
银丝竹3种颜色叶片光合特性研究
2018-05-11 08:04:30赖金莉李欣欣陈凌艳荣俊冬郑郁善
生态环境学报 2018年2期
赖金莉,李欣欣,陈凌艳,荣俊冬,郑郁善, *
1.福建农林大学园林学院,福建 福州 350002;2.福建农林大学林学院,福建 福州 350002
银丝竹(Bambusa multiplexcv.Silverstrip)是禾本科(Gramineae)簕竹属(Bambusa)孝顺竹(Bambusa multiplex)的一个变种,与原变种主要区别在于其绿色的竿、箨鞘、叶片上间有黄白色纵条纹,而这种纵条纹的出现是由于银丝竹叶绿素生物合成受阻导致叶绿体无法正常发育,这种非正常发育的叶片异常美丽,为庭院中栽培的观赏品种,应用十分广泛,宜于庭园中孤植、群植,可作为划分空间的高篱或植于宽阔的庭园绿地(陈松河,2009;陈凌艳,2017)。银丝竹作为一种观赏价值极高的园林植物,具有很好的发展前景,但是目前有关银丝竹的研究却鲜有报道,其主要集中在对其抗寒性的研究方面(徐俊,2010;黄程前等,2013;滕召勇,2013),而对其不同叶色的光合生理研究尚未见报道。
在长期的外界选择压力下,植物叶片在进化过程中具有很强的变异性,为适应多变的外界环境(水分、温度和光照),易形成多种不同适应类型的叶片,充分体现着植物对环境变化的适应(Klich,2000)。植物生理生态特性与植物各器官特性有紧密联系,尤其是叶片的光合作用(程建峰等,2012)。光合作用是植物生长及其他代谢活动的生理基础,不同的光合特性往往是植物在生长过程产生差异的直接原因之一(张虎等,2017),其机制包括光合色素、光系统、电子传递系统和 CO2还原途径(Ashraf et al.,2013)。而植物在进行光合作用时,其光合色素对光能的吸收和利用起着重要的作用,在一定范围内,叶绿素含量越多,光合越强(潘瑞炽,2012)。已有的研究主要集中在光合特性与生态环境因子之间的关系研究(程业森等,2017;习玉森等,2016;徐祥增等,2017;李征珍等,2017;陈延松等,2017;李彩斌等,2017),极少涉及叶片自身特性与光合作用的关系研究(王海珍等,2014)。鉴于此,选择以银丝竹为试验材料,研究其全绿叶、花叶和全白叶3种不同颜色叶片与光合特性的关系,旨在了解银丝竹的光合生产力,并为其种植管理提供理论基础和参考依据。
1 试验材料与方法
1.1 试验材料
