
mTORC1信号通路及其与糖尿病的相关研究进展
2018-05-02 03:46刘江华
中西医结合心血管病杂志(电子版) 2018年7期
吴 迪,刘江华
(南华大学附属第一医院内分泌科,湖南 衡阳 421001)
1 mTORC1的简介
1.1 mTORC1的结构与功能
mTORC1(mechanistic target of rapamycin complex 1)是由哺乳动物雷帕霉素靶蛋白mTOR、mTOR调节相关蛋白Raptor、哺乳动物LST8同源蛋白mLST8、富脯氨酸AKT底物1-PRAS40和含DEP结构域的mTOR交互蛋白DEPTOR这5种物质构成的多蛋白复合物[1]。其主要作用包括促进细胞的生物合成、细胞生长增殖、营养物质的吸收、抑制细胞自噬等,同时其活性受到来自雷帕霉素、胰岛素、生长因子、某些特定氨基酸及其衍生物、理化因素等一系列因素的严格控制[2]。
1.2 mTORC1信号通路
1.2.1 mTORC1上游信号转导与调节
PI3K-Akt通路:胰岛素样生长因子可通过络氨酸激酶受体RTK激活PI3K-Akt信号通路,使Akt分别于丝氨酸残基939、丝氨酸残基981、苏氨酸残基1462磷酸化结节硬化复合物2(Tuberous Sclerosis Complex 2,TSC2)[3]。这些TSC2的磷酸化的位点可以破坏TSC1/TSC2二聚体,使TSC2与TSC1不再相关联,TSC2失去其GTP酶活化蛋白(GTPase activating protein,GAP)的活性,解除其对Rheb-GTP的水解作用。mTORC1必须通过活性小G蛋白Ras同源蛋白(RHeb)激活,从而经胰岛素信号通路合成蛋白[4]。Akt也可以磷酸化PRAS40,使其从mTORC1中的Raptor上脱落,消除其阻碍Raptor与mTORC1的2个底物4EBP1和S6K结合的作用,从而达到激活mTORC1的目的。
MAPK/ERK通路:有丝分裂原,如胰岛素样生长因子IGF-1可以激活MAPK/ERK通路,抑制TSC1/TSC2复合体,活化mTORC1[5]。当生长因子与临近的络氨酸激酶受体RTK结合时,接头蛋白GRB2结合其SH2结构域,从而募集了能够激活G蛋白RAS的鸟苷酸交换因子Sos。RAS可以激活RAS-RAF-MEK-ERK级联信号转导[6],最终使ERK磷酸化TSC2丝氨酸残基644,激活核糖体S6激酶RSK使其磷酸化TSC2丝氨酸残基1798。这些磷酸化反应可以导致TSC1/TSC2二聚体的分裂,阻止其去活化Rheb,保持mTORC1活性。有相关实验表明,RSK可以磷酸化Raptor,帮助其抵抗PRAS40的抑制作用[7]。
Wnt通路:Wnt通路参与了生物体细胞生长和增殖的调控,有研究表明该通路的激活和mTORC1的激活密切相关,Wnt通路的激活可以抑制糖原激酶GSK3B[8]。当Wnt通路失活时,GSK3B可以使TSC2在丝氨酸残基1341/1337位点磷酸化,同时磷酸化AMPK丝氨酸残基1345位点,而研究表明AMPK的磷酸化是GSK3B磷酸化TSC2的必要前提条件,因此Wnt通路也参与了mTORC1信号通路的调控,也正因为如此mTORC1可以促进生物体内的蛋白质合成[8]。
细胞因子:如TNF-α可以通过IKK2诱导mTOR的激活[9]。IKK2可以使TSC1在丝氨酸残基487/511位点的磷酸化,从而使TSC异二聚体复合物分裂,保持Rheb-GTP结合状态。
能量代谢:翻译需要消耗大量的能量,特别是ATP的消耗巨大。如果ATP消耗过多,水解生成大量的AMP,AMP/ATP比例失衡,就会导致AMPK的激活。AMPK,即AMP依赖的蛋白激酶,是生物能量代谢调节的关键分子,是研究糖尿病及其他代谢相关疾病的核心。它表达于各种代谢相关的器官中,能被机体的运动、多种激素、细胞压力以及影响细胞代谢物质等各种刺激激活。相关研究表明,AMPK是机体保持葡萄糖平衡所必需的,其激活能改善由2型糖尿病引起的代谢失衡[10]。AMPK可以磷酸化TSC2丝氨酸残基1387位点,激活其复合物的GAP活性,将Rheb-GTP水解为Rheb-GDP。通过该途径可以使mTORC1失活,阻碍蛋白质的合成。AMPK还可以磷酸化Raptor的2个丝氨酸残基位点。被磷酸化的Raptor可以招募14-3-3蛋白与之结合,使其可以游离于mTORC1复合体之外。缺少了Raptor的无法招募底物,就无法通过mTORC1合成蛋白质。另外,肿瘤抑制因子丝氨酸/苏氨酸蛋白激酶STK11也可以激活AMPK。深入mTORC1在这方面的研究可能有助于阐明其与肿瘤紧密联系的机制[11]。
