
热空气处理对靖安椪柑幼果柠檬酸代谢的影响
2018-05-04 06:11:12杨滢滢阚超楠陈楚英刘善军陈金印
江西农业大学学报 2018年2期
高 阳,陈 明,杨滢滢,阚超楠,陈楚英,刘善军,陈金印,2*
(1.江西农业大学 农学院/江西省果蔬采后处理关键技术与质量安全协同创新中心/江西省果蔬保鲜与无损检测重点实验室,江西 南昌 330045;2.萍乡学院,江西 萍乡 337055)
柑橘是世界上最重要的商品水果,是我国南方栽培面积最广的果树,它包括的种类很多,主要栽培的有甜橙、宽皮柑橘、柚、金橘和柠檬等,而每一种类又有许多优良品种。靖安椪柑(CitrusreticulataBlanco cv. Ponkan)是颇具特色的江西地方品种,有“远东橘王”之称,具有果形美、色泽艳、肉脆和耐贮藏等优点,但存在果实偏酸等问题[1-2]。
柑橘果实有机酸以柠檬酸为主,柠檬酸代谢是生物体内重要的代谢途径,它是果实品质研究的一个热点[3]。近年来,大多数研究表明,柑橘果实柠檬酸积累水平与合成无直接相关,而受降解速率的调控,肖玉明等[4]研究表明干旱胁迫下温州蜜柑果实柠檬酸大量积累,造成其主要原因是柠檬酸降解酶基因CitIDH与CitNADPIDH表达量下调。GABA途径是果实柠檬酸降解重要环节[5],Cercós等[6]利用柑橘EST 数据,通过基因芯片对柑橘果实发育和成熟阶段的7 000个基因的表达变化进行了研究,提出了柑橘果实成熟期间柠檬酸代谢和利用的一条新途径(GABA途径):柠檬酸先降解成异柠檬酸,再生成2-酮戊二酸和谷氨酸;谷氨酸经谷氨酸脱羧酶(GAD)的催化生成γ-氨基丁酸(GABA),GABA在相关酶的作用下生后生成半醛琥珀酸和琥珀酸[7],该条途径在柠檬酸的降解过程中起着重要作用。
热处理可以有效地促进果实有机酸含量的下降,Wang[8]研究表明‘Northeaster’草莓45 ℃热处理40 min或 42 ℃热处理60~100 min后可滴定酸含量明显降低,且柠檬酸和苹果酸含量均减少。红富士苹果[9]和新余蜜橘[10]经热空气处理后在可滴定酸含量显著降低。同时也有研究发现,热处理能显著降低椪柑贮藏期间有机酸的含量,增加可溶性糖的积累[11]。
为更好地探索柑橘果实柠檬酸的降解机制,本文以含酸量较高的靖安椪柑幼果(盛花后120 d)[12]为试验材料,研究了热空气处理对靖安椪柑果实柠檬酸代谢相关基因表达的影响,以期为提高椪柑果实品质方法的制定提供理论依据。
1 材料与方法
1.1 试验材料
靖安椪柑果实于2014年9月10日(盛花后120 d)采自江西省靖安县果业局果园。果实成熟度和大小一致,采摘后立即运回江西省果蔬保鲜与无损检测重点实验室。
