
12种润楠属植物叶柄解剖学特征研究
2018-04-25 09:30:02薛洪富赵振军徐思远费永俊
西南林业大学学报 2018年2期
张 建 薛洪富 赵振军 向 云 徐思远 费永俊
(长江大学园艺园林学院,湖北 荆州 434025)
樟科 (Lauraceae) 润楠属 (Machilus) 植物,全世界总共约100多种,主要分布于东南亚和东亚,我国有68种,主要分布于西南、中南部至台湾,北达山东、湖北及甘肃、陕西南部[1-3]。润楠属植物是我国南方主要的经济林木,在园林应用、木材、香料等方面占有重要地位,同时具有一定的药用价值[4]。目前润楠属植物分类学的研究主要集中在经典形态学方面,如花被裂片外是否被毛,果实的大小及形状等。近年来,也有少数研究人员利用叶表皮形态学[5-7]、孢粉学[8]、化学成分[9-10]、分子生物学[11-12]等方法对部分润楠属植物进行研究,以寻找适用于润楠属属下分类系统的辅助分类特征及属内种间的系统演化关系的证据。目前润楠属植物的分类系统尚不完善,本研究以长江大学楠木种质资源评价与创新中心收集的润楠属植物为研究对象,拟通过石蜡切片法,对12种润楠属植物的叶柄进行形态解剖学的比较研究,从而寻找出新的润楠属属下分类特征,为进一步解决属内种间系统关系问题以及资源利用提供参考依据。
1 材料与方法
1.1 材料采集
实验材料为12种润楠属植物,分别为薄叶润楠 (Machilusleptophylla)、芳槁润楠 (Machilussuaveolens)、粉叶润楠 (Machilusglaucifolia)、广东润楠 (Machiluskwangtungensis)、红楠 (Machilusthunbergii)、基脉润楠 (Machilusdecursinervis)、龙眼润楠 (Machilusoculodracontis)、木姜润楠 (Machiluslitseifolia)、润楠 (Machiluspingii)、塔序润楠 (Machiluspyramidalis)、狭叶润楠 (Machilusrehderi) 和长梗润楠 (Machiluslongipedicellata),均为种植在长江大学西校区盆景园的2年生植株。 2016年4月,采集12种润楠属植物的叶柄,并立即置于70%FAA固定液中,4 ℃冰箱保存备用。
1.2 研究方法
采用石蜡切片法,用刀片将叶柄切成3 mm左右若干段,切片厚度8~10 μm,番红-固绿对染,中性树胶封片,然后在显微镜下进行观察、拍照,叶柄整体观察采用10倍物镜,维管束观察采用40倍物镜。
观测指标包括横切面形状、下表皮到维管束距离比 (下表皮到维管束距离/叶柄宽)、表皮细胞形状、维管束长宽比 (维管束长/维管束宽)、叶柄横切面长宽比 (叶柄横切面长/宽) 及叶柄上表面形状。
2 结果与分析
2.1 叶柄中部横切面的基本结构分析