低氧应激:当细胞处于低氧环境下时,细胞会通过限制蛋白质的合成以减少能量的消耗,低氧诱导因子HIF1α可以稳定并激活REDD1的转录。REDD1蛋白能与TSC2相结合,阻碍14-3-3蛋白对TSC复合体的抑制作用,保持其GAP活性,维持Rheb与GDP的结合形式,从而抑制mTORC1的活性。此外,在低氧环境下,线粒体ATP合成减少,AMPK活性增加,进而抑制mTORC1活性[12]。
1.2.2 mTORC1下游信号转导与调节
mTORC1主要通过磷酸化下游的真核转录起始因子4E结合蛋白1(4E-BP1)和p70 S6 kinase 1(S6K1)[13],激活mRNA5'端的翻译起始复合物,进而激活翻译,调控细胞的增殖分化、自噬与调亡等。
4E-BP1:活性mTORC1可以磷酸化翻译抑制因子4EBP1,将其从真核转译起始因子eIF4E中释放出来,然后eIF4E便与eIF4A和eIF4G相结合[14]。解旋酶会移除mRNA的5'-UTR区域的5个茎环结构以防止蛋白质过早的翻译,随后翻译起始复合物在5'端帽状结构形成,开始招募小核糖体亚基40S并扫描AUG起始密码子起始位点[15]。当核糖体接触AUG密码子翻译开始。
S6K:去磷酸化的S6K位于eIF3支架复合体上,活性mTORC1被征募至该支架复合体上然后磷酸化S6K使其激活[1]。mTORC1至少磷酸化S6K1的2个氨基酸残基,其中最关键的一个修饰位于苏氨酸残基(T389)上[16]。mTORC1磷酸化S6K1又可以刺激PDK1磷酸化S6K1。活性S6K1可以通过激活S6核糖体蛋白(核糖体的一个组成部分)和eIF4B并使它们被招募形成转录前起始复合物PIC[17]。S6K1也可以通过磷酸化mTORC1负调控区域的2个位点参与mTORC1的正反馈调节刺激mTORC1的活性。S6K1还可以磷酸化泛素酶β-TrCP(BTRC)降解作用的标记物程序性细胞死亡蛋白4(PDCD4),PDCD4是一个肿瘤抑制因子,它可以通过与eIF4A相结合避免参与形成转录起始复合物[18]。
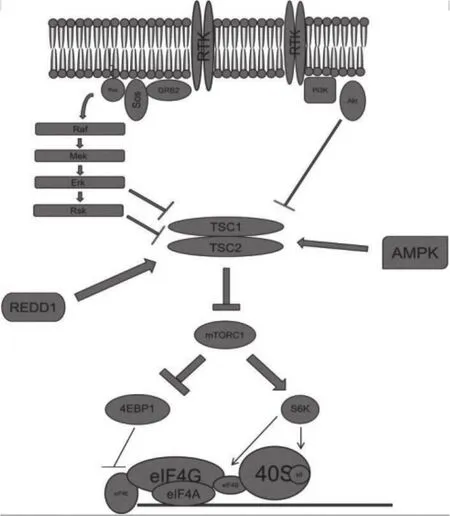
图1 mTORC1信号通路概述
2 mTORC1与糖尿病
2.1 mTORC1与胰岛β细胞
在胰岛β细胞中,mTORC1和mTORC2扮演着各自独特的角色,活性mTORC1通过调节Cyclin D2的合成与稳定性来调控β细胞的生长、增殖[19]。有研究报道对雄性肥沙鼠使用雷帕霉素rapamycin的长期治疗可以导致增加胰岛β细胞凋亡和渐进性高血糖[20]。mTORC1的激活可以促进胰岛β细胞分泌胰岛素降低血糖。近年来,不断有研究表明mTORC1在调控胰岛β细胞功能上存在着两面性,比如在对TSC2敲除小鼠的研究中发现,幼年小鼠表现为β细胞数量增加,出现低血糖和高胰岛素血症,但在老龄小鼠则表现为β细胞数量减少,出现渐进性高血糖和低胰岛素血症[21]。S6K可以在不增加胰岛β细胞数量的前提下,通过增加胰岛素分泌,提高糖耐量,这表明mTORC1主要通过S6K通路调控胰岛素分泌[22]。此外,有相关研究表明β细胞在2型糖尿病中因氧化应激与内质网应激所导致的胰岛素合成、分泌异常增多和β细胞凋亡也与mTORC1相关[23]。
2.2 mTORC1与糖代谢
mTORC1除了能直接通过影响胰岛β细胞的生长、增殖参与到血糖的调控中来,还能通过S6K1途径调控HIF1α和胆固醇调节元件结合蛋白SREBP1和SREBP2参与调控糖代谢[24]。HIF1α可以促进己糖激酶、葡糖糖转运蛋白1、乳酸脱氢酶等基因的表达,促进葡萄糖的吸收与利用。mTORC1对α-酮戊二酸脱氢酶也有抑制其活性的作用[25]。