供试材料为3盆1年生银丝竹盆栽苗。试验地在福建农林大学竹类研究所苗圃内,地处经度119°23′E,纬度 26°10′N,四季常青,阳光充足,雨量充沛,霜少无雪,夏长冬短,气温适宜,温暖湿润,无霜期达326 d。年平均日照数为1700~1980 h;年平均降水量为 900~2100 mm;年平均气温为20~25 ℃,属典型的亚热带季风气候。盆栽苗购买于2017年5月12日并保持盆栽原状栽于福建农林大学竹类研究所苗圃内,正常供水1个月缓苗至定苗,于2017年6月15日开始进行试验。每盆银丝竹挑选中上部长势较好的全绿叶、花叶(深绿叶片间有白或黄绿色条纹的叶片)和全白叶(如图 1)进行实验。
1.2 测定方法
1.2.1 叶绿素含量的测定采用丙酮混合法提取
叶绿素含量测定参照李合生(2000)的方法,略有改动:分别取银丝竹全绿、花叶和全白叶新鲜叶片若干,洗净叶片表面的污渍,剪碎(去掉中脉),各称取0.25 g,每种叶色各称取3份,浸入25 mL丙酮与无水乙醇的混合液(体积比 1∶1)中浸提48 h至叶片完全变白,以丙酮和无水乙醇混合液为空白对照,在波长分别为470、663、645 nm处测定光密度(optical density,OD)值,每份样品测3次,取平均值,并计算叶绿素a、叶绿素b及类胡萝卜素含量。
1.2.2 光合光响应的测定
利用Li-6400便携式光合作用测定系统(Li-cor 6400,USA)测定光合-光响应曲线。先预实验确定银丝竹不同叶片的光饱和点,将待测叶片在光饱和点处对应的光强下诱导30 min,再利用Li-6400-02B LED红蓝光源提供不同强度的光合有效辐射(PAR,Photosynthetically active radiation),光合有效辐射设定为 0、20、40、60、80、100、125、150、200、400、600、800、1200、1600、1800、2000 μmol·m-2·s-1,测定时用 CO2钢瓶将参比室的 CO2浓度稳定在(400±0.5) μmol·mol-1,气体流速 Flow 值设为(500±0.5) mmol·s-1,设置每次改变光强后最短稳定时间为180 s,最长稳定时间为300 s。测定净光合速率 Pn(μmol·m-2·s-1)、气孔导度 Gs(μmol·mol-1)、胞间 CO2浓度 Ci(μmol·mol-1)、蒸腾速率 Tr(μmol·m-2·s-1),并计算植物水分利用瞬时效率WUE=Pn/Tr。
1.2.3 CO2响应曲线的测定
利用Li-6400-02B LED红蓝光源控制光强在光饱和点处,安装CO2钢瓶注入系统提供CO2,控制CO2浓度在 0~1800 μmol·mol-1,设置梯度为 0、50、100、150、200、300、400、600、800、1000、1200、1400、1600、1800 μmol·mol-1,气体流速 Flow 值设为(500±0.5) mmol·s-1,设置每次改变光强后最短稳定时间为180 s,最长稳定时间为300 s。

