
不同氮营养盐浓度对斜生栅藻生长及化学组成的影响
2018-04-28 06:20罗晓霞刘锦上李长玲
广东农业科学 2018年1期
罗晓霞,刘锦上,李长玲
(1.广东海洋大学水产学院/广东省藻类养殖及应用工程技术中心,广东 湛江 524088;2.茂名市金阳热带海珍养殖有限公司,广东 茂名 525000)
在“藻类→浮游动物→鱼类”的水生食物网环节中,初级生产者(藻类)营养的好坏是草食性浮游动物产量的限制因子之一[1-2],而浮游动物的产量则影响了最上层营养级鱼类的产量。初级生产者(藻类)的氮、磷含量决定了其作为浮游动物的食物质量[3-6]。环境中氮缺乏会导致藻体的多不饱和脂肪酸(PUFAs)降低[7],从而限制浮游动物的产量[8]。有研究发现藻体N含量越低,浮游动物的稳定同位素δ15N值越高,浮游动物摄食不同氮含量的同一种藻类时,其稳定同位素δ15N值变化范围为0~6‰[9]。因此,初级生产者(藻类)的氮含量变化会影响整个水生食物网的生产力及营养级判断,尤其在营养盐浓度变化较大的水体。水体中营养盐的含量不仅会影响藻类的生长率,同时还会影响藻类细胞的生化组成[5-6,10]。不同氮磷营养盐浓度对米氏凯伦藻(Karenia mikimotoi)[11]、铜绿微囊藻(Microcystis aeruginosa)[12]、刚毛藻(Cladophora glomerata)[13]、小球藻(Chlorella sp.)[14]等藻类生长的影响已有相关报道。藻类的生长速率也会影响藻体的 C、N、P 比值[10]。Hessen等[5]发现月牙藻(Selenastrum capricornutum)在不同磷营养盐浓度下藻体C、N、P含量显著不同。然而,目前对于不同氮营养盐对藻类化学组成(C、N、P)含量的研究较少。因此,我们研究了不同浓度氮营养盐对斜生栅藻生长和化学成分的影响,以期为水生食物网能量传递影响机制提供基础参考资料。
1 材料与方法
1.1 试验材料
供试藻种为斜生栅藻(Scenedesmus obliqnus),由暨南大学水生所藻种室提供,采用BG11培养液培养。
1.2 试验方法
以BG11培养基配方为基础,用超纯水配置成无氮培养基,再以NaNO3为氮源,通过添加不同体积的NaNO3得到不同氮浓度的培养液。试验设7个初始氮质量浓度处理:0.247、2.47、24.7、123.5 、247、370.5、740 mg/L,每个处理 3次重复。
接种培养:斜生栅藻扩大培养10 d后,选处于指数生长期的藻进行实验。在无菌操作台(VS-840K,苏泰)把藻液加入配制好的不同氮质量浓度的培养液,起始藻密度控制在20×104个/mL。培养容器为1 000 mL 锥形瓶,每瓶藻液为400 mL,用光照恒温培养振荡器(TS-111GZ型)进行培养。培养温度为25(±1)℃,光照强度为2 800 lx,光暗比14 h∶12 h。培养6 d后,对藻类进行计数,测藻体N、P含量。
