
内蒙古短花针茅荒漠草原土壤线虫群落的垂直分布特征
2018-05-07 03:50贾美清韩国栋杨向芸张国刚
天津师范大学学报(自然科学版) 2018年2期
孟 元 ,贾美清 ,林 宇 ,韩国栋 ,黄 静,隋 鑫 ,尹 雪 ,杨向芸 ,张国刚 ,5
(1.天津师范大学生命科学学院,天津300387;2.天津市水资源与水环境重点实验室,天津300387;3.天津出入境检验检疫局动植物与食品检测中心,天津300461;4.内蒙古农业大学草原与资源环境学院,呼和浩特010019;5.天津师范大学天津市动植物抗性重点实验室,天津300387)
土壤线虫是土壤动物的重要组成部分,其种类和数量丰富,食性多种多样,占据食物网中的多个营养级,直接参与生态系统的物质循环和能量流动,易于分离,许多种类能在极端条件下存活并能对环境变化作出反应[1-2].因此,线虫作为指示生物用来评估土壤的健康水平越来越受关注[3].
荒漠草原位于草原和荒漠两大陆地生态系统的中间地带[5],我国内蒙古荒漠草原约占内蒙古草原总面积的10.7%.由于植物稀少,气候干旱,土壤养分状况较差,加上人为因素干扰,导致荒漠化日益严重.与其他草原类型相比,荒漠草原稳定性较差,对外界环境和人为因素的干扰比较敏感.近年来,由于畜牧业的快速发展,荒漠草原食草动物数量大幅度增加,导致植物种类减少,植被盖度降低,草原荒漠化加剧.目前对荒漠草原生态系统的研究主要集中在气候变化、放牧方式等对草原的群落结构、植被空间异质性及土壤微生物的影响等方面[4-5],关于荒漠草原土壤线虫对环境变化响应的研究相对较少.本研究对内蒙古短花针茅荒漠草原土壤线虫的群落组成和多样性进行调查,旨在揭示土壤线虫在荒漠草原生态系统中的地位和作用,为荒漠草原的恢复和利用提供理论参考.
1 材料与方法
1.1 研究区域概况
样地位于内蒙古自治区乌兰察布市四子王旗农牧科学院长期定位试验站(N41°47′17″,E111°53′46″),海拔高度为1 456 m,平均气温为3.4℃,样地内的放牧为开放自由式.植被类型为典型荒漠草原,植物组成匮乏,建群种为短花针茅(Stipa breviflora),优势种为冷蒿(ArtemisiafrigidaWilld)和无芒隐子草(Cleistogenes songorica),伴生种主要有木地肤(Kochia prostrata)、阿氏旋花(Apgarbindweed)、细叶葱(Alliumtenuissimum)、羊草(Leymus chinensis)等.
1.2 研究方法
1.2.1 土壤样品采集
2016年8月初,在试验区内选择3块典型的短花针茅荒漠草原样地,在每块样地内分别用土壤环刀采集 0~5、5~10、10~20、20~30 cm 的 4 层土壤,每层 3 次重复.为了减少土壤线虫空间分布差异所造成的影响,每次重复对样地内进行多点取样,每个取样点地上植被类型必须为典型短花针茅荒漠草原群落.将土样装入密封袋中带回实验室内,放入4℃冰箱中备用.
1.2.2 土壤线虫分离和鉴定
漏斗中铁丝网上放入纱布包裹好的土壤,漏斗上端设置有60 W的白炽灯,下端装有带止水夹的乳胶管,加入蒸馏水浸没土壤,20℃条件下分离,期间不断加入蒸馏水.24 h后将液体收集到离心管中,65℃水浴加热30 min,加入等体积的TAF固定液,保存待用.
土壤线虫鉴定:根据文献[6]在显微镜和体式显微镜等仪器下对线虫进行分类鉴定.
1.2.3 数据处理方法
本研究以每100g干土中土壤线虫的数量对线虫进行计数.基于不同类群的数量百分比划分优势:个体数占总数10%以上的为优势种;占总数1%~10%的为常见种;占总数1%以下的为罕见种.根据形态特征和取食特性将土壤线虫划分为4个功能类群:植食性类群(phytophagous nematode)、食细菌类群(bacterivorous nematode)、食真菌类群(fungi-feeding nematode)和捕食杂食类群(omni-vorous nematode).采用如下参数计算土壤线虫的群落多样性:
Shannon-Wiener多样性指数(H):H=-∑Pi× lnPi
Pielou均匀度指数(J):J=H/lnS
Margalef丰富度指数(SR):SR=(S-1)/lnN植物寄生线虫指数(PPI):PPI= ∑vi× fi
自由生线虫成熟度指数(MI):MI= ∑vi× fi′
式中:S为物种数;Pi=Ni/N,为各个种群的个体数量与群落总个体数量的比值;vi为土壤线虫第i类群的c-p值;fi为植物寄生线虫第i类群的个体占群落总个体数的比例;fi′为自由生线虫第i类群的个体占群落总个体数的比例.