将采摘的果实分为2组:对照组果实不作处理,处理组果实在培养箱[(40±0.5)℃、90%~95% RH]中热空气处理48 h,处理果和对照果采用单果塑料薄膜包装,于温度保鲜柜[(10±1)℃、85%~90% RH]中进行低温贮藏。每10 d取一次样测定相关指标,每个处理每次取样10个果,3次重复。
1.2 测定指标及方法
1.2.1 可滴定酸(TA)含量测定 采用酸碱中和滴定法测定,结果以柠檬酸的量换算。
1.2.2 有机酸含量的提取与测定 果实有机酸含量的提取参照Chen等[13]的方法,采用高效液相色谱法(HPLC)测定其含量。有机酸测定的色谱条件为:C18柱(4.6 mm×250 mm),柱温为25 ℃;流动相为50 mmol/L磷酸氢二氨(pH=2.7),流速为0.5 mL/min;二极管阵列检测器检测。
1.2.3 RNA的提取与逆转录 RNA的提取采用CTAB法进行。提取的RNA用琼脂糖凝胶电泳法进行检测。RNA逆转录采用TaKaRa公司的反转录试剂盒,制备cDNA用于荧光定量分析。
1.2.4 荧光定量PCR 使用Bio-RAD荧光定量PCR仪(CFX96TMReal-Time System),美国TaKaRa公司的SYBR Preminx Ex TaqTM进行Q-PCR基因定量表达分析。内参基因与柠檬酸相关基因引物设计参照陈明[13]的设计,引物序列由华大科技公司合成。
PCR反应体系为25 μL:ddH2O 9.5 μL,上、下游引物各0.5 μL,SYBR(TaKaRa,日本)12.5 μL,cDNA 2 μL。反应程序为:95.0 ℃的预变性30 s,40个循环的95 ℃ 5 s,60 ℃ 30 s;溶解曲线:60 ℃保温30 s,95 ℃保温30 s。
1.3 数据统计与分析
试验采用完全随机设计,使用Excel2003进行数据统计和GraphPad Prism 5 Demo作图,用DPS7.0软件中Duncan’s新复极差法进行显著性差异分析(P<0.05)。
2 结果与分析
2.1 热空气处理对靖安椪柑果实有机酸和可滴定酸含量变化的影响
靖安椪柑果实有机酸以柠檬酸为主,随贮藏时间延长,柠檬酸含量呈不断下降趋势(图1A),但热空气处理显著促进了柠檬酸的降解,热处理48 h后果实柠檬酸含量为33.44 mg/g,显著低于对照果实的35.14 mg/g,热空气处理和对照果实柠檬酸含量在贮藏第60天时分别为28.98 mg/g和32.38 mg/g。奎宁酸和酒石酸含量在贮藏期间均呈先升后降的趋势(图1B、C),热空气处理果实的酒石酸含量在贮藏第2天时为0.90 mg/g,而对照为0.64 mg/g,两者差异显著;热空气处理果实的奎宁酸含量在贮藏第2天时达到最大值7.98 mg/g,而对照在贮藏第10天时才达到最大值8.22 mg/g。