P<0.01),与土壤容重、pH之间不存在显著相关关系。蔗糖酶与土壤有机碳、全氮呈极显著正相关(r=0.527和r=0.530,P<0.01),与土壤容重呈显著正相关(r=0.310,P<0.05),与土壤pH呈极显著负相关(r=-0.556,P<0.01)。碱性磷酸酶、脲酶、蔗糖酶与土壤全磷之间均不存在显著相关关系。碱性磷酸酶与蔗糖酶呈极显著正相关(r=0.444,P<0.01),脲酶与蔗糖酶呈显著性正相关(r=0.310,P<0.05)。
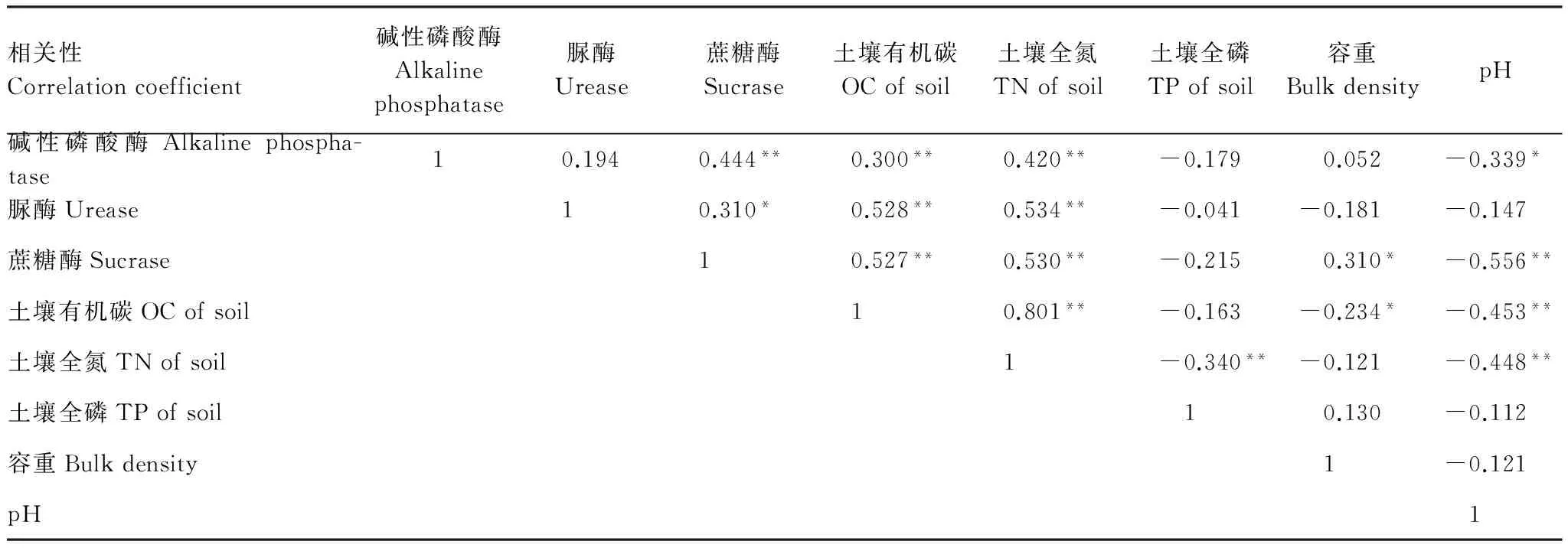
表3 土壤酶活性与土壤基本理化性质相关关系
** 在 0.01 水平上显著相关;* 在 0.05 水平上显著相关(df=74)
3 讨论
3.1 不同植被凋落叶碳、氮、磷含量变化及碳氮比
凋落叶的类型、化学组成、矿化速率等在一定程度上影响森林土壤的理化性质。由于地表凋落叶的不断分解,使得矿质养分元素源源不断地归还到土壤之中,使有限的养分元素被植物循环利用,既保持土壤肥力,又维护生态系统的平衡。王清奎等[22]认为在森林生态系统中,凋落叶是土壤速效养分和有机质的主要来源,是土壤有机碳积累的主要途径,给土壤中异养微生物提供碳源。凋落叶作为联系植物体和土壤的载体对其营养元素的研究有重要意义[6]。陕西省森林群落凋落物三大元素含量分别为C (367.40g/kg)、N (12.03g/kg)、P(1.44g/kg)[23]。本试验中,刺槐、油松、侧柏、桦树、辽东栎凋落叶全碳含量分别为398.22、501.02、474.35、483.29、431.37g/kg,均高于陕西省森林群落凋落物平均碳水平,表明各植被固定碳能力较强。刺槐、辽东栎全氮含量分别为17.10、16.26g/kg,均高于陕西省森林群落凋落物平均氮水平,而油松、侧柏、桦树全氮含量分别为5.34、8.81、8.21g/kg,均低于陕西省森林群落凋落物平均氮水平。同时5个植被凋落叶全氮含量与全球木本凋落叶N含量(10.9g/kg)相比,刺槐、辽东栎全氮含量均高于全球木本凋落叶N水平,而油松、侧柏、桦树全氮含量均低于全球木本凋落叶N水平[24]。侧柏与辽东栎凋落叶全磷含量分别为1.48、1.83g/kg,均高于陕西省森林群落凋落物平均磷水平,而刺槐、油松、桦树凋落叶全磷含量分别为1.43、1.42、1.41g/kg,均低于陕西省森林群落凋落物平均磷水平。同时5个植被凋落叶全磷含量均高于全球木本凋落叶P含量(0.85g/kg)[24]。本研究区各植被凋落叶表现出高C、高P,部分植被低N的现象。油松、侧柏、桦树全氮含量略低,可能是由于植被类型、当地环境因素的差异所导致的。