2 结果与分析
2.1 土壤线虫的数量变化
土壤线虫在不同土层的数量分布如图1所示.
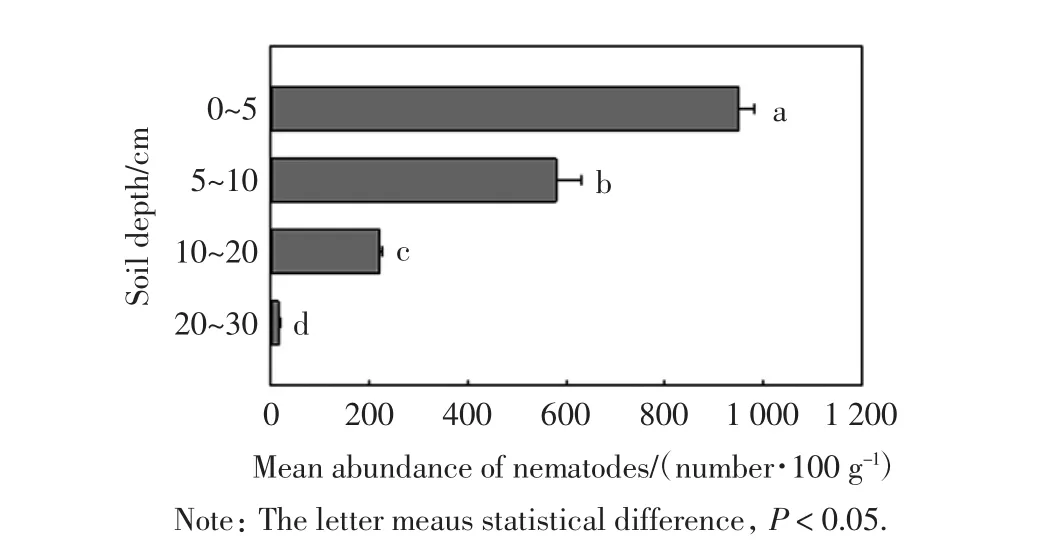
图1 不同土壤深度线虫的数量Fig.1 Number of nematodes at different soil depth
由图1可以看出,不同土层间的土壤线虫数量差异均具有统计学意义(P<0.05).土壤线虫数量的垂直分布呈现明显的表聚型,即线虫主要集中在0~5 cm层,该层土壤线虫数量占总数的53.78%,表明0~5 cm层土壤环境更适合土壤线虫生存.随着土层深度的增加,土壤线虫数量逐渐减少,20~30 cm层土壤线虫数量最少,仅占总数的0.91%.
2.2 土壤线虫不同功能类群的数量变化
土壤线虫4个功能类群在不同土壤深度下的数量及比例如图2所示.
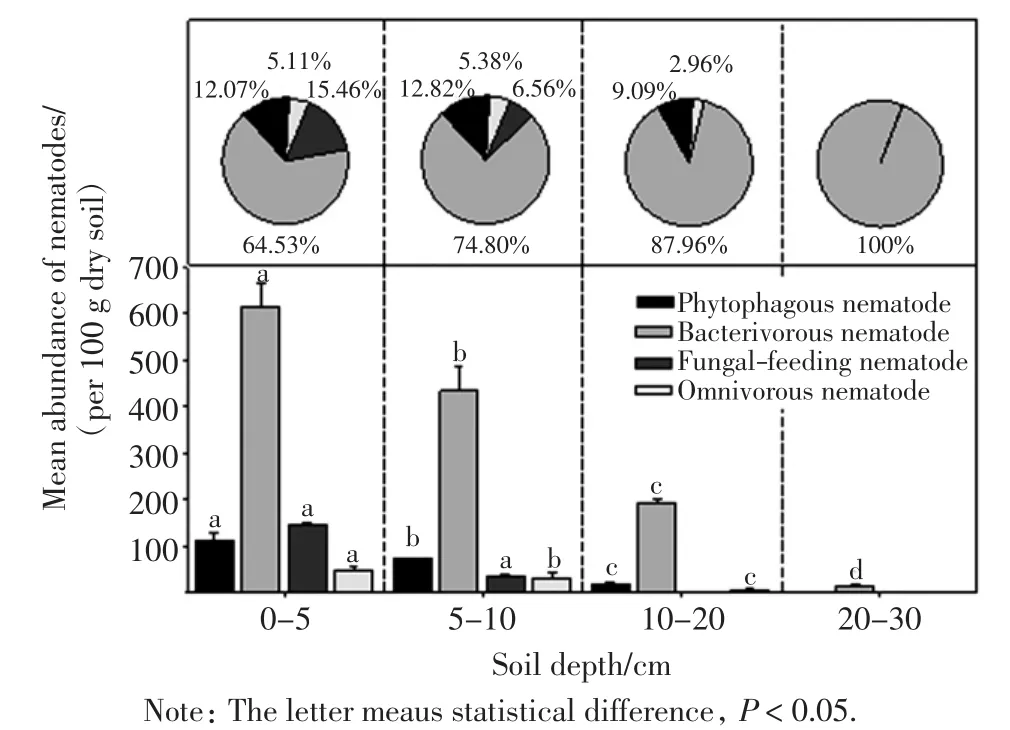
图2 不同土壤深度线虫群落中各功能类群线虫的数量及比例Fig.2 Mean abundance and proportion of four trophic groups of the nematode communities at different soil depth
由图2可以看出,4个功能类群线虫的垂直分布均呈现表聚型.植食性线虫的数量在0~5 cm层最高,5~10 cm和10~20 cm层分别下降了35.29%和82.35%,20~30 cm层中未发现植食性线虫.食细菌线虫在0~5 cm层每100 g干土中的数量为615只,其他3个土层依次减少了29.28%、68.33%和97.40%.食真菌线虫仅出现在0~5 cm和5~10 cm土层.捕食杂食性线虫5~10 cm和10~20 cm层的数量分别比0~5 cm土层减少36.11%和86.11%,20~30 cm层中未发现捕食杂食性线虫.
在各层土壤线虫群落中,食细菌性线虫占绝对优势,所占比例随着土壤深度的增加而增加,在4个土层中的占比分别为64.53%、74.80%、87.96%和100%.植食性和食真菌类群数量相近,捕食杂食类群数量最少.