图1 银丝竹3种不同颜色叶片Fig.1 Three different color leaves of B. multiplex cv silverstripe
1.3 数据处理
利用 Excel 2007对试验数据进行整理,采用SPSS 19.0软件进行单因素方差分析并对银丝竹不同叶色的光合参数进行了差异性分析。利用Origin 8.5软件绘制了银丝竹的光响应和CO2响应曲线。

式(1)中,Pn为净光合速率;α是光响应曲线的初始斜率;β为修正系数;γ为一个与光强无关的系数;Rd为暗呼吸;
式(2)中,Ci为胞间CO2浓度;a是CO2响应曲线的初始羧化效率;b和c为系数(单位为mol·μmol-1);Rp为光呼吸速率,由于光下暗呼吸很弱,可以将光下叶片向空气中释放CO2的速率近似看作光呼吸速率(蔡时青等,2000)。
2 结果与分析
2.1 光合色素含量测定结果
由表1可知,银丝竹全绿叶、花叶和全白叶之间光合色素含量差异显著,并且全绿叶的叶绿素 a(Chl a)、叶绿素b(Chl b)、总叶绿素(Chl a+b)和类胡萝卜素(Car)含量显著高于花叶和全白叶(P<0.05),全白叶的Chl a、Chl b、Chl a+b和Car含量显著低于全绿叶和花叶(P<0.05),而全绿叶各色素含量分别比花叶高36.84%、58.78%、42.47%、22.75%,全绿叶分别比全白叶高98.45%、98.45%、98.45%、98.36%,而3种叶片的叶绿素a/b(Chl a/b)和总叶绿素/类胡萝卜素(Chl a+b/Car)的比值差异不显著(P>0.05)。

表1 银丝竹不同颜色叶片光合色素含量的变化Table1 Variation of photosynthetic pigment content of B. multiplex cv. Silverstrip
2.2 光响应曲线及参数测定结果
光响应曲线反映了植物的光合速率随光合有效辐射的变化规律。如图2所示,银丝竹全绿叶和花叶的Pn随着PAR的增大,由负值慢慢变大直至最大值,随后,Pn呈逐渐下降趋势,即在达到光饱和点(LSP)后不再上升,甚至出现下降的趋势,此时出现了光抑制现象。全绿叶片在低光强(PAR≤200 μmol·m-2·s-1)下存在一个快速光响应阶段,即 Pn随着PAR的增大而迅速上升;而花叶的Pn较低,整体上随PAR变化而变化的幅度较平缓。
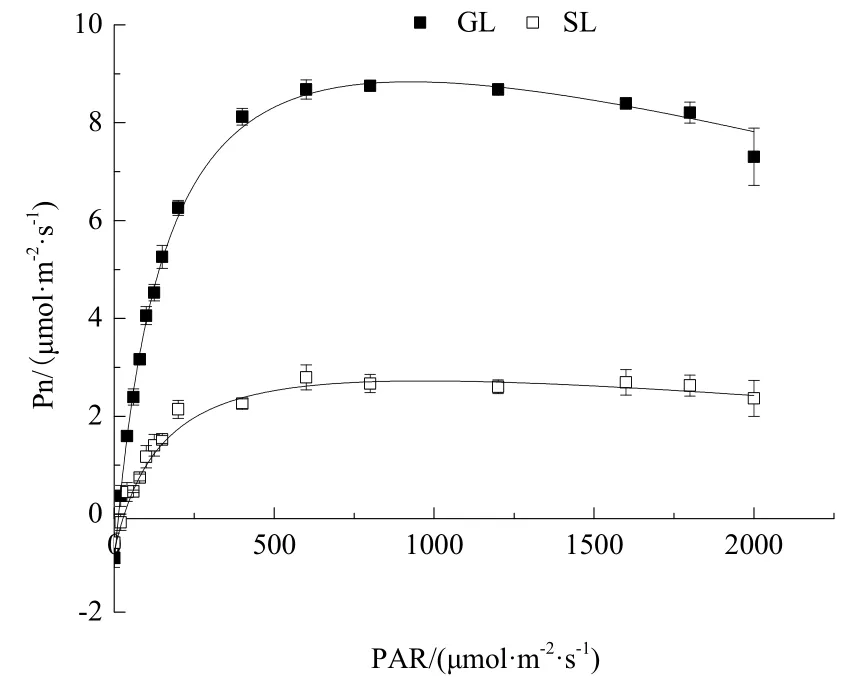
图2 银丝竹不同颜色叶片的光响应曲线Fig.2 Light response curves among different leaf color