靖安椪柑果实苹果酸含量很低(图1D),在整个贮藏期间均低于0.053 mg/g,且两个处理间差异不显著。靖安椪柑果实总有机酸和可滴定酸含量变化整体和柠檬酸类似(图1E、F),均呈整体下降的趋势,从贮藏第2天到第60天热空气处理果实的总有机酸和可滴定酸含量均低于对照。综合分析可知,热空气处理能显著促进靖安椪柑果实有机酸含量的下降,且以降低柠檬酸为主。
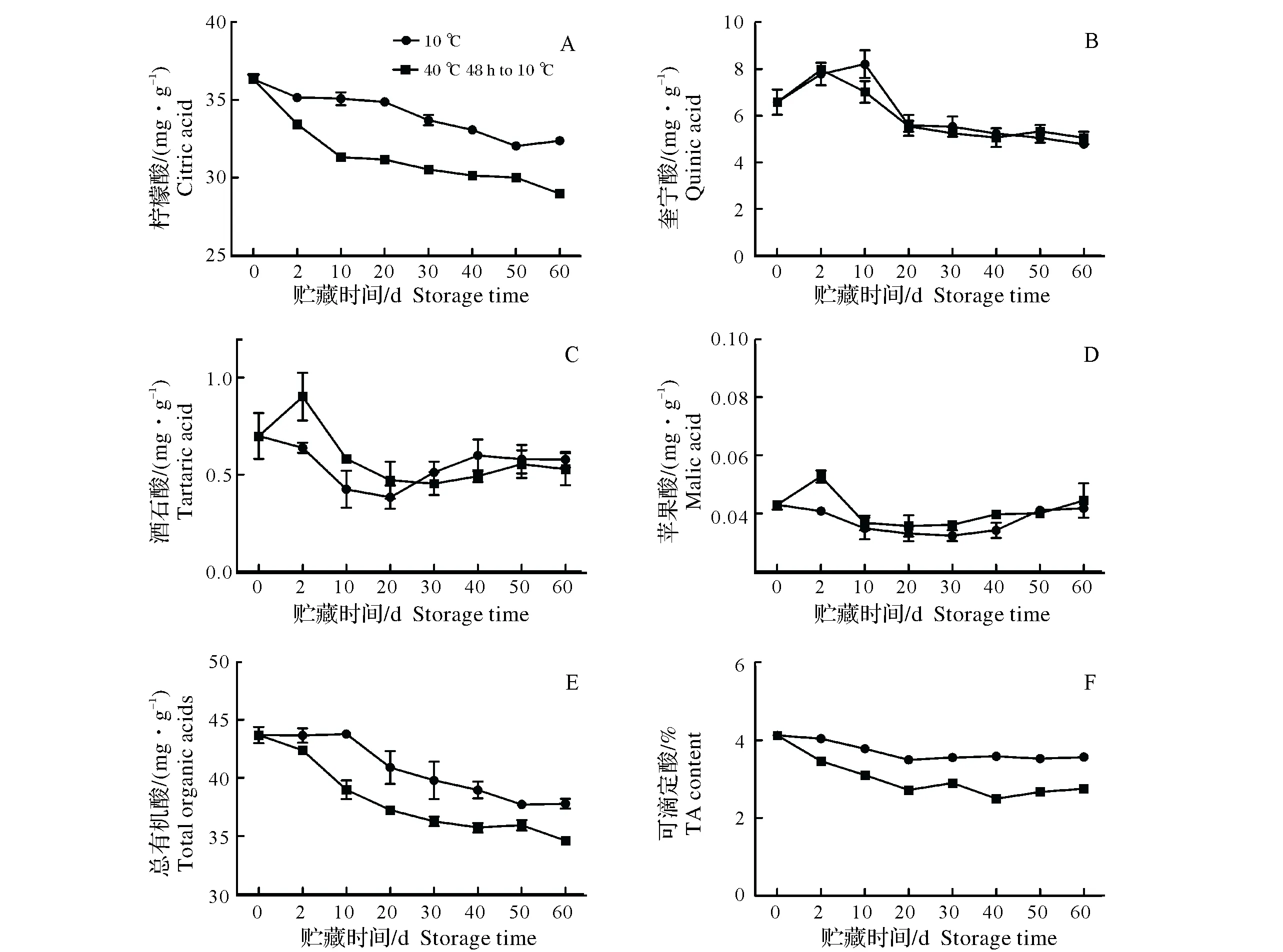
图1 热空气处理对靖安椪柑果实有机酸和可滴定酸含量的影响Fig.1 Effects of hot air treatment on the content of organic acids and TA in Jingan ponkan fruits
2.2 热空气处理对靖安椪柑果实柠檬酸代谢相关基因表达的影响
2.2.1 热空气处理对果实CitCSs表达量的影响 柑橘果实柠檬酸的合成与柠檬酸合成酶(CS)密切相关,在柑橘基因组中有2个CS基因家族成员,分别为CitCS1和CitCS2。从图2可以看出,靖安椪柑果实的CitCS1表达经过热空气处理48 h后显著高于对照,且在整个贮藏期间保持较高水平。CitCS2表达在贮藏期间呈先升后降的趋势,热空气处理诱导CitCS2表达在贮藏前期略高于对照,但从贮藏第40天开始热处理果实一直低于对照。
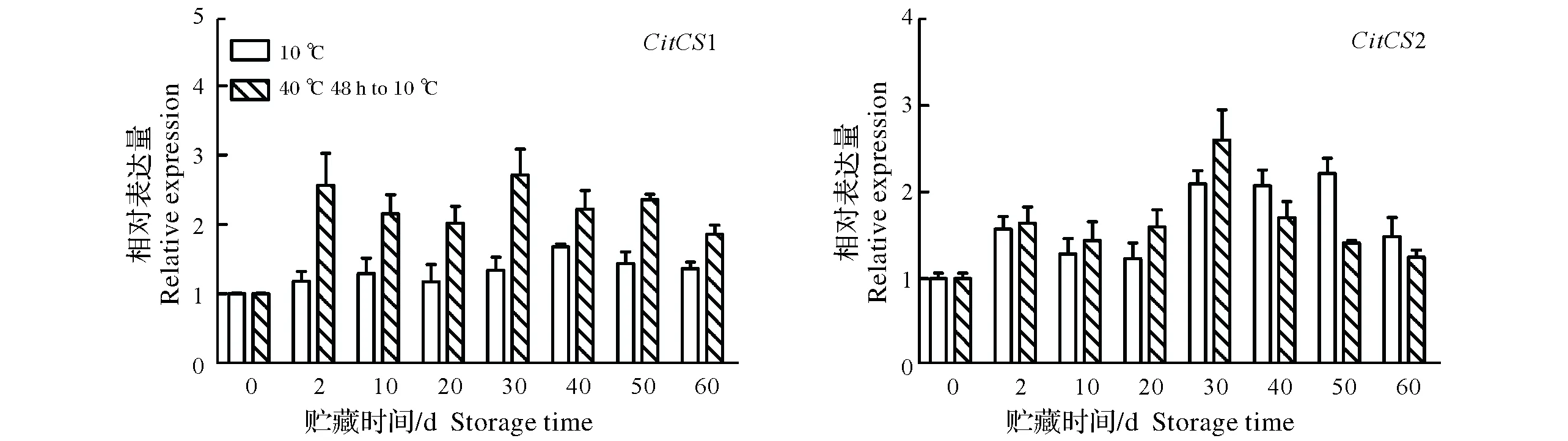
图2 热处理对靖安椪柑果实CitCSs表达量的影响Fig.2 Effects of hot air treatment on expressions of CitCSs in Jingan ponkan fruits
2.2.2 热空气处理对果实CitAcos表达量的影响 顺乌头酸酶(Aco)将柠檬酸催化分解为异柠檬酸。柑橘Aco基因包括CitAco1、CitAco2和CitAco3 3个家族成员。由图3可知,靖安椪柑果实CitAco1表达量随柠檬酸含量的下降呈上升的趋势,热空气处理后从第10天开始果实CitAco1表达量显著高于对照,在贮藏第60天时,热空气处理果实CitAco1表达量是对照的2.07倍。CitAco2表达是呈先上升后下降的趋势,热空气处理果实在贮藏第20天时到达最大,对照在第10天表达量达到最大,但热空气处理后的整个贮藏期间CitAco2表达量均高于对照。靖安椪柑果实CitAco3表达并不稳定,热空气处理仅诱导贮藏前期CitAco3表达,尤其是处理2 d后急剧增加,为对照的3.18倍。

图3 热空气处理对靖安椪柑果实CitAcos表达量的影响Fig.3 Effects of hot air treatment on expressions of CitAcos in Jingan ponkan fruits