Moore等[25]在加拿大12a的凋落物分解研究表明:多数凋落物碳氮比在33—68时,氮释放。本研究中,油松凋落叶碳氮比值为95.06,明显高于68,凋落叶全氮含量最低,刺槐、辽东栎凋落叶碳氮比值分别为23.75、26.87,虽然低于33,其凋落叶全氮含量均高于侧柏和桦树。李雪峰等[26]对长白山次生针阔混交林凋落叶分解与碳、氮释放关系研究表明,分解速率与碳氮比值呈显著负相关,碳氮比值越小,其分解速率越快,有利于养分循环[17]。本研究中5个植被凋落叶碳氮比值为:刺槐<辽东栎<侧柏<桦树<油松,因此其凋落叶分解速率表现为刺槐>辽东栎>侧柏>桦树>油松,虽然刺槐凋落叶分解速率快,但其林下土壤全氮含量最低,可能受自身植被类型影响。
3.2 不同植被凋落叶和土壤碳、氮、磷含量相关关系及土壤碳氮比
土壤养分组成是植物生长极其重要的影响因子, 植物光合作用、矿质代谢等过程与土壤养分供应状况紧密相关[27]。森林土壤有机养分来源的主要途径是林地枯落物。研究不同植被类型的枯落物分解速率,对于研究土壤有机质含量变化及土壤肥力变化,显得十分重要[28]。刺槐凋落叶全碳含量略低于其他4个植被,而全氮含量高于其他4个植被,表明刺槐叶片固氮能力较强,但其土壤碳氮肥力低于其他植被,表明刺槐凋落叶分解后养分归还于土壤的过程中,大部分碳氮养分有所流失,因此刺槐能否作为该区的人工造林植被还需进一步考察。本试验发现林下土壤全磷含量受凋落叶全氮含量影响较大,森林土壤养分(N、P等)主要来源于凋落叶的分解,另外受地表草本植被覆盖度、水热条件等其他环境因素影响。
土壤碳氮比值作为影响土壤有机碳分解的重要因子,其变化将对土壤有机碳动态产生重要影响。本试验中,刺槐、油松、侧柏、桦树和辽东栎土壤碳氮比值分别为10.26、9.70、11.13、12.60和12.06,均低于全球土壤碳氮比值平均水平(13.3),但符合我国湿润温带土壤碳氮比值在10—12的规律[29]。 土壤有机层的碳氮比值较低表明有机质具有较快的矿化作用。土壤有机质通过影响土壤物理、化学、生物学性质进而影响植被的生长状况[30],有机质分解后可以为土壤中微生物提供代谢能量[31]。桦树土壤碳氮比值显著高于刺槐、油松(P<0.05),辽东栎土壤碳氮比值显著高于油松(P<0.05),而刺槐、油松、侧柏土壤碳氮比值差异性不显著,说明刺槐、油松、侧柏土壤有机质分解速度相近,而桦树和辽东栎土壤有机质分解速度较快。
3.3 不同植被土壤酶活性变化
植被类型的不同使得其凋落叶、土壤理化性质、土壤微生物量有差别,因此酶活性也会有差异。蔗糖酶、脲酶以及碱性磷酸酶分别能够表征土壤C、N和P的供应程度,土壤酶活性是土壤质量、生态环境效应评价中极为重要的指标之一,它在评价土壤肥力、环境监测、和土地利用等方面有重要意义[32- 34]。酶活性受季节变化影响,脲酶、碱性磷酸酶活性随季节的变化为先增加后降低再增加再降低,蔗糖酶活性随季节的变化表现为先升高后下降的趋势[35],酶活性在夏季和秋季较高[36]。本试验采样时间为9月初,酶活性处于相对较高时期。本试验中天然次生林桦树、辽东栎3种土壤酶活性均高于人工林刺槐,在人工林中,侧柏的3种土壤酶活性均高于刺槐和油松。土壤酶系统是土壤中生物活性最强的部分,其与植物生长之间存在必然的内在联系,因此植被类型是影响土壤水解酶活性的关键因子之一。同时,土壤中微生物通过酶催化分解大分子有机物为可溶性的养分,供微生物同化利用[37],其中水解酶活性直接影响纤维素、半纤维素的分解[38],影响着森林有机质周转,进而影响森林的生产力和可持续性。