光响应曲线拟合结果显示(表2),银丝竹全绿叶的初始量子效率(α)、最大净光合速率(Pnmax)和暗呼吸速率(Rd)分别为0.077、8.960和0.957,均高于花叶,且全绿叶与花叶初始量子效率(α)、最大净光合速率(Pnmax)存在显著差异(P<0.05),而暗呼吸速率(Rd)差异性不显著(P>0.05)。花叶的LSP和光补偿点(LCP)高于全绿叶,但是差异不显著(P>0.05)。全白叶片测不出光合-光响应曲线,故不予分析。
2.3 气体交换参数测定结果
图3表明,随着光强的增加,银丝竹叶片的Gs逐渐增大,从而引起Tr的增大,但Ci随着气孔张开以及光合速率的加快而迅速下降,在PAR≥500 μmol·m-2·s-1时趋于平稳,而 WUE 的变化趋势和 Ci刚好相反,随着PAR的增大,WUE逐渐增大直至平稳甚至呈下降趋势。其中,Gs、Tr和WUE均表现为全绿叶>花叶>全白叶,而3种叶片的Ci的大小关系则刚好相反。
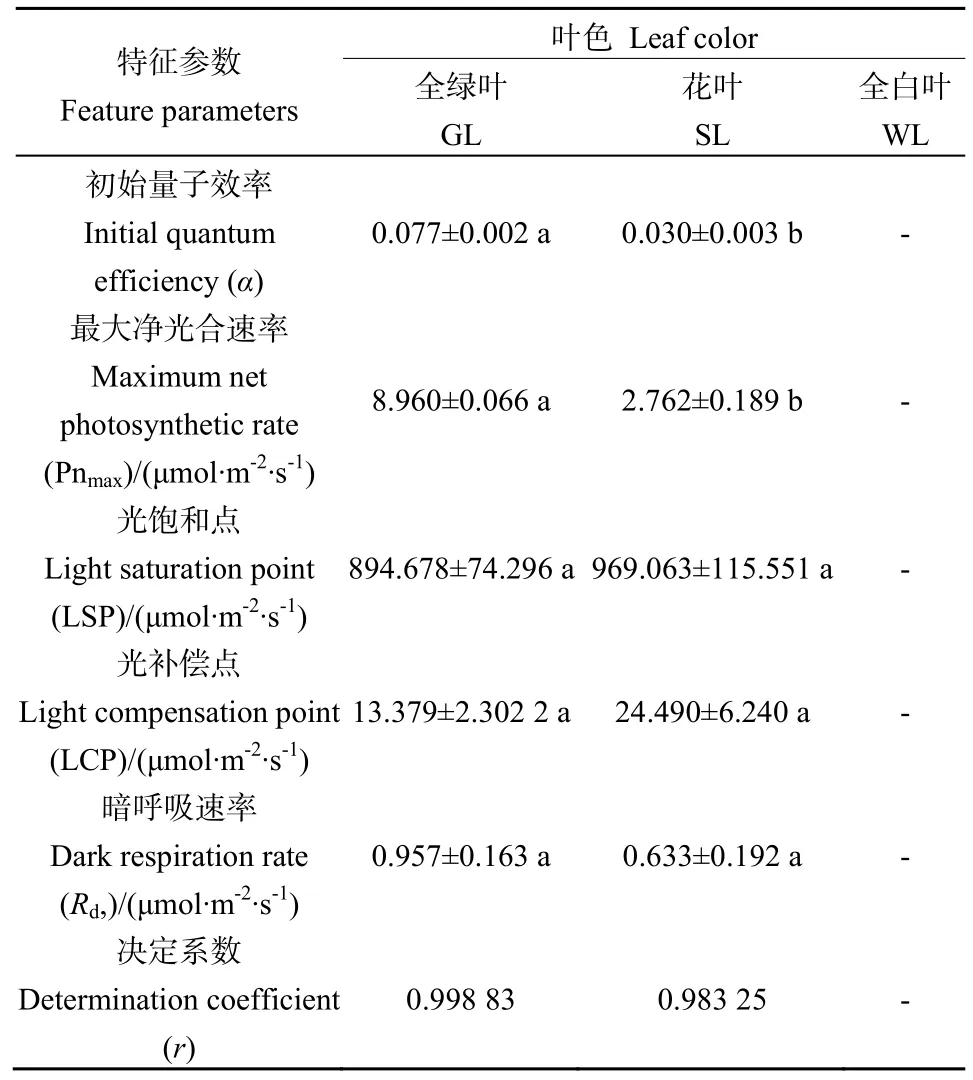
表2 不同颜色叶片光合-光响应曲线特征参数Table2 Feature parameters of light response for different leaf color
2.4 CO2响应曲线测定结果
CO2响应曲线反映了植物光合速率随着 CO2浓度变化的规律。银丝竹全绿叶和花叶的Pn值都随着CO2浓度的增加而增大并达到最大值(图4),Pn都在低 CO2浓度下快速提高,而后缓慢上升。全绿叶Pn在整个过程中的增幅较大,而花叶的变化幅度则较小。
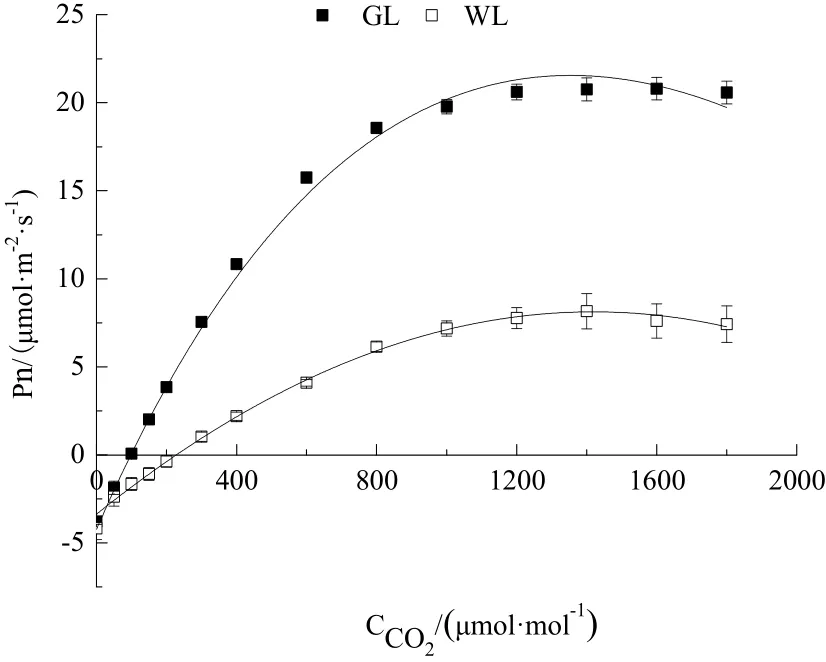
图4 银丝竹不同颜色叶片净光合速率对CO2浓度的响应Fig.4 Response of net photosynthetic rate of different leaf color of B. multiplex cv. Silverstripto air CO2 concentration

图3 银丝竹不同颜色叶片的气孔导度、胞间CO2、蒸腾速率及水分利用效率对光强的响应Fig.3 Response of Gs, Ci, Tr and WUE to light in different leaf color

表3 不同颜色叶片CO2响应曲线模拟的光合参数Table3 Photosynthesis parameters of CO2 response curve for different leaf color
根据拟合结果(表3),银丝竹全绿叶和花叶的初始羧化效率、光合能力和CO2补偿点存在显著差异(P<0.05),CO2饱和点和光呼吸速率差异不显著(P>0.05)。在银丝竹不同颜色的叶片中,全绿叶片具有较高的 CO2饱和点(1518.864 μmol·mol-1),但其 CO2补偿点却较低(90.562 μmol·mol-1);花叶片具有较高的 CO2补偿点(223.651 μmol·mol-1)和较低的 CO2饱和点(1420.903 μmol·mol-1)。全白叶片测不出CO2响应曲线,故不予分析。