2.3 mTORC1与自噬
自噬是真核细胞的主要降解途径,细胞必须通过自噬完成从细胞质中移除受损细胞器、蛋白质和小细胞碎片,回收老化、损伤材料,将它们分解然后合成新的细胞结构[26]。自噬也可以清除蛋白质聚集体和破坏细胞器,导致细胞功能障碍。mTORC1在激活后可以磷酸化自噬相关蛋白Atg13,阻止其与Atg1、Atg17、Atg101形成ULK1复合物[27],进而阻止细胞质膜上ULK1复合物参与自噬体前体的形成,抑制自噬[28]。mTORC1抑制自噬的同时刺激蛋白质合成及细胞生长可能导致受损蛋白质和细胞器的积累,进而造成细胞水平的损害[29]。正常自噬过程中出现的问题与糖尿病、心血管疾病、神经退行性疾病和癌症有关。
有不少研究表明,胰岛β细胞自噬基因Atg7的缺失,能导致自噬作用减弱,血清胰岛素水平下降,糖耐量减低[30]。糖尿病模型诱导药物链脲佐菌素STZ能在作用早期增加自噬相关蛋白LC3-VMP1和Beclin-1-VMP1的表达,说明自噬对药物实验早期胰岛β细胞具有保护作用[31]。但是,不同程度的自噬,对胰岛β细胞产生的影响也不尽相同,如2型糖尿病患者中自噬体和自噬泡的累积能引起胰岛β细胞的死亡增加[32]。尽管自噬对胰岛β细胞作用并不十分明确,但可以肯定的是mTORC1—自噬—胰岛β细胞3者之间联系密切。
2.4 mTORC1与细胞免疫及T1DM
雷帕霉素在临床上常作为免疫抑制剂用于抑制T细胞和B细胞的增殖,但是其对mTORC1的抑制作用常常导致生成更多功能更强的记忆T细胞。雷帕霉素对mTORC1的抑制作用不仅能提高T细胞增殖期初始T细胞转变为记忆T细胞前体的能力[33],还可以促进记忆T细胞在收缩期分化为成熟的T细胞[34]。有趣的是,雷帕霉素对mTORC1的抑制可以增加老龄小鼠B细胞数量,增强免疫力[35]。雷帕霉素抑制免疫系统的矛盾现象与多个因素有关,其中包括了与调节性T细胞Treg的相互作用[34]。在过去,mTOR一直被认为是免疫细胞的一个抑制因子,但随后不断有研究表明mTORC1可以促进Treg的生成,增强T细胞抑制免疫的作用,并且mTORC1参与调控的胆固醇和脂质代谢和Treg产生的免疫抑制分子CTLA4和ICOS也存在着一定联系[36]。
目前认为,1型糖尿病是一种以抗原呈递细胞APC和T细胞为介导的自身免疫性疾病,其发病有赖于T细胞(CD4+和CD8+)所表达的抗β细胞抗原反应,CD8+T细胞是启动自身免疫反应的必备条件,而活化的CD4+T细胞则是引起T1DM的必备条件,促炎细胞因子是β细胞的中介因子。已有研究表明,敲除Rheb基因或以雷帕霉素抑制CD4+T细胞的mTORC1活性,可以抑制Th1、Th17细胞分化,促进Th2细胞分化[37]。因此可以推断mTORC1与1型糖尿病之间存在着某种联系。
3 结 语
mTORC1活性受细胞内多种信号调控,其中细胞因子、能量代谢、细胞环境等信号主要经由TSC1/TSC2-Rheb途径传递。氨基酸等营养分子类信号激活mTORC1的机制尚不十分清楚,但普遍被认为是通过一条不同的独立途径,有众多蛋白质复合物参与该通路的调控。本文未就营养分子如氨基酸介导的mTORC1信号通路的在溶酶体上激活机制作阐述,但该通路已有众多突破性研究进展,如该信号通路的核心骨架Ragulator五元复合物调控RHeb,mTORC1在溶酶体上的定位、组装并招募下游蛋白的作用机制[38];KICSTOR复合体中四个蛋白分子在GATOR1定位于溶酶体和mTORC1信号激活调控中的作用等[39]。
尽管在过去的十余年中,mTORC1信号通路在调控细胞生长、增殖、凋亡、介导免疫等各方面机制有了较为深入的了解,与糖代谢脂质代谢等代谢性疾病的联系也十分明确,进一步研究明确其调控机制可以为糖尿病及其他相关疾病的治疗提供潜在的新思路和方法。
[1]Wullschleger S,Loewith R, Hall MN. TOR signaling in growth and metabolism[J].Cell. 2006 Feb 10;124(3):471-84.