2.2.3 热空气处理对果实CitIDHs表达量的影响 异柠檬酸脱氢酶(IDH)参与柠檬酸的降解,它将异柠檬酸降解为2-酮戊二酸。由图4可知,靖安椪柑果实CitIDH1表达量随贮藏时间的增加总体呈递增的趋势,热空气处理果实CitIDH1表达量在贮藏第10天显著高于对照。热空气处理48 h后果实CitIDH2表达量急剧上升,达到了对照的2.98倍,之后其表达量迅速回落,这说明热空气处理能短暂诱导CitIDH2的表达。靖安椪柑果实CitIDH3表达在贮藏期间整体呈上升的趋势,在贮藏第10天时,热空气处理果实CitIDH3表达量显著高于对照,且之后一直高于对照。
2.2.4 热空气处理对果实CitGADs和CitGSs表达量的影响 谷氨酸一方面可以在谷氨酸脱氢酶(GAD)的作用下进入GABA循环,另一方面它还可以在谷氨酰胺合成酶(GS)作用下催化生成谷氨酰胺进入氨基酸代谢。从图5可以看出,CitGAD4表达量在整个贮藏期间呈先升后降的趋势,对照果实在贮藏第30天达到最大值,热空气处理果实CitGAD4表达量在贮藏第2天急剧上升,达到了对照的1.98倍,且一直保持较高水平。而靖安椪柑果实CitGAD5表达量在整个贮藏期间都较低水平,在贮藏第2、10和30天时热空气处理果实CitGAD5表达量显著高于对照。靖安椪柑果实CitGS2表达量在各贮藏期间比较稳定,热空气处理和对照之间无显著性差异。

图4 热空气处理对靖安椪柑果实CitIDHs基因表达量的影响Fig.4 Effects of hot air treatment on expressions of CitIDHs in Jingan ponkan fruits
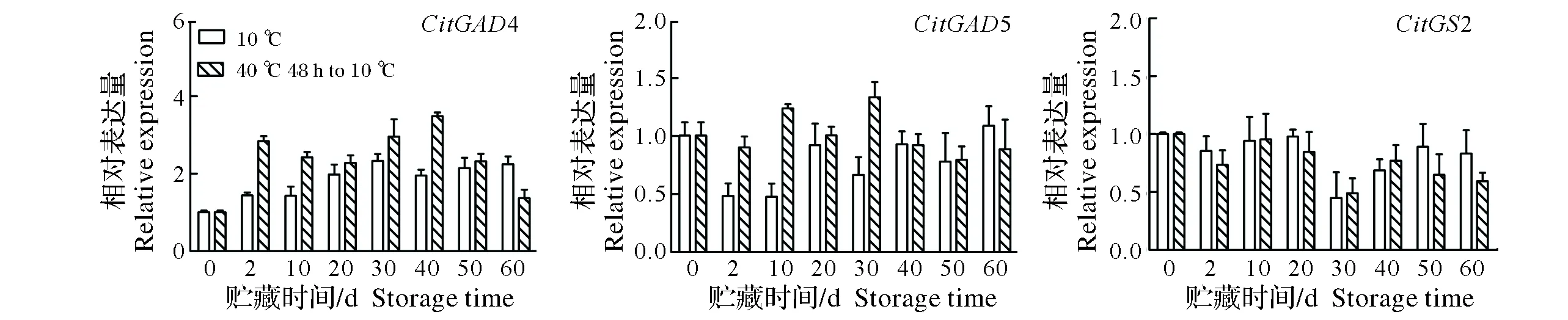
图5 热空气处理对靖安椪柑果实CitGADs和CitGS表达量的影响Fig.5 Effects of hot air treatment on expressions of CitGADs and CitGS in Jingan ponkan fruits
3 讨论与结论
采后热空气处理对果实品质有显著影响,其结果因品种不同而存在差异。杜正顺等[14]用40 ℃热空气处理“丰香”草莓30 min可以明显抑制贮藏期间果实花青素的积累,延缓果实可溶性固形物、可滴定酸含量和硬度下降,保持果实的贮藏品质。Lara等[15]研究表明,热处理能让Dixiland桃柠檬酸和苹果酸含量分别降低50%和20%。关于柑橘果实热处理的研究已有报道,李绍佳[16]和陈明[17]研究表明40 ℃热空气处理48 h能有效降低椪柑果实有机酸含量,提高果实品质。本研究结果表明,热空气处理能显著加速靖安椪柑幼果柠檬酸含量的下降,从而降低有机酸含量,在贮藏后60 d热处理果实柠檬酸含量比对照低1.70 mg/g,这与课题组之前的研究结果[11]一致。