3.4 不同植被土壤酶活性与土壤理化性质之间的关系
周礼恺等[39]认为土壤的异质性决定了土壤酶的种类和数量上的差异性。也有研究表明,影响土壤中水解酶活性因素很多,包括土壤养分、植被类型的影响和土壤动物等生物因素,以及凋落物质量、土壤团聚体、海拔[40]和人为干扰等[41]。由表3可以看出,影响酶活性的关键因素是土壤有机碳、全氮含量,同时碱性磷酸酶、蔗糖酶活性受pH影响较大。土壤氮、磷是土壤微生物必需的营养元素[42],影响有机碳、全氮的重要因素是脲酶,土壤养分与土壤脲酶有密切的相关关系[43- 44]。土壤酶与有机质密切相关,有机质含量高时土壤酶活性可以保持较高水平[45]。随着土壤碳氮比值的提高,微生物活动所需的碳源增加,其分解代谢趋向活跃,微生物量碳周转加速,因此土壤有机碳的累积矿化量及矿化速率都明显提高[46]。因此,土壤碳氮比变化会引起土壤微生物活性及矿化率变化,进而影响土壤质量和土壤碳氮循环[47]。通过比较土壤碳氮比值,可以了解到人工林土壤有机质分解速度相近,而天然次生林土壤有机质分解速度较快。虽然刺槐、油松、侧柏土壤有机质分解速度相近,而侧柏林地土壤酶活性比刺槐、油松高,且侧柏林地土壤有机碳、氮含量高于刺槐和油松林地,表明侧柏较其他两个人工植被能更好地改善该地区土壤养分状况。刺槐土壤有机碳含量低于其他植被,同时水解酶活性表现出低于其他四种植被。而在黄土高原退耕坡地不同植被类型土壤酶活性研究中发现刺槐林地土壤脲酶、碱性磷酸酶和蔗糖酶均表现出较高的活性,反映了其良好的氮素和磷素供应状况以及较强的分解土壤中营养物质的能力[34],与本试验研究结果不同,表明同一植被类型下土壤酶活性受地理环境因素影响较大。有研究指出水解酶活性与有机质、全氮、全磷之间呈显著正相关,而与pH 呈显著负相关[48],而本试验结果表明水解酶活性与有机碳、全氮呈显著正相关,而与全磷、pH呈负相关,另外蔗糖酶活性受土壤容重影响较大。
4 结论
(1) 通过分析比较5种植被凋落叶、土壤碳氮磷含量以及碳氮比值,发现刺槐凋落叶含有大量养分,但对土壤酶活性及理化性质影响较小,其作为人工造林树种有待进一步考察,而侧柏相对表现出优势,能更好地改善该地区土壤碳氮肥力及酶活性。
(2)土壤酶活性主要受植被类型、土壤有机碳、全氮、容重及pH影响,与土壤全磷相关性不显著。植被恢复与人工造林过程中土壤有机碳和全氮含量的增加是提高土壤酶活性的主要因素。
(3) 人工造林虽然对该区土壤肥力的恢复和酶活性的提高有一定作用,但不及天然次生林。天然次生林辽东栎为顶级群落,植被恢复过程中对改善土壤化性状等占绝对优势,其土壤碳氮肥力及脲酶、蔗糖酶活性均高于桦树,且土壤容重最低。
参考文献(References):