[2]Bond P.Regulation of mTORC1 by growth factors, energy status,amino acids and mechanical stimuli at a glance[J].J Int Soc Sports Nutr.,2016,13:8.
[3]Ma XM,Blenis J. Molecular mechanisms of mTOR-mediated translational control[J].Nat Rev Mol Cell Biol.2009,10(5):307-18.
[4]Mendoza MC1,Er EE,BlenisJ."The Ras-ERK and PI3K-mTOR pathways: cross-talk and compensation[J].Trends Biochem Sci.2011,36(6):320-8.
[5]Mendoza MC, Er EE,Blenis J.The Ras-ERK and PI3K-mTOR pathways: cross-talk and compensation[J].Trends Biochem Sci,2011,36(6):320-8.
[6]McCubrey JA,Steelman LS,Chappell WH,et al.Ras/Raf/MEK/ERK and PI3K/PTEN/Akt/mTOR cascade inhibitors: how mutations can result in therapy resistance and how to overcome resistance[J].Oncotarget,2012,3(10):1068-111.
[7]Carrière A,Cargnello M,Julien LA,et al.MAPK signaling stimulates mTORC1 activity by promoting RSK-mediated raptor phosphorylation[J].Curr Biol,2008,18(17):1269-77.
[8]Majid S,Saini S,Dahiya R.Wnt signaling pathways in urological cancers: past decades and still growing[J].Mol Cancer,2012,10;11:7.
[9]Salminen A,Hyttinen JM,Kauppinen A, el at.Context-Dependent Regulation of Autophagy by IKK-NF-κB Signaling: Impact on the Aging Process[J].Int J Cell Biol,2012.
[10]Zhang BB,Zhou G,Li C.AMPK:an emerging drug target for diabetes and the metabolic syndrome[J].Cell Metab,2009,9(5):407-16.
[11]Nagalingam A,Arbiser JL,Bonner MY, et al. Honokiol activates AMP-activated protein kinase in breast cancer cells via an LKB1-dependent pathway and inhibits breast carcinogenesis[J].Breast Cancer Res,2012,21;14(1):R35.
[12]Wang S,Song P,Zou MH.AMP-activated protein kinase,stress responses and cardiovascular diseases[J].Clin Sci(Lond),2012,122(12):555-73.
[13]Hay N,Sonenberg N.Upstream and downstream of mTOR[J].Genes Dev,2004,18(16):1926-45.
[14]Wang H,Zhang Q, Wen Q,et al.Proline-rich Akt substrate of 40kDa(PRAS40):a novel downstream target of PI3k/Akt signaling pathway[J].Cell Signal,201224(1):17-24.
[15]Lee T,Pelletier J. Eukaryotic initiation factor 4F: a vulnerability of tumor cells[J].Future Med Chem,2012,4(1):19-31.
[16]Saitoh M, Pullen N, Brennan P,et al.Regulation of an activated S6 kinase 1 variant reveals a novel mammalian target of rapamycin phosphorylation site[J].J Biol Chem.2002,277(22):20104-12.
[17]Peterson RT, Schreiber SL,Translation control: connecting mitogens and the ribosome[J].Curr Biol,1998,8(7):R248-50.
[18]Schmid T, Jansen AP,Baker AR, et al.Translation inhibitor Pdcd4 is targeted for degradation during tumor promotion[J].Cancer Res.2008,68(5):1254-60.
[19]Balcazar N,Sathyamurthy A,Elghazi L, et al. mTORC1 activation regulates beta-cell mass and proliferation by modulation of cyclin D2 synthesis and stability[J].J Biol Chem,2005,280(27):25485-90.