栅藻密度的计数:实验结束时,取20 mL藻液,用鲁哥氏液固定,并用血球计数板进行计数。
1.3 藻体N、P含量测定
藻体N、P含量的测定方法分别按国标GB11894-89、GB111893-89的方法进行,用紫外分光光度计(UV-5500)进行测定。每个重复取10 mL藻液离心,然后把上层清液倒掉,再补充相同体积的超纯水,如此反复离心2次后,补充超纯水至10 mL。最后,用吸管把藻液打均匀,取1 mL测藻体N含量,取2 mL测藻体P含量。
1.4 藻体C含量的测量
实验结束时,用预燃烧(45℃,4 h)的GF/F 膜(Whatman)抽滤50 mL藻液,置于60℃烘至恒重,经CHNS/O 元素分析仪测其碳重。计算藻类平均生长率μ:

式中,t为藻类培养时间,N0为起始细胞数,Nt为培养时间t后的细胞数。
试验数据用SPSS 17.0 进行单因素方差分析,原始数据符合方差齐性,运用Tukey’s HSD法进行差异显著性测验。
2 结果与分析
2.1 不同氮浓度对栅藻生长的影响
从图1可以看出,不同氮浓度对斜生栅藻的生长有显著影响,在初始氮浓度为0.247~247 mg/L时,藻体生长率随氮浓度的升高而升高,氮浓度为247 mg/L时,其生长率是最低氮浓度(0.247 mg/L)的2.6倍。然而,当氮浓度超过247 mg/L时,细胞生长率显著下降,高氮浓度处理(370 mg/L及740 mg/L)的生长率显著低于247 mg/L氮浓度处理。
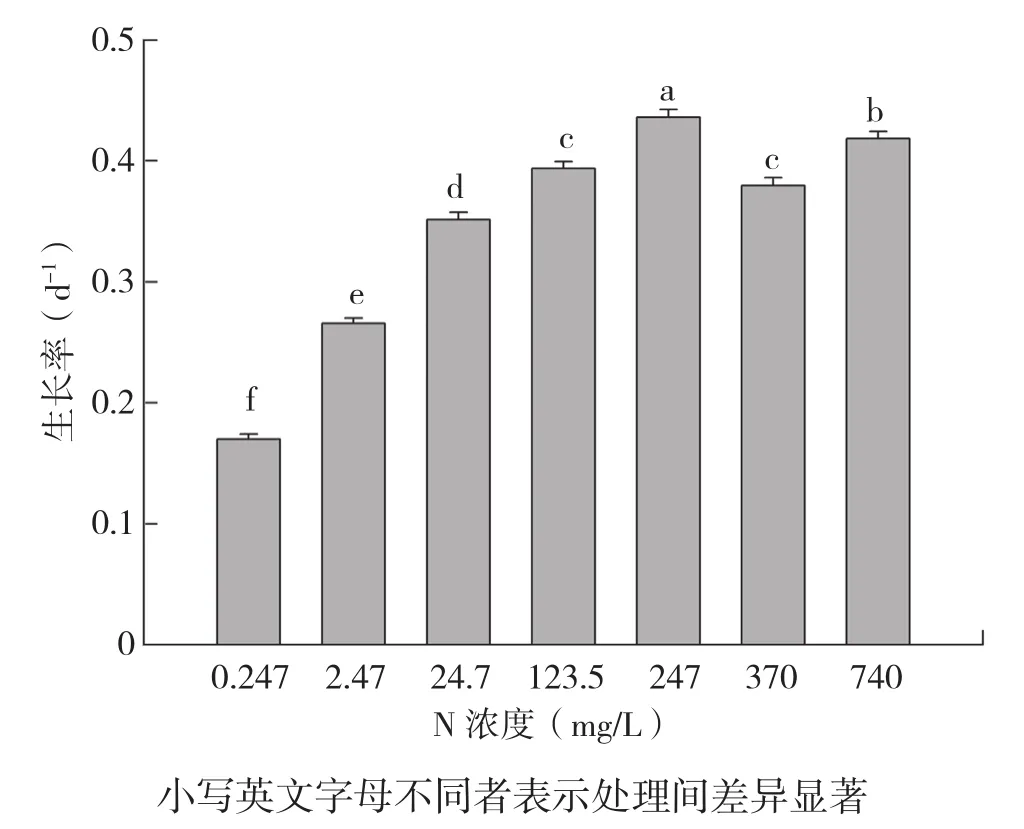
图1 不同氮浓度处理斜生栅藻的生长率
2.2 不同氮浓度对栅藻化学组成的影响
从表1可以看出,不同氮浓度对栅藻藻体N含量有显著影响,在初始氮浓度为0.247~24.7 mg/L时,藻体单位体积N含量随着初始氮浓度的增加而增加,但是随着氮浓度继续增加(≥24.7 mg/L),各处理栅藻N含量差异并不明显。当氮浓度为0.247 mg/L时,其单位细胞N含量显著低于其他氮浓度处理。

表1 不同氮浓度处理栅藻藻体的N含量
不同初始氮浓度对栅藻藻体P含量影响显著(表2)。随着氮浓度的逐渐增加,栅藻单位体积P含量呈先增加后降低的趋势。氮浓度为2.47~247 mg/L时单位体积P含量无显著差异,均显著高于最低氮浓度(0.247 mg/L)。但是当氮浓度增大至370~740 mg/L时,栅藻单位体积P含量则显著下降,其值显著低于其他氮浓度处理。
栅藻单位细胞P含量则随着初始氮浓度的增加而急剧下降。最低氮浓度(0.247 mg/L)处理单位细胞P含量是最高氮浓度(740 mg/L)处理的8倍。
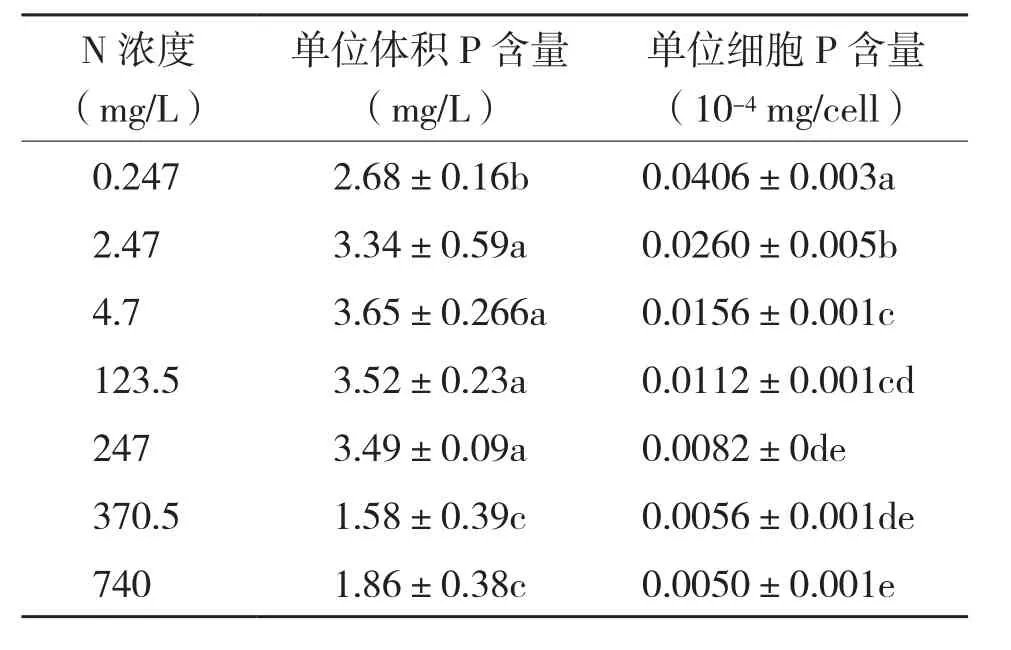
表2 不同氮浓度处理栅藻藻体的P含量
栅藻单位体积C含量随着初始氮浓度的增加而增加,在氮浓度为247 mg/L时达到最高值,之后随着氮浓度的继续增加而下降(表3)。单位细胞C含量在低氮浓度下最高,随后逐渐下降,在氮浓度为24.7~370 mg/L时达到平稳状态。