3 讨论
许庆全(2017)等在研究墨兰(Cymbidium sinense)“达摩”叶艺品系时发现,叶片黄色区域中叶绿素 a、叶绿素 b、总叶绿素、类胡萝卜素含量均大幅减少,总叶绿素含量减少导致叶片黄化。在本研究中,银丝竹的叶绿素 a、叶绿素 b、总叶绿素、类胡萝卜素含量表现为:全绿叶>花叶>全白叶,这与陈凌艳等(2017)对银丝竹的测定结果类似。光合色素(如叶绿素和类胡萝卜素)可以利用太阳能固定二氧化碳,在植物光合作用的光反应中起关键作用(Singh et al.,2016),其中叶绿素含量越高,越有利于植物在低光照条件下有效吸收光能,有利于光合作用的进行,增加有机物的积累(吴志庄等,2017),而叶绿素含量的减少会导致叶片衰老和光合速率的不可逆降低(杨富军等,2013),同时,叶片中叶绿素含量的变化也会改变叶片的颜色(Li et al.,2017)。本研究中,银丝竹全白叶叶绿素含量显著低于全绿叶和花叶,由此可以推测银丝竹全白叶的存在是由于叶片中叶绿素含量太少而导致,这与王啸晨等(2012)对菲黄竹(Sasa auricoma)的研究结果类似,从而进一步导致其光合作用减弱。
光合作用是评价植物生长状况的重要指标,因为它与植物的光合生产力直接相关(Jumrani et al.,2017)。光响应参数能较好地反映植物的光合能力、光能利用率及光抑制水平等特性(王海珍等,2014)。银丝竹的全绿叶和花叶的Pn随着PAR的增大而增加,当PAR超过某一数值后,Pn便不再增加且有下降的趋势,此时便出现了光饱和现象。银丝竹全绿叶和花叶对高光强的反应不一样,全绿叶在高光强下仍然保持较高的净光合速率,且在相同的PAR下,全绿叶的Pn明显高于花叶,可以说明全绿叶受到的光抑制较小,光合效率更高。LCP、LSP、Pnmax和α是反映植物光响应特征的重要参数(Xia et al.,2011),其中,LCP与LSP反映了植物对光照条件的适应性(Deng et al.,2003),Pnmax和α则是表征植物光合作用产生光抑制的特征参数(王海珍等,2014)。银丝竹全绿叶的Pnmax和α显著高于花叶,而LCP和LSP却明显低于花叶,表明银丝全绿叶光合活性高于花叶,且受光抑制的程度也较小,通过提高Pnmax增强叶片的光能利用率,使银丝竹在南方地区夏秋季高温、强光的环境下仍能保持较高的光合活性。提高银丝竹对干物质的积累和物质产量的形成以及增强其对逆境环境的抵抗能力,有利于其在南方夏秋季高温干旱的环境中正常生长。