2.3 土壤线虫的群落组成及数量关系
在内蒙古短花针茅荒漠草原共分离鉴定出土壤线虫26个属,群落组成如表1所示.

表1 不同深度土壤线虫的群落组成Tab.1 Amount and type of nematodes at different soil depth
由表 1可知,丽突属(Acrobeles)和拟丽突属(Acrobeloides)为优势种,占线虫总数的65.48%,其中丽突属占46.75%,拟丽突属占18.73%.柄端球属(Paurodontus)、小杆属(Rhabditis)、鹿角唇属(Cervidellus)、矮化属(Tylenchorhynchus)、垫咽属(Tylencholaimus)、真滑刃属(Aphelenchus)、滑刃属(Aphelenchoides)、真矛线属(Eudorylaimus)、孔咽属(Aporcelaimus)、钵盘属(Mylodiscs)、单齿属(Mononchus)为常见属,占线虫总数的29%,其余属为罕见属.丽突属、拟丽突属在各个土层均有分布;板唇属(Chiloplacus)、裸矛属(Psilenchus)、大矛属(macrodora)、桑尼属(Thornia)、狭咽属(Discolaimium)只出现在 0~5 cm 土层;真头叶属(Eucephalobus)、螺旋属(Helicotylenchus)、短体属(Pratylenchus)、头垫刃属(Tetylenchus)、伪垫刃属(Nothotylenchus)、轮属(Criconrmoides)、垫咽属、真滑刃属、滑刃属、真矛线属、拟桑尼属(Thorneella)、钵盘属、单齿属在10~20 cm层开始消失;在20~30 cm层只发现了丽突属和拟丽突属;散香属(Beleodorus)只出现在5~10 cm层,孔咽属出现在5~10 cm层和10~20 cm层.随着土壤深度的增加,土壤线虫数量逐渐减少,绝大多数土壤线虫的垂直分布呈表聚型.
2.4 土壤线虫群落多样性及成熟度指数
内蒙古短花针茅荒漠草原不同深度土壤线虫群落的多样性特征如表2所示.