表3 不同氮浓度处理栅藻藻体的C含量
不同氮浓度对栅藻C、N、P比率有明显的影响,栅藻的C∶N∶P原子比随着初始氮浓度的增加而增加(表4)。

表4 不同N浓度对栅藻藻体C∶N∶P比率的影响
3 结论与讨论
3.1 不同氮质量浓度对斜生栅藻生长的影响
有研究表明,营养盐浓度是影响藻类生长的重要影响因素[10-12,14-16]。在氮浓度为 0~1 g/L时,杜氏藻的细胞密度、生长率随着氮浓度的增加而增加[17]。米氏凯伦藻的最高细胞密度和生长率在30~750,mg/L,NaNO3浓度间随氮浓度的升高而升高[11]。本研究也发现类似的结果,在初始氮浓度为0.247~247 mg/L时,藻体细胞密度随氮浓度的升高而升高。但是氮浓度过高也会抑制藻类的生长。杜氏藻在氮浓度超过1.0 g/L时,其生长下降[17]。米氏凯伦藻的细胞密度在氮浓度高达750~3 000 mg/L时随氮浓度的升高而降低[11]。本试验中,斜生栅藻的细胞密度在氮浓度为247 mg/L时达到最高峰,之后随着氮浓度的增加而显著下降。曹春晖等[11]认为高氮磷条件会导致植物细胞单盐毒害,从而抑制藻细胞的分裂,使藻细胞进入对数期时间推迟,从而使高氮条件下稳定期细胞密度及生长率低于中氮磷条件。过高或过低的氮浓度条件下,培养液氮磷比的失调也是抑制藻类生长的原因之一。
不同氮磷比的营养盐对藻细胞的生长有明显影响,而不同的藻类,其生长的最适氮磷比不同[15]。新月柱鞘藻在营养盐N∶P质量比为160∶1 时,生长速率最快;而青岛大扁藻和米氏凯伦藻分别在4∶1 和80∶1 的条件下生长速率最快[15]。丰茂武等[12-13]研究发现,当氮磷质量比为40∶1时,铜绿微囊藻、刚毛藻的生长最佳。本试验中培养液的N∶P质量比随着氮浓度的增高而增加,栅藻的生长趋势则随着培养液N∶P比的升高呈现先增加后下降的趋势,生长最佳的氮浓度(247 mg/L)所对应的培养液N∶P质量比为46∶1,与丰茂武等[12]的研究结果相似。本试验中,初始氮浓度0.247~2.47 mg/L为斜生栅藻生长的限制因子,斜生栅藻在氮浓度为24.7~740 mg/L(培养液N∶P比为46∶1)时能正常生长,最佳生长氮浓度为247 mg/L。
本试验发现栅藻在氮限制(氮浓度为0.247~2.47 mg/L)的情况下,藻体细胞比正常氮浓度下培养的藻体细胞个体更大,而且细胞蛋白核消失或变小。Hessen等[5]发现月牙藻(Selenastrum capricornutum)在磷限制的情况下,藻体细胞增大,出现细胞壁增厚现象。Van Donk等[18]认为绿藻在磷缺乏的情况下,藻体自身会存储大量的碳作淀粉颗粒,因此导致细胞个体增大。Harrison等[19]发现藻类在氮限制的情况下,自身的碳水化合物增高,蛋白质含量下降。我们在前期实验发现栅藻在磷限制的情况下,藻体个体细胞增大,细胞壁增厚(待发表)。因此,在营养盐限制条件下,某些微藻的细胞形态会发生不同程度的变异,从而更加适应复杂的环境。