LCP能够反映植物对弱光的适应能力,LSP能够反映植物对强光的适应能力(李迎春等,2011;吴志庄等,2013),银丝竹全绿叶片的 LCP较高,而花叶的LSP较高,因此花叶利用强光的能力比全绿叶片高,而利用弱光的能力较差,这也是银丝竹花叶在夏季多于其他两种叶色,而在春季少于全绿叶的原因,这与孙艳(2014)对竹种的生态适应性的研究结果一致:银丝竹叶绿素含量在春季最高,在夏季最低,而叶绿素的含量决定了叶片的颜色(潘瑞炽,2012)。
植物气孔作为光合作用和呼吸作用与外界气体交换的通道,影响着植物的蒸腾、光合和呼吸等一系列过程。本试验中,银丝竹的3种叶色的叶片Gs都随着PAR的增加而增大,在低PAR条件下,其增幅较大,表明银丝竹3种叶色的叶片在弱光环境下随着PAR的增加,将作为光合反应底物的CO2迅速消耗,这也是银丝竹胞间CO2先在低光照强度下迅速下降的原因。此时植物为了补充反应底物进而提高植物叶片与大气间的气体交换速率,加快了植物的蒸腾速率,此时水分利用效率也增加。
植物碳同化受核酮糖-1, 5-二磷酸羧化酶(Rubisco)活性和1, 5-二磷酸核酮糖(RuBP)再生速率的限制,而 1, 5-二磷酸核酮糖(RuBP)再生与光合电子传递速率直接关联(黄红英等,2009;王海珍等,2014)。一般认为 CO2响应曲线中初始羧化效率(CE)与核酮糖-1, 5-二磷酸羧化酶(Rubisco)活性呈正相关(Deng et al.,2003)。叶片的羧化效率(CE)越高,说明光合作用对 CO2的利用越充分(董晓颖等,2005)。本试验中,全绿叶片的初始羧化效率(CE))显著高于花叶,说明全绿叶片具有较强的光合电子传递能力和核酮糖-1, 5-二磷酸羧化酶(Rubisco)活性,故对 CO2的利用较花叶更充分,可以推测全绿叶片光合活性更高与充分利用CO2有光。本研究只是从光合生理层面对银丝竹不同叶色作了对比,陈凌艳等(2017)已经从叶绿体结构差异上研究了银丝竹叶片变色的机理,今后的研究方向可以转向转录组甚至基因来探索其叶片变色的机理。
4 结论
较花叶而言,全绿叶无论是从光响应曲线还是CO2响应曲线上看,都具有更高的光合能力,这跟全绿叶中叶绿素含量高有一定的关系。就园林观赏竹类而言,叶片大部分都是绿色的,而花叶品种较少,本研究中发现花叶利用强光的能力比全绿叶高,故在人工温室育苗过程中可以通过提高光强来培育更多的银丝竹花叶品种。
参考文献:
ASHRAF M, HARRIS P J C. 2013. Photosynthesis under stressful environments: an overview [J]. Photosynthetica, 51(2): 163-190.