[1] 朱志诚. 秦岭及其以北黄土区植被地带性特征.地理科学, 1991, 11(2):157- 163.
[2] 李裕元, 邵明安. 黄土高原子午岭森林群落演替与结构特征演化. 西北植物学报, 2003, 23(5): 693- 699.
[3] 王力, 邵明安, 王全九, 贾志宽. 黄土高原子午岭天然林与刺槐人工林地土壤干化状况对比. 西北植物学报, 2005, 25(7):1279- 1286.
[4] 吕春花, 郑粉莉, 安韶山. 子午岭地区植被演替过程中土壤养分及酶活性特征研究. 干旱地区农业研究, 2009, 27(2): 227- 232.
[5] 邢肖毅, 黄懿梅, 黄海波, 安韶山, 刘栋. 黄土丘陵区子午岭不同植物群落下土壤氮素及相关酶活性的特征. 生态学报, 2012, 32(5): 1403- 1411.
[6] 白雪娟, 曾全超, 安韶山, 张海鑫, 王宝荣. 黄土高原不同人工林叶片-凋落叶-土壤生态化学计量特征. 应用生态学报, 2016, 27(12): 3823- 3830.
[7] 史衍玺, 唐克丽. 人为加速侵蚀下土壤质量的生物学特性变化. 土壤侵蚀与水土保持学报, 1998, 14(1): 28- 33.
[8] 柳云龙, 吕军, 王人潮. 低丘侵蚀红壤复垦后土壤微生物特征研究. 水土保持学报, 2001, 15(2): 64- 67.
[9] 张萍, 郭辉军, 刀志灵, 龙碧云. 高黎贡山土壤微生物生化活性的初步研究. 土壤学报, 2000, 37(2): 275- 279.
[10] Tabatabai M A, Bremner J M. Assay of urease activity in soils. Soil Biology and Biochemistry, 1972, 4(4): 479- 487.
[11] Singh D K, Kumar S. Nitrate reductase, arginine deaminase, urease and dehydrogenase activities in natural soil (ridges with forest) and in cotton soil after acetamiprid treatment. Chemosphere, 2008, 71: 412- 418.
[12] 邱梅, 张海, 张宇, 张志强. 黄土高原不同坡位苹果林土壤酶活性及微生物量动态变化研究. 西北农业学报, 2014, 23(4): 152- 159.
[13] Guo L B, Sims R E H. Litter production and nutrient return in New Zealand eucalypt short rotation forests: implications for land management. Agriculture, Ecosystems & Environment, 1999, 73(1): 93- 100.