薄叶润楠 (图1-1) 外观呈马蹄形,表皮细胞排列紧密,染色较深,皮层细胞排列疏松,紧挨韧皮部的薄壁细胞体积明显缩小,韧皮部细胞染色情况较深,可能与含有大量粘液物质有关。木质部细胞排列呈 “C” 型,下表皮与木质部之间的薄壁细胞染色较浅,体积稍大。芳槁润楠 (图1-2) 表皮外面明显观察到蜡质层,皮层薄壁细胞椭圆形,在韧皮部和临近韧皮部的皮层细胞中,部分细胞呈现黑色,这可能是油性细胞或粘液细胞染色造成的。总体来看,木质部和韧皮部所占叶柄的比例较小。粉叶润楠 (图1-3) 也可以观察到明显的蜡质层,表皮细胞由2层构成,韧皮部细胞中观察到较多染成黑色的细胞,木质部细胞中可观察到体积明显增大的木质部细胞。广东润楠 (图1-4) 外观呈Ω形,在表皮和外皮层中可观察到染色较深的细胞,中皮层中染色较深细胞较少,韧皮部细胞中染色较深细胞所占比例较大,木质部细胞的外壁染色也较深,在维管射线中也可观察到染色较深的细胞。总体来看,广东润楠维管束结构不够明显,染色较深细胞较多,可能这与广东润楠含有较多油性细胞或粘液细胞有关。红楠 (图1-5) 外观与广东润楠相似,表皮染色较深,皮层中散生染色较深细胞,韧皮部细胞染色较深,呈黑色,木质部细胞大小不一。基脉润楠 (图1-6) 可观察到明显的蜡质层,维管束较其他润楠属植物有明显伸长,韧皮部中观察到体积较大染色较深的细胞,但数量明显偏少,木质部细胞多为方形。龙眼润楠 (图1-7) 表皮和外皮层染色普遍较深,皮层细胞排列也较为紧密,韧皮部染色情况与广东润楠情况相似,木质部细胞排列整齐。木姜润楠 (图1-8) 染色较深细胞较少,只在皮层有少量分布,韧皮部细胞染色较深,木质部细胞排列整齐。润楠 (图1-9) 整体染色较深,表皮、外皮层染色较深,皮层中也分布较多的染色较深细胞,韧皮部所占比例较小,木质部细胞排列整齐,在下表皮和木质部之间,可观察到2个体积明显偏大的粘液细胞。塔序润楠 (图1-10) 染色较深,在韧皮部上方分布有类似木质部细胞的结构,韧皮部中也分布染色较深的大细胞,木质部细胞排列整齐紧密,有厚壁细胞的存在。狭叶润楠 (图1-11) 呈明显的Ω形,皮层中分布数量较多体积较大染色较深的细胞,维管束呈明显的 “C” 形,木质部细胞排列整齐有规则。长梗润楠 (图1-12) 表皮和外皮层细胞染色较深,在维管束左右两侧的皮层细胞分布较多染色较深细胞,韧皮部染色较深,韧皮部细胞大小不一。
根据上述分析结果可知,12种润楠属叶柄中部的横切面形状基本都呈半圆形态,但大致可分为马蹄形和Ω形2种,马蹄形润楠属植物包括薄叶润楠、芳槁润楠、粉叶润楠、基脉润楠、龙眼润楠、塔序润楠和长梗润楠,Ω形润楠属植物包括广东润楠、红楠、木姜润楠、润楠和狭叶润楠;表皮细胞有2种形状,分别为长方形和近似圆形2种;表皮细胞以内区域大部分为薄壁组织,维管束部分呈 “C” 字形规则的分布于其中;以横切面下表皮一侧为判断位置,表皮细胞到维管束距离相对比 (表皮到维管束距离/叶柄宽) 差异也有较大区别,从狭叶润楠的0.282到木姜润楠的0.504,相对数据之间的差异表明上表皮一侧的薄壁组织的相对厚度有所不同,结合上表皮一侧的薄壁组织的相对厚度和观察结果来看,薄叶润楠,广东润楠、龙眼润楠和狭叶润楠远轴一侧薄壁组织相对最薄;芳槁润楠、粉叶润楠、红楠、基脉润楠、润楠、塔序润楠和长梗润楠厚度居中;木姜润楠最厚。