3.2 不同氮质量浓度对栅藻化学组成的影响
水体中营养盐的含量不仅会影响藻类的生长率,同时还会影响藻类细胞的生化组成[5-6,10,20]。Hessen 等[5]发现月牙藻藻体的C、N、P含量与培养液中的磷浓度呈正相关。本试验中随着初始氮浓度的增加,单位体积栅藻的C、N、P含量显著不同。在氮浓度为0.247~24.7 mg/L时,藻体单位体积N含量(mg/L)随着初始氮浓度的增加而增加,但是之后随着氮浓度继续增加,各浓度处理间栅藻的N含量差异不明显,因此,初始氮浓度24.7 mg/L为栅藻N含量吸收的饱合浓度。栅藻单位体积P含量(mg/L)随着氮浓度的逐渐增加,呈先增加后下降的趋势,因此,氮浓度太高抑制藻体P含量的增长。在氮缺乏时,栅藻个体细胞P含量(mg/cell)显著增加,其细胞P含量是高氮浓度处理的8倍。表明在氮缺乏的情况下,细胞个体大量吸收周围含量充足的磷元素,以达到体内的生化平衡,这也是藻类自我调整的策略。
Redfield定律认为,藻类细胞组成的原子比率为 C∶N∶P=106∶16∶1[10,21-22]。本试验发现,在不同氮浓度下,藻体C∶N∶P原子比率不同,为10∶0.14∶1~62∶4.6∶1,其值与Redfield比值相距较大。有研究发现当外部营养源、光照强度超过藻类生长所需的条件时,Redfield 定律就失去其应用范围[5,12,23]。Goldman等[10]发现在没有营养盐限制、浮游植物生长率较高的环境,浮游植物的化学原子比通常趋向于Redfield比率C∶N∶P=106∶16∶1。海链藻(Thalassiosira pseudonana)在生长率低于10%时,其N∶P原子比为5∶1;当生长率高于90%时,N∶P比为15∶1,逐渐接近Redfield比值的N∶P=16∶1。本试验中,栅藻的生长周期为6 d,还未达到对数期,即生长率未达到最高,因此藻体的N∶P比值较低,未能体现Redfield值的定律。而在氮限制的情况下,藻体的C∶N∶P比值更低至10∶0.14∶1,表明藻体自身的N含量极低。有研究发现氮营养盐不足,藻体的PUFAs含量急剧下降[7],而藻类PUFAs含量决定了更高营养级浮游动物的产量[1,7,19],因此环境中的氮浓度不仅影响藻类的生长、生化成分,而且进一步影响更高营养级浮游动物。不同氮含量的斜生栅藻对于浮游动物生长的影响还有待进一步研究证实,从而为水生食物网能量传递影响机制提供基础参考资料。
参考文献:
[1]Saikia S S. Nandi C and P in aquatic food chain:a review on C:P stoichiometry and PUFA regulation[J]. Knowledge and Management of Aquatic Ecosystems,2010(3):1-14.