[14] Vitousek P M, Poder S, Houlton B Z, Chadwick O A. Terrestrial phosphorus limitation: Mechanism, implication, and nitrogen-phosphorus interactions. Ecological Applicaions, 2010, 20(1): 5- 15.
[15] 项文化, 黄志宏, 闫文德, 田大伦, 雷丕锋. 森林生态系统碳氮循环功能耦合研究综述. 生态学报, 2006, 26(7): 2365- 2372.
[16] 任书杰, 于贵瑞, 陶波, 王绍强. 中国东部南北样带654种植物叶片氮和磷的化学计量学特征研究. 环境科学, 2007, 28(12): 2665- 2673.
[17] 丁小慧, 宫立, 王东波, 伍星, 刘国华. 放牧对呼伦贝尔草地植物和土壤生态化学计量学特征的影响. 生态学报, 2012, 32(15): 4722- 4730.
[18] 廖建雄, 王根轩. CO2和温度升高及干旱对小麦叶片化学成分的影响. 植物生态学报, 2000, 24(6): 744- 747.
[19] 邹厚远, 刘国彬, 王晗生. 子午岭林区北部近50年植被的变化发展. 西北植物学报, 2002, 22(1): 1- 8.
[20] 鲍士旦. 土壤农化分析. 第三版. 北京: 中国农业出版社, 2000.
[21] 关松荫. 土壤酶及其研究法. 北京: 中国农业出版社, 1986: 274- 339.
[22] 王清奎, 汪思龙, 于小军, 张剑, 刘燕新. 杉木与阔叶树叶凋落物混合分解对土壤活性有机质的影响. 应用生态学报, 2007, 18(6):1203- 1207.
[23] 姜沛沛, 曹扬, 陈云明. 陕西省森林群落乔灌草叶片和凋落物C、N、P生态化学计量特征. 应用生态学报, 2016, 27(2): 365- 372.
[24] Kang H Z, Xin Z J, Berg B, Burgess P J, Liu Q, Liu Z, Li Z H, Liu C Z. Global pattern of leaf litter nitrogen and phosphorus in woody plants. Annals of Forest Science, 2010, 67(8): 811- 811, doi:10.1051/forest/2010047.
[25] Moore T R, Trofymow J A, Prescott C E, Titus B D, Cidet Working Group. Nature and nurture in the dynamics of C, N and P during litter decomposition in Canadian forests. Plant and Soil, 2011, 339(1/2): 163- 175.
[26] 李雪峰, 韩士杰, 胡艳玲, 赵玉涛. 长白山次生针阔混交林叶凋落物中有机物分解与碳、氮和磷释放的关系. 应用生态学报, 2008, 19(2): 245- 251.
[27] 宾振钧, 王静静, 张文鹏, 徐当会, 程雪寒, 李柯杰, 曹德昊. 氮肥添加对青藏高原高寒草甸6个群落优势种生态化学计量学特征的影响. 植物生态学报, 2014, 38(3), 231- 237.
[28] 杨承栋. 我国人工林土壤有机质的量和质下降是制约林木生长的关键因子.林业科学, 2016, 52(12):1- 12.
[29] 王绍强, 于贵瑞. 生态系统碳氮磷元素的生态化学计量学特征. 生态学报, 2008, 28(8): 3937- 3947.
[30] Maia S M F, Ogle S M, Cerri C C, Cerri C E P. Changes in soil organic carbon storage under different agricultural management systems in the southwest Amazon region of Brazil. Soil and Tillage Research, 2010, 106(2): 177- 184.
[31] Vanlauwe B. Integrated soil fertility management research at TSBF: the framework, the principles, and their application//Bationo A, ed. Managing Nutrient Cycles to Sustain Soil Fertility in Sub-Saharan Africa. Nairobi, Kenya: Academy Science Publishers, 2004: 25- 42.
[32] 杨宁, 邹冬生, 杨满元, 胡利珍, 邹芳平, 宋光桃, 林仲桂. 衡阳紫色土丘陵坡地不同恢复阶段植被特征与土壤性质的关系. 应用生态学报, 2013a, 24(1): 90- 96.