[20]M Fraenkel, M Ketzinel-Gilad,Y Ariav,et al.mTOR inhibition by rapamycin prevents beta-cell adaptation to hyperglycemia and exacerbates the metabolic state in type 2 diabetes[J].Diabetes,2008,57(4):945-957.
[21]Ardestani A,Lupse B, Kido Y,et al.mTORC1Signaling:A Double-Edged Sword in Diabetic β Cells[J].Cell Metab,2018,27(2):314-331.
[22]Rachdi L, Balcazar N,Osorio-Duque F,et al.Disruption of Tsc2 in pancreatic beta cells induces beta cell mass expansion and improved glucose tolerance in a TORC1-dependent manner[J].Proc Natl Acad Sci U S A. 2008,105(27):9250-5.
[23]Wang J1, Yang X1,Zhang J.Bridges between mitochondrial oxidative stress,ER stress and mTOR signaling in pancreatic β cells[J].Cell Signal,2016,28(8):1099-104.
[24]Düvel K,Yecies JL,Menon S,et al. Activation of a metabolic gene regulatory network downstream of mTOR complex 1[J].Mol Cell,2010,30;39(2):171-83.
[25]Shimodahira M1,Fujimoto S,Mukai E,et al.Rapamycin impairs metabolism-secretion coupling in rat pancreatic islets by suppressing carbohydrate metabolism[J].J Endocrinol.2010,204(1):37-46.
[26]Choi AM,Ryter SW,Levine B. Autophagy in human health and disease[J].N Engl J Med. 2013,368(19):1845.
[27]Alers S,Löffler AS,Wesselborg S,et al. Role of AMPK-mTORUlk1/2 in the regulation of autophagy:cross talk,shortcuts, and feedbacks[J].Mol Cell Biol,2012,32(1):2-11.
[28]Pyo JO, Nah J,Jung YK. Molecules and their functions in autophagy[J].Exp Mol Med,2012,44(2):73-80.
[29]Proud CG.Amino acids and mTOR signalling in anabolic function[J].Biochem Soc Trans,2007,35(Pt 5):1187-90.
[30]Fujitani Y,Kawamori R, Watada H.The role of autophagy in pancreatic beta-cell and diabetes[J].Autophagy,2009,5(2):280-2.
[31]Grasso D.Sacchetti ML,Bruno L,et al.Autophagy and VMP1 expression are early cellular ev ents in experimental diabetes[J].Pancreatology,2009,9(1):81-88.
[32]Masini M,Bugliani M,Lupi R,et al.Autophagy in human type 2 diabetes pancreatic beta cells[J].Diabetolog ia,2009,52(6):1083-1086.
[33]Araki K, Turner AP, Shaffer VO,et al.mTOR regulates memory CD8 T-cell diあerentiation[J].Nature,2009,460(7251):108-12.
[34]Araki K,Youngblood B,Ahmed R.The role of mTOR in memory CD8 T-cell diあerentiation[J].Immunol Rev.2010,235(1):234-43.
[35]Chen C,Liu Y, Liu Y, Zheng P . mTOR regulation and therapeutic rejuvenation of aging hematopoietic stem cells[J].Sci Signal,2009,2(98):ra75.
[36]Zeng H1,Yang K, Cloer C, et al.mTORC1 couples immune signals and metabolic programming to establish T(reg)-cell function[J].Nature.2013,499(7459):485-90.
[37]Delgoffe GM1, Pollizzi KN,Waickman AT, et al. The kinase mTOR regulates the diあerentiation of helper T cells through the selective activation of signaling by mTORC1 and mTORC2[J].Nat Immunol,2011,12(4):295-303.
[38]Zhang T, Wang R,Wang Z,et al.Structural basis for Ragulator functioning as a scaあold in membrane-anchoring of Rag GTPases and mTORC1[J].Nat Commun. 2017,8(1):1394.
[39]Wolfson RL, Chantranupong L,Wyant GA, et al. KICSTOR recruits GATOR1 to the lysosome and is necessary for nutrients to regulate mTORC1[J].Nature.2017,543(7645):438-442.
猜你喜欢
生物化学与生物物理进展(2022年7期)2022-07-25
生物化学与生物物理进展(2022年6期)2022-07-21
食品安全导刊(2022年17期)2022-07-04
九江学院学报(自然科学版)(2022年2期)2022-07-02
中国农业科学(2022年12期)2022-06-28
中国畜牧杂志(2022年1期)2022-01-20
西南农业学报(2021年10期)2021-12-14
南昌大学学报(医学版)(2021年1期)2021-11-29
中学生数理化(高中版.高考理化)(2021年2期)2021-03-19
中成药(2019年12期)2020-01-04