DENG X, LI X M, ZHANG X M, et al. 2003. The studies about the photosynthetic response of the four desert plants [J]. Acta Ecologica Sinica, 23(3): 598-605.
JUMRANI K, BHATIA V S, PANDEY G P. 2017. Impact of elevated temperatures on specific leaf weight, stomatal density, photosynthesis and chlorophyll fluorescence in soybean [J]. Photosynthesis Research,131(3): 333-350.
KLICH M G. 2000. Leaf variations in elaeagnus angustifolia, related to environmental heterogeneity [J]. Environmental & Experimental Botany, 44(3): 171-183.
LI X, KANAKALA S, HE Y H, et al. 2017. Physiological characterization and comparative transcriptome analysis of white and green leaves ofananascomosusvar.bracteatus[J]. Plos One,DOI:10.1371/journal.pone.0169838.
SINGH S K, REDDY V R, FLEISHER D H, et al. 2016. Relationship between photosynthetic pigments and chlorophyll fluorescence in soybean under varying phosphorus nutrition at ambient and elevated CO2[J]. Photosynthetica, 55(3): 1-13.
XIA J B, ZHANG S Y, ZHANG G C, et al. 2011. Critical responses of photosynthetic efficiency inCampsis radicans(L.) seem to soil water and light intensities [J]. African Journal of Biotechnology, 10(77):17748-17754.
蔡时青, 许大全. 2000. 大豆叶片 CO2补偿点和光呼吸的关系[J]. 植物生理学报, 26(6): 545-550.
陈凌艳, 何丽婷, 赖金莉,等. 2017. 银丝竹不同叶色叶绿素合成及叶结构差异[J]. 森林与环境学报, 37(4):385-391.
陈松河. 2009. 观赏竹园林景观应用[M]. 北京: 中国建筑工业出版社:83-84.
陈延松, 袁华玲, 卫文渊, 等. 2017. 夏季遮阳对‘红阳’猕猴桃净光合速率的影响及其与生理生态因子的关系[J]. 果树学报, 34(9):1144-1151.
程建峰, 陈根云, 沈允钢. 2012. 植物叶片特征与光合性能的关系[J]. 中国生态农业学报, 20(4): 466-473.
程业森, 赵晨光, 高立平. 2017. 阿拉善白刺生长季光合速率及蒸腾速率特征研究[J]. 干旱区资源与环境, 31(11): 161-168.
储钟稀, 童哲, 冯丽洁, 等. 1999. 不同光质对黄瓜叶片光合特性的影响[J]. 植物学报, 41(8): 867-870.
董晓颖, 李培环, 王永章, 等. 2005. 水分胁迫对不同生长类型桃叶片水分利用效率和羧化效率的影响[J]. 灌溉排水学报, 24(5): 67-69.
杜鹏珍, 廖绍波, 孙冰, 等. 2014. 班克木幼苗的光合色素及叶绿素荧光特性[J]. 中南林业科技大学学报, 34(9): 49-54.
黄程前, 黄滔, 刘玮, 等. 2013. 20种观赏丛生竹的抗寒性测定[J]. 湖南城市学院学报(自然科学版), 22(2): 59-62.
黄红英, 窦新永, 孙蓓育, 等. 2009. 两种不同生态型麻疯树夏季光合特性的比较[J]. 生态学报, 29(6): 2861-2867.
李彩斌, 郭华春. 2017. 耐弱光基因型马铃薯在遮阴条件下的光合和荧光特性分析[J]. 中国生态农业学报, 25(8): 1181-1189.
李合生. 2000. 植物生理生化实验原理和技术[M]. 北京: 高等教育出版社.
李迎春, 杨清平, 陈双林, 等. 2011. 龟甲竹光合生理特性及其与主要影响因子关系. 林业工程学报, 25(2), 35-39.