表2 不同深度土壤线虫群落的多样性Tab.2 Diversity of the nematode communities at different soil depth
由表2可以看出,随着土层深度的增加,土壤线虫群落的多度(Abundance)呈下降趋势,各层之间的差异均具有统计学意义(P<0.05),多度的垂直分布呈现明显的表聚型.线虫多样性指数(H)随着土层深度的增加而逐渐下降,各层之间的差异具有统计学意义(P<0.05).均匀度指数(J)在20~30 cm层的数值最大,显著高于其他土层,其他土层的线虫均匀度指数相近.丰富度指数(SR)在0~5 cm层数值最大,略高于5~10 cm层,显著高于10~20 cm和20~30 cm层(P<0.05).植物寄生线虫指数(PPI)在 0~5、5~10 和 10~20 cm层变化不显著,5~10 cm层的植物寄生线虫指数略高.自由生线虫指数(MI)是用于评价扰动程度的指标,低MI值表明系统扰动强烈,反之则表明系统稳定,自由生线虫指数在0~5 cm层的数值显著高于5~10 cm层和10~20 cm层,与20~30 cm层的差异不显著,说明0~5 cm和20~30 cm层抗干扰能力强,系统相对稳定.
3 讨论
在内蒙古短花针茅荒漠草原共发现土壤线虫26个属,丽突属和拟丽突属是该地区的优势种.土壤线虫种群密度为每100 g干土中有1 765条,与以往研究相近(每100 g干土中有504~3 121条)[7].内蒙古荒漠草原与其他3种草原类型相比土壤线虫数量与种类较少,这与张瑞易的研究结果一致[8].这可能是因为该地区植物种类和数量稀少,气候干旱,土壤养分状况较差.土壤线虫是典型的湿生动物,土壤湿度也是影响线虫数量的重要原因.内蒙古短花针茅荒漠草原土壤线虫总数随着土壤深度的增加呈现下降趋势,垂直分布特征为表聚型[9],0~5 cm层线虫的数量最大,这与Liu等[10]的研究结果一致.这主要是因为0~5 cm土层的土壤比较疏松且营养丰富,适宜土壤线虫的生存[11].不同深度土壤线虫数量的变化可能与其食物资源有关[12],随着土壤深度的增加,线虫的食物资源越来越匮乏,导致土壤线虫的数量减少.本研究发现内蒙古短花针茅荒漠草原的土壤线虫群落中食细菌线虫数量最多,而且食细菌线虫数量在各个土层之间的差异性显著,这与Zhang等[13]对沙丘土壤线虫的研究结果一致.由于本研究区域为较干旱的内蒙古短花针茅荒漠草原,较高的土壤有机质含量导致细菌资源丰富,为食细菌线虫提供了充足的营养物质,因此食细菌线虫数量较多[14].而大多数植食性线虫存活于根际土壤中,主要取食土壤中的植物根系,而荒漠草原中植物种类及数量相对较少,因此植食性线虫食物来源较少,导致植食性线虫的种类和数量减少.自由生线虫指数MI可以用来反映土壤生态系统受干扰的程度[15].本研究发现,短花针茅荒漠草原的MI指数变化范围为1.81~2.00,低于Zhang等[16]报道的2.58~2.88.随着土层深度的增加,自由生线虫指数MI呈现先下降后上升的趋势,表明内蒙古短花针茅荒漠草原土壤表层和深层的线虫群落具有较强的抗干扰能力.
深入了解线虫生物多样性和土壤生态系统功能可以更好地发挥土壤线虫在草原生态系统的生物指示作用.在以土壤线虫作为指示生物的研究中,可以通过了解线虫群落和优势种对环境变化作出的响应来评价生态系统的土壤健康水平.这种指示作用需要对线虫进行种类鉴定[17],依据线虫形态特征的传统鉴定方法对鉴定者要求极高[18],随着分子生物技术的发展,越来越多的学者开始尝试利用分子方法评价线虫多样性[19].本研究对内蒙古短花针茅荒漠草原土壤线虫群落进行了形态学研究,为进一步建立土壤线虫形态学鉴定与其分子身份的对应关系提供理论依据.
参考文献:
[1]PEN-MOURATOV S,HU C,HINDIN E,et al.Effect of sand-dune slope orientation on soil free-living nematode abundance and diversity[J].Helminthologia,2010,47(3):179-188.
[2]GOMES G S,HUANG S P,CARES J E.Nematode community,trophic structure and population fluctuation in soybean fields[J].Tropical Plant Pathology,2003,28(3):258-266.
[3]NEHER D A.Role of nematodes in soil health and their use as indicators[J].Journal of Nematology,2001,33(4):161-168.
[4]HAN F,ZHANG Q,BUYANTUEV A,et al.Effects of climate change on phenology and primary productivity in the desert steppe of Inner Mongolia[J].Journal of Arid Land,2015,7(2):251-263.