[33] Ren C, Kang D, Wu J P, Zhao F Z, Yang G H, Han X H, Feng Y Z, Ren G X. Temporal variation in soil enzyme activities after afforestation in the loess plateau, china. Geoderma, 2016, 282: 103- 111.
[34] 翟辉, 张海, 邱梅, 郝月姣, 张超. 黄土高原退耕坡地不同类型林分土壤生物学活性的研究. 西北林学院学报, 2016, 31(4):33- 38.
[35] 景宇鹏, 李跃进, 年佳乐, 蔺亚莉. 土默川平原不同盐渍化土壤酶活性特征的研究. 生态环境学报, 2013, 22(9):1538- 1543.
[36] 万忠梅, 宋长春. 土壤酶活性对生态环境的响应研究进展. 土壤通报, 2009, 40(4): 951- 956.
[37] Burns R G, DeForest J L, Marxsen J, Sinsabaugh R L, Stromberger M E, Wallenstein M D, Weintraub M N, Zoppini A. Soil enzymes in a changing environment: current knowledge and future directions. Soil Biology and Biochemistry, 2013, 58: 216- 234.
[38] Sinsabaugh R L. Phenol oxidase, peroxidase and organic matter dynamics of soil. Soil Biology and Biochemistry, 2010, 24(3): 391- 404.
[39] 周礼恺, 张志明, 曹承绵. 土壤酶活性的总体在评价土壤肥力水平中的作用. 土壤学报, 1983, 20(4): 413- 418.
[40] 樊金娟, 李丹丹, 张心昱, 何念鹏, 部金凤, 王情, 孙晓敏, 温学发. 北方温带森林不同海拔梯度土壤碳矿化速率及酶动力学参数温度敏感性. 应用生态学报, 2016, 27(1):17- 24.
[41] 曹慧, 孙辉, 杨浩, 孙波, 赵其国. 土壤酶活性及其对土壤质量的指示研究进展. 应用与环境生物学报, 2003, 9(1): 105- 109.
[42] 鲁如坤. 土壤农业化学分析方法. 北京: 中国农业科技出版社, 2000.
[43] 杨宁, 邹冬生, 杨满元, 陈璟,陈志阳, 林仲桂, 宋光桃. 衡阳紫色土丘陵坡地不同植被恢复阶段土壤酶活性特征研究. 植物营养与肥料学报, 2013b, 19(6):1516- 1524.
[44] 赵维娜, 陈奇伯, 王艳霞, 聂蕾, 杨媛媛. 高山栎天然林土壤有机质及酶活性的通径分析. 西部林业科学, 2016, 45(5): 58- 64.
[45] 隋跃宇, 焦晓光, 高崇生, 程伟, 张兴义, 刘晓冰. 土壤有机质含量与土壤微生物量及土壤酶活性关系的研究. 土壤通报, 2009, 40(5): 1036- 1039.
[46] 窦晶鑫, 刘景双, 王洋, 赵光影. 三江平原草甸湿地土壤有机碳矿化对C/N的响应. 地理科学, 2009, 29(5): 773- 778.
[47] 罗由林, 李启权, 王昌全, 张维, 张浩, 李林鲜, 陈俊伟, 马煜. 川中丘陵县域土壤碳氮比空间变异特征及其影响因素. 应用生态学报, 2015, 26(1): 177- 185.
[48] 李林海, 邱莉萍, 梦梦. 黄土高原沟壑区土壤酶活性对植被恢复的响应. 应用生态学报, 2012, 23(12): 3355- 3360.
猜你喜欢
新农民(2022年22期)2022-11-07
星星·诗歌原创(2021年3期)2021-06-01
露天采矿技术(2020年6期)2021-01-05
现代农村科技(2020年7期)2020-12-19
扬子江(2018年1期)2018-01-26
中国食用菌(2017年2期)2017-03-28
森林公安(2017年1期)2017-01-26
作文评点报·作文素材初中版(2016年5期)2016-03-11
语文世界(小学版)(2015年2期)2015-04-07
新诗(2014年1期)2014-08-15