李征珍, 杨琼, 石莎, 冯金朝. 2017. 蒙古沙冬青光合作用特征及其影响因素[J]. 生态学杂志, 36(9): 2481-2488.
刘建锋, 杨文娟, 史胜青, 等. 2011. 崖柏与侧柏光合特性和叶绿素光参数的比较研究[J]. 西北植物学报, 31(10): 2071-2077.
马姜明, 黄婧, 杨栋林, 等. 2015. 桂林喀斯特石山 50种常见植物叶片光合色素含量及耐荫性定量评价[J]. 林业科学, 51(10): 67-74.
潘瑞炽. 2012. 植物生理学[M]. 第7版. 北京: 高等教育出版社.
孙艳. 2014. 百竹园26种竹种生态适应性初步研究[D]. 福州: 福建农林大学.
滕召勇. 2013. 湖南丛生观赏竹抗寒性研究[D]. 长沙:湖南农业大学.
王海珍, 韩路, 徐雅丽, 等. 2014. 胡杨异形叶光合作用对光强与CO2浓度的响应[J]. 植物生态学报, 38(10): 1099-1109.
王啸晨, 张磊, 岳祥华, 等. 2012. 菲黄竹叶片色素含量、结构与颜色之间关系初探[J]. 世界竹藤通讯, 10(6): 5-9.
吴志庄, 杜旭华, 熊德礼, 等. 2013. 不同类型竹种光合特性的比较研究[J]. 生态环境学报, 22(9): 1523-1527.
吴志庄, 高贵宾, 欧建德, 等. 2017. 生物炭肥对毛竹林下三叶青叶绿素含量、光合与荧光特性的影响[J]. 西北林学院学报, 32(5): 59-63,103.
习玉森, 姜卫兵, 文杨, 等. 2016. 不同生长型桃树光合效能及其季节变化特征初探[J]. 西北植物学报, 36(9): 1836-1845.
徐俊. 2010. 银丝竹等四种城市观赏竹抗寒性生理研究[D]. 成都: 四川农业大学.
徐祥增, 张金燕, 张广辉, 等. 2017. 环境光强对三七光合能力及能量分配的影响[J]. 应用生态学报, DOI: 10.13287/j.1001–9332.201801.008.
许庆全, 杨凤玺, 叶庆生, 等. 2017. 墨兰‘达摩’叶艺品系光合色素含量、叶绿素荧光特性和叶绿体超微结构的比较[J]. 热带作物学报,38(7): 1210-1215.
杨富军, 赵长星, 闫萌萌, 等. 2013. 栽培方式对夏直播花生叶片光合特性及产量的影响[J]. 应用生态学报, 24(3): 747-752.
叶子飘, 于强. 2007. 一个光合作用光响应新模型与传统模型的比较[J].沈阳农业大学学报, 38(6): 771-775.
叶子飘, 于强. 2008. 光合作用光响应模型的比较[J]. 植物生态学报,32(6): 1356-1361.
叶子飘, 于强. 2009. 光合作用对胞间和大气 CO2响应曲线的比较[J].生态学杂志, 28(11): 2233-2238.
张虎, 姜文龙, 武启飞, 等. 淹水胁迫对湖北海棠生长及光合作用的影响[J]. 东北林业大学学报, DOI: 10.13759/j.cnki.dlxb.2017.12.005.
猜你喜欢
中国音乐学(2022年2期)2022-08-10 09:20:54
读者·校园版(2019年20期)2019-10-18 01:24:32
知识经济·中国直销(2018年11期)2018-11-26 01:18:20
音乐研究(2018年5期)2018-11-22 06:43:54
少年漫画(艺术创想)(2018年7期)2018-11-19 03:00:26
知识经济·中国直销(2018年8期)2018-08-23 09:16:00
知识经济·中国直销(2018年7期)2018-07-27 02:49:44
现代园艺(2018年3期)2018-02-10 05:18:20
知识经济·中国直销(2017年7期)2017-07-24 14:12:44
快乐语文(2017年9期)2017-06-15 20:28:36