CS广泛存在于果实细胞中,是柠檬酸合成的关键酶[18]。张规富[19]发现不同时期的水分胁迫均能促进发育阶段柑橘果实CitCS1和CitCS2的表达上调,这可能是导致果实有机酸积累原因之一。本试验结果显示,热空气处理诱导靖安椪柑果实CitCS1和CitCS2表达,但同时促进果实柠檬酸含量降低,说明热空气处理可能通过加速柠檬酸的降解来促进靖安椪柑果实有机酸含量的下降,这与前人[11]的研究结果一致。
大量研究表明,柠檬酸的降解与Aco密切相关,Jiang 等[20]发现地表覆膜后椪柑果实Aco活性显著降低,柠檬酸含量显著增加。在番茄中当SlAco3a和SlAco3b受抑制时,Aco活性及其转录水平均下降,成熟果实柠檬酸含量增加。本试验结果表明(40±0.5)℃热空气处理后,靖安椪柑果实基因CitAco1、CitAco2表达量在整个贮藏期均上调,CitAco3表达量在热空气处理2 d后急剧增加,推测热空气处理诱导CitAco1、CitAco2表达可能是幼果期靖安椪柑果实有机酸含量下降的原因之一。IDH在果实有NADP-IDH和NAD-IDH两种存在形式,催化异柠檬酸降解为2-酮戊二酸[21]。热空气处理诱导靖安椪柑果实CitIDH1和CitIDH3表达,短暂诱导CitIDH2的表达,导致柠檬酸进一步降解。
GABA途径是近来年认为是柠檬酸降解的一个重要途径,Aprile等[22]研究发现GABA途径中GAD和氨基酸合成相关酶基因在甜柠檬中呈高表达,认为其参与了柠檬酸降解。LIU 等[23]研究发现注射脱落酸和硫酸钾叶面喷施能显著增加温州蜜柑TA含量,并显著降低GAD活性以及CsGAD1转录,进一步表明CsGAD1在柑橘果实的柠檬酸降解利用中的重要作用。本试验研究发现,热空气处理后,靖安椪柑果实CitGAD4表达量在贮藏第2天急剧上升,且一直保持较高水平,而在整个贮藏期间CitGAD5表达量都较低,且CitGS2基因的表达量在整个贮藏期间比较稳定,热空气处理和对照之间无显著性差异。由此推测热空气处理诱导CitGAD4表达激活GABA途径从而促进柠檬酸降解。
综合以上结果可以得出,热空气处理能促进靖安椪柑幼果柠檬酸的降解,使得果实有机酸含量下降,这种变化和柠檬酸代谢相关基因表达密切相关,其中CitAcos、CitIDH1/3和CitGAD4表达上调可能是导致果实有机酸下降的主要原因。
参考文献:
[1] 罗章.靖安椪柑:远东橘王[J].质量探索,2011,8(6):43.