[2]Elser J J,Fagan W F,Denno R F,et al.Nutritional constraints in terrestrial and freshwater food webs[J]. Nature,2000,408(6812):578-580.
[3]Plath K M. Boersma Mineral limitation of zooplankton:stoichiometric constraints and optimal foraging[J]. Ecology,2001,82(5):1260-1269.
[4]Sterner R W. Daphnia growth on varying quality of Scenedesmus:mineral limitation of zooplankton[J]. Ecology,1993:2351-2360.
[5]Hessen D O,Færøvig P J T. Andersen Light,nutrients,and P:C ratios in algae:grazer performance related to food quality and quantity[J]. Ecology,2002,83(7):1886-1898.
[6]DeMott W R,Gulati R D K. Siewertsen Effects of phosphorus‐deficient diets on the carbon and phosphorus balance of Daphnia magna[J].Limnology and oceanography,1998,43(6):1147-1161.
[7]廖启斌,李文权. 营养盐对三角褐指藻脂肪酸含量与百分组成的影响[J]. 海洋环境科学,2000,19(2):6-9.
[8]徐怡,胡忠军,刘其根,等. 温度和不同氮磷浓度培养的小球藻对透明溞生长和繁殖的影响[J]. 上海海洋大学学报,2011,20(5):712-719.
[9]Adams T S,Sterner. The effect of dietary nitrogen content on trophic level15N enrichment[J].Limnology and oceanography,2000,45(3):601-607.
[10]Goldman J C,McCarthy J J D G. Peavey Growth rate influence on the chemical composition of phytoplankton in oceanic waters[J]. Nature,1979,279(5710):210-215.
[11]曹春晖,刘文岭,施定基,等. 不同氮磷浓度对米氏凯伦藻生长的影响[J]. 天津科技大学学报,2010,25(2):22-25.
[12]丰茂武,吴云海,冯仕训,等. 不同氮磷比对藻类生长的影响[J]. 生态环境,2008,17(5):1759-1763.
[13]张猛,石瑛,丁义晶. 不同氮磷比和 Fe3+浓度对刚毛藻生长繁殖的影响[J]. 江苏农业科学,2013,41(6):196-198.
[14]马彩虹,麻晓霞,马玉龙,等. 不同氮磷浓度对小球藻生长性能的影响[J]. 安徽农业科学,2012,40(22):11367-11369.
[15]孙军,刘东艳,陈宗涛,等. 不同氮磷比率对青岛大扁藻,新月柱鞘藻和米氏凯伦藻生长影响及其生存策略研究[J]. 应用生态学报,2004(11):2122-2126.
[16]Juneja A,Ceballos R M,Murthy G S. Effects of environmental factors and nutrient availability on the biochemical composition of algae for biofuels production:a review[J]. Energies,2013,6(9):4607-4638.
[17]王俊,马玉心,崔大练,等. 不同浓度氮、磷对杜氏盐藻生长的影响[J]. 浙江海洋学院学报:自然科学版,2012,31(2):142-146.
[18]Van Donk E,Lurling M,Hessen D,et al.Changed cell wall morphology and reduced grazer vulnerability in nutrient deficient phytoplankton.in Phycologia[C]. USA:Int phycological soc new business office,1997.
[19]Harrison P,Thompson P,Calderwood G. Effects of nutrient and light limitation on the biochemical composition of phytoplankton[J]. Journal of Applied Phycology,1990,2(1):45-56.
[20]Feng P,Deng Z,Fan L,et al. Lipid accumulation and growth characteristics of Chlorella zofingiensis under different nitrate and phosphate concentrations[J]. Journal of bioscience and bioengineering,2012,114(4):405-410.
[21]Redfield A C. The influence of organisms on the composition of seawater[J]. The sea,1963,2:26-77.
[22]陈洁,高超. Redfield比值在富营养化研究中的应用及发展[J]. 四川环境,2016,35(6):109-114.
[23]徐燕青,陈建芳,高生泉,等. 太平洋中西部海域浮游植物营养盐的潜在限制[J]. 生态学报,2012,32(2):394-401.
猜你喜欢
林业与生态科学(2022年4期)2022-12-24
今日农业(2022年14期)2022-11-10
上海蔬菜(2022年2期)2022-04-29
国际商业技术(2022年4期)2022-04-21
当代水产(2021年8期)2021-11-04
潍坊学院学报(2020年2期)2021-01-18
军事文摘(2020年20期)2020-11-16
防护林科技(2020年9期)2020-11-09
今日农业(2019年10期)2019-01-04
中南林业调查规划(2017年3期)2017-12-29