[5]黄维,贾美清,许帅,等.短花针茅荒漠草原土壤可培养真菌种群结构及其对放牧强度的响应[J].天津师范大学学报(自然科学版),2017,37(1):51-56.HUANG W,JIA M Q,XU S,et al.Population structure of cultivable soil fungi and its response to grazing intensities in Stipa breviflora desert steppe[J].Journal of Tianjin Normal University(Natural Science Edition),2017,37(1):51-56(in Chinese).
[6]谢辉.植物线虫分类学[M].北京:高等教育出版社,2005.XIE H.Tavanomy of Plant Nematodes[M].Beijing:Higher Education Press,2015(in Chinese).
[7]薛会英,胡锋,罗大庆.藏北高寒草甸植物群落对土壤线虫群落功能结构的影响[J].生态学报,2013,33(5):1482-1494.XUE H Y,HU F,LUO D Q.Effects of alpine meadow plant communities on soil nematode functional structure in Northern Tibet,China[J].ActaEcologicaSinica,2013,33(5):1482-1494(inChinese).
[8]张瑞益.北方主要草地类型土壤线虫群落结构及多样性的比较研究[D].长春:东北师范大学,2015.ZHANG R Y.Comparison of Soil Nematode Communities Structure and DiversityAcrosstheMain Grasslands in Northern China[D].Changchun:Northeast Normal University,2015(in Chinese).
[9]WESEMALE W M L,MOENS M.Vertical distribution of the plantparasitic nematode,meloidogyne chitwoodi,under field crops[J].Europ J Plant Pathol,2008,120:249-257.
[10]LIU Y,HUA J,JIANG Y,et al.Nematode communities in greenhouse soil of different ages from Shenyang suburb[J].Helminthologia,2006,43(1):51-55.
[11]VIKETOFTM,BENGTSSONJ,SOHLENIUSB,etal.Long-termeffects of plant diversity and composition on soil nematode communities in model grasslands[J].Ecology,2009,90(1):90-99.
[12]FERRIS H,MCKENRY M V.Nematode community structure in a vineyard soil[J].Journal of Nematology,1976,8(2):131-137.
[13]ZHANG X K,LIANG W J,JIANG D M,et al.Soil nematode community structure in a Chinese sand dune system[J].Helminthologia,2007,44(4):204-209.
[14]阮维斌,吴建波,张欣,等.内蒙古中东部大针茅群落土壤线虫多样性研究[J].应用与环境生物学报,2007,13(3):333-337.RUAN W B,WU J B,ZHANG X,et al.Soil nematode diversity in Stipa grandis community in the mid-east of Inner Mongolia[J].Chinese Journal of Applied and Environmental Biology,2007,13(3):333-337(in Chinese).
[15]PENMOURATOV S,HU C,HINDIN E,et al.Effect of sand-dune slope orientation on soil free-living nematode abundance and diversity[J].Helminthollogia,2010,47(3):179-188.
[16]ZHANG W D,XIAO Y,WANG X F,et al.Soil nematode community characteristics around the Gangue hill of Fushun West Open-pit mine[J].Helminthollogia,2011,48(1):116-123.
[17]刘方明,孟维韧.土壤线虫作为生态系统恢复指示生物的研究进展[J].安徽农业科学,2008,36(7):2913-2915.LIU F M,MENG W R.Study advance of soil nematodes as bioindicatorof ecosystem restoration[J].Journal of Anhui Agricultural Sciences,2008,36(7):2913-2915(in Chinese).
[18]卓侃,廖金铃.植物线虫分子鉴定研究进展[J].植物保护,2015,41(6):1-8.ZHUO K,LIAO J L.Advances in molecular identification of plant nematodes[J].Plant Protection,2015,41(6):1-8(in Chinese).
[19]PHUPHISUT O,MAIPANICH W,PUBAMPEN S,et al.Molecular identification of the strongyloid nematode Oesophagostomum aculeatum intheAsianwildelephantElephasmaximus[J].JournalofHelminthology,2016,90:434-440.
猜你喜欢
今日农业(2022年14期)2022-09-15
小哥白尼(趣味科学)(2022年5期)2022-08-15
农业科技与信息(2020年14期)2020-12-18
绿色中国(2019年14期)2019-11-26
绿色科技(2019年14期)2019-11-19
中国森林病虫(2019年5期)2019-10-11
江苏农业科学(2019年5期)2019-09-02
陕西农业科学(2019年6期)2019-07-19
中成药(2018年2期)2018-05-09
浙江大学学报(农业与生命科学版)(2017年1期)2017-04-17