Luo Z.Jing’an ponkan:far east orange king[J].Quality Exploration,2011,8(6):43.
[2] Lin Q,Wang C,Dong W,et al.Transcriptome and metabolome analyses of sugar and organic acid metabolism in Ponkan(Citrusreticulata) fruit during fruit maturation[J].Gene,2015,554(1):64-74.
[3] Chen L S,Tang N,Jiang H X,et al.Changes in organic acid metabolism differ between roots and leaves of Citrus grandis in response to phosphorus and aluminum interactions[J].Journal of Plant Physiology,2009,166(18):2023-2034.
[4] 肖玉明,卢晓鹏,黄成能,等.水分胁迫对温州蜜柑果实品质及柠檬酸代谢相关基因表达的影响[J].湖南农业大学学报(自然科学版),2014,40(3):281-287.
Xiao Y M,Lu X P,Huang C N,et al.Effects of water stress on the fruit quality of citrate and the expression of genes related to metabolism of citric acid[J].Journal of Hunan Agricultural University(Natural Sciences Edition),2014,40(3):281-287.
[5] Aristoy M C,Orlando L,Navarro J L,et al.Characterization of spanish orange juice for variables used in purity control[J].J Agric Food Chem,1989,37(3):596-600.
[6] Cercós M,Soler G,Iglesias D J,et al.Global analysis of gene expression during development and ripening of citrus fruit flesh.A proposed mechanism for citric acid utilization[J].Plant Molecular Biology,2006,62(4/5):513-527.
[7] 杨滢滢,高阳,郑嘉鹏,等.柑橘果实柠檬酸降解代谢途径与调控研究进展[J].江西农业大学学报,2016,38(3):476-482.
Yang Y Y,Gao Y,Zheng J P,et al.Advances in research on the approach of degradation metabolism of citric acid in citrus fruit and it's regulation[J].Acta Agriculturae Universitatis Jiangxiensis,2016,38(3):476-482.
[8] Wang C Y.Effect of moist hot air treatment on some postharvest quality attributes of strawberries[J].Journal of Food Quality,2000,23(1):51-59.
[9] 张春云,周会玲,张维,等.热处理对红富士苹果虎皮病和贮藏效果的影响[J].西北农林科技大学学报(自然科学版),2013,41(6):117-123.
Zhang C Y,Zhou H L,Zhang W,et al.Effects of hot-air treatment on superficial scald and storage of Red Fujiapples[J].Journal of Northwest A&F University(Natural Science Edition),2013,41(6):117-123.
[10] 陈楚英,彭旋,万春鹏,等.采后热空气处理对新余蜜桔冷藏保鲜效果的影响[J].中国南方果树,2014,43(5):39-41.
Chen C Y,Peng X,Wang C P,et al.Effect of hot air treatment on postharvest preservation of Xinyu tangerine[J].South China Fruits,2014,43(5):39-41,48.
[11] Chen M,Jiang Q,Yin X R,et al.Effect of hot air treatment on organic acid-and sugar-metabolism in Ponkan(Citrusreticulata) fruit[J].Scientia Horticulturae,2012,147(4):118-125.
[12] 高阳,杨滢滢,郑嘉鹏,等.靖安椪柑果实发育阶段糖、酸组分含量变化[J].江西农业大学学报,2016,38(4):631-636.
Gao Y,Yang Y Y,Zheng J P,et al.Changes of sugar and acid components in fruit development of Jing’an orange[J] .Jiangxi Agricultural University,2016,38(4):631-636.
[13]Chen M,Xie X L,Lin Q,et al.Differential expression of organic acid degradation-related genes during fruit development of Navel oranges(Citrussinensis) in two habitats[J] .Plant Mol Biol Rep,2013,31(5):1131-1140.
[14]杜正顺,巩惠芳,汪良驹,等.热水预处理延长冷藏草莓果实保鲜效应的研究[J].江苏农业学报,2008,24(6):922-928.
Du Z S,Gong H F,Wang L J,et al.Hot water pretreatment extending preservation of cold-stored strawberry fruit[J].Journal of Jiangsu Agricultural Sciences 2008,24(6):922-928.
[15] Lara M V,Borsani J,Budde C O,et al.Biochemical and proteomic analysis of ‘Dixiland’ peach fruit(Prunuspersica) upon heat treatment[J].Journal of Experimental Botany,2009,60(15):4315-4333.
[16] 李绍佳.CitERF13 与 CitVHA-c4 协同调控柑橘果实柠檬酸代谢研究[D].杭州:浙江大学,2016.
Li S J.Synergistic effect of CitERF13 and CitVHA-c4 on citric acid metabolism in citrus fruits[D] .Hangzhou:Zhejiang University,2016.
[17] 陈明.椪柑和脐橙果实柠檬酸合成与降解相关基因表达及其调控研究[D].杭州:浙江大学,2013.
Chen M.Citric acid and navel orange fruit citrate synthesis and degradation-related gene expression and regulation[D].Hangzhou:Zhejiang University,2013.
[18] Notton B A,Blanke M M.Phosphoenolpyruvate carboxylase in avocado fruit:purification and properties[J].Phytochemistry,1993,33(6):1333-1337.
[19] 张规富,卢晓鹏,谢深喜.不同时期水分胁迫对椪柑果实柠檬酸代谢相关基因表达的影响[J].果树学报,2015,32(4):525-535.
Zhang G F,Lu X P,Xie S X.Influence of water stress in different development stage on the citric acid metabolism-related genes expression in the Ponkan fruits[J].Journal of Fruit Science,2015,32(4):525-535.
[20] Jiang N,Jin L F,da Silva J A T,et al.Activities of enzymes directly related with sucrose and citric acid metabolism in citrus fruit in response to soil plastic film mulch[J].Scientia Horticulturae,2014,168(3):73-80.
[21] 郝兆丰,袁进成,刘颖慧.异柠檬酸脱氢酶在植物抗氧化胁迫中的作用[J].生物技术通报,2012(6):32-35.
Hao Z F,Yuan J C,Liu Y H.Role of isocitrate dehydrogenase on oxidative stress in plants[J].Biotechnology Bulletin,2012(6):32-35.
[22] Aprile A,Federici C,Close T J,et al.Expression of the H+-ATPase AHA10 proton pump is associated with citric acid accumulation in lemon juice sac cells[J].Functional & Integrative Genomics,2011,11(4):551-563.
[23] Liu X,Hu X M,Jin L F,et al.Identification and transcript analysis of two glutamate decarboxylase genes,CsGAD1 and CsGAD2,reveal the strong relationship between CsGAD1 and citrate utilization in citrus fruit[J].Molecular Biology Reports,2014,41(9):6253-6262.
猜你喜欢
小学科学(2023年9期)2023-04-13 12:53:14
中国水土保持(2022年3期)2022-03-24 12:14:50
家庭医学(下半月)(2019年9期)2019-10-12 08:03:56
消费导刊(2018年10期)2018-08-20 02:56:14
中成药(2018年5期)2018-06-06 03:12:15
食品工业科技(2017年21期)2017-11-21 10:49:59
大众考古(2015年1期)2015-06-26 07:20:36
中国茶叶加工(2015年3期)2015-02-27 07:55:30
石油化工应用(2014年10期)2014-03-11 17:40:26
石油化工应用(2014年9期)2014-03-11 17:40:14