在横切面上表皮一侧观察中发现,该侧形状不一,分内凹和外凸2种。内凹部分又有所区别,如薄叶润楠,中间呈 “v” 型沟;芳槁润楠、木姜润楠、狭叶润楠、长梗润楠内凹程度较大;粉叶润楠和基脉润楠内凹程度较低。外凸部分也有所差别,如龙眼润楠凸出程度较低,而广东润楠、红楠、润楠、塔序润楠凸出程度较大。叶柄中部横切面的长宽比有明显的区别,主要分2类,第1类为叶柄横切面中长大于宽,第2类为叶柄横切面的长小于宽,除了润楠和塔序润楠为第2类外,其余10种均为第1类。
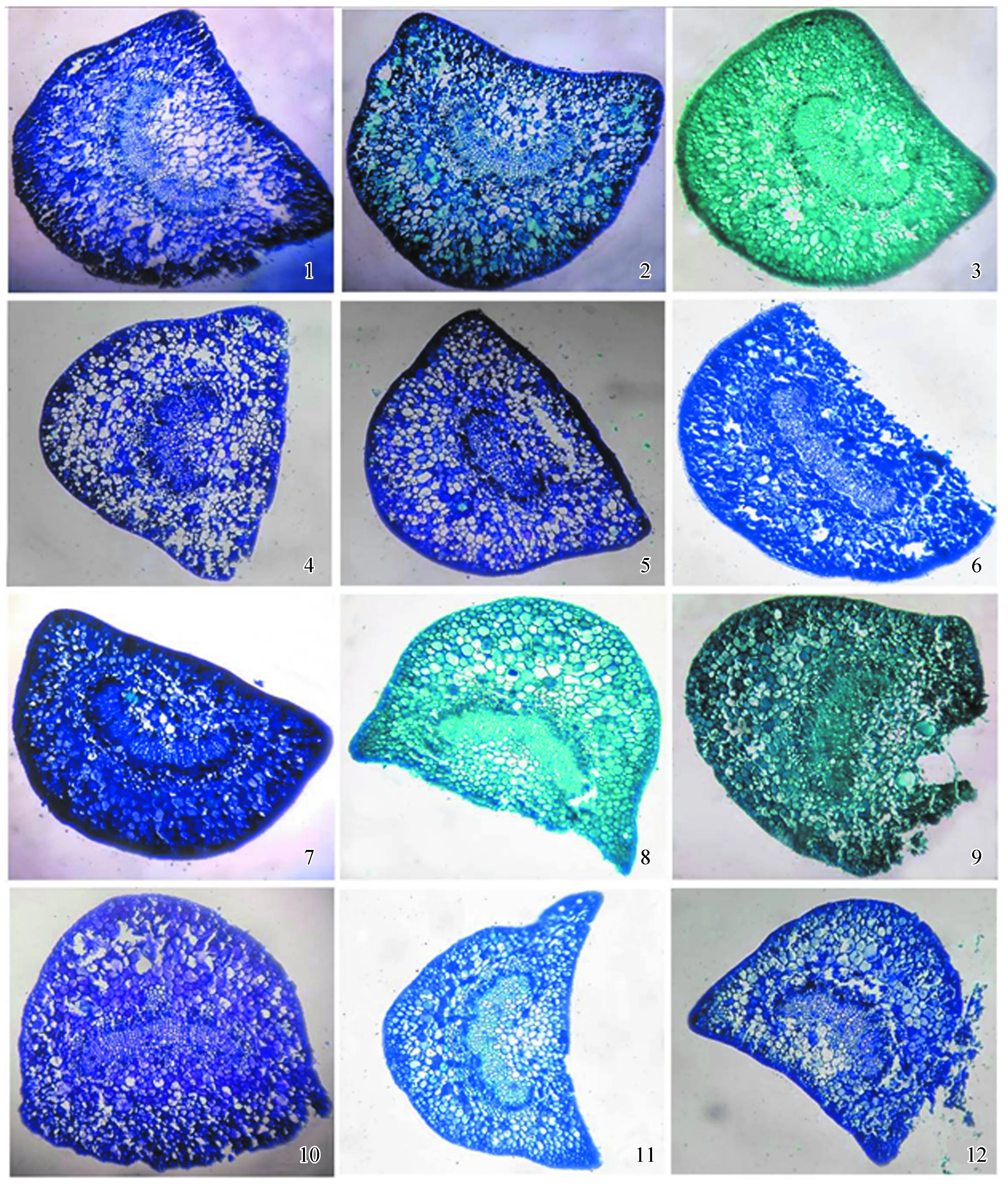
1~12依次是薄叶润楠、芳槁润楠、粉叶润楠、广东润楠、红楠、基脉润楠、龙眼润楠、木姜润楠、润楠、塔序润楠、狭叶润楠、长梗润楠。
图112种润楠属植物叶柄中部横切面观察(×10)
Fig.1 Observation of the middle transverse section of petioles of 12Machilus(×10)
2.2 叶柄中部横切面的维管束结构分析
观察发现,12种润楠属植物叶柄部横切面的维管束部分均呈 “C” 型居于叶柄中部偏近上表皮一侧(图2)。薄叶润楠(图2-1)紧邻韧皮部的皮层细胞有体积较大染色较深的细胞,韧皮部细胞排列不整齐,木质部部分细胞存在细胞壁加厚。芳槁润楠(图2-2)紧邻韧皮部的细胞存在大量厚壁细胞,韧皮部细胞染色较深,木质部细胞存在较多细胞壁加厚细胞。粉叶润楠(图2-3)韧皮部散生染色较深体积较大细胞,可观察到形成层细胞,木质部细胞存在大小2种类型。广东润楠(图2-4)韧皮部细胞染色较深,木质部细胞存在细胞壁加厚情况。红楠(图2-5)韧皮部细胞染色较深,木质部细胞排列不整齐。基脉润楠(图2-6)维管束较其他润楠属植物有明显伸长,韧皮部中观察到体积较大染色较深的细胞,但数量较少,木质部细胞形态不规则。龙眼润楠(图2-7)韧皮部染色较深,木质部细胞排列整齐。木姜润楠(图2-8)韧皮部细胞染色较深,木质部细胞排列整齐。润楠 (图2-9) 韧皮部细胞体积较小,数量偏少,木质部细胞排列整齐。塔序润楠 (图2-10) 韧皮部中有少量染色较深的大细胞,木质部细胞排列整齐紧密,有厚壁细胞的存在。狭叶润楠 (图2-11) 紧邻韧皮部的皮层细胞有染色加深的大细胞,韧皮部排列不规则,木质部细胞排列整齐有规则。长梗润楠 (图2-12) 韧皮部中存在染色较深的大细胞,木质部细胞存在细胞壁加厚现象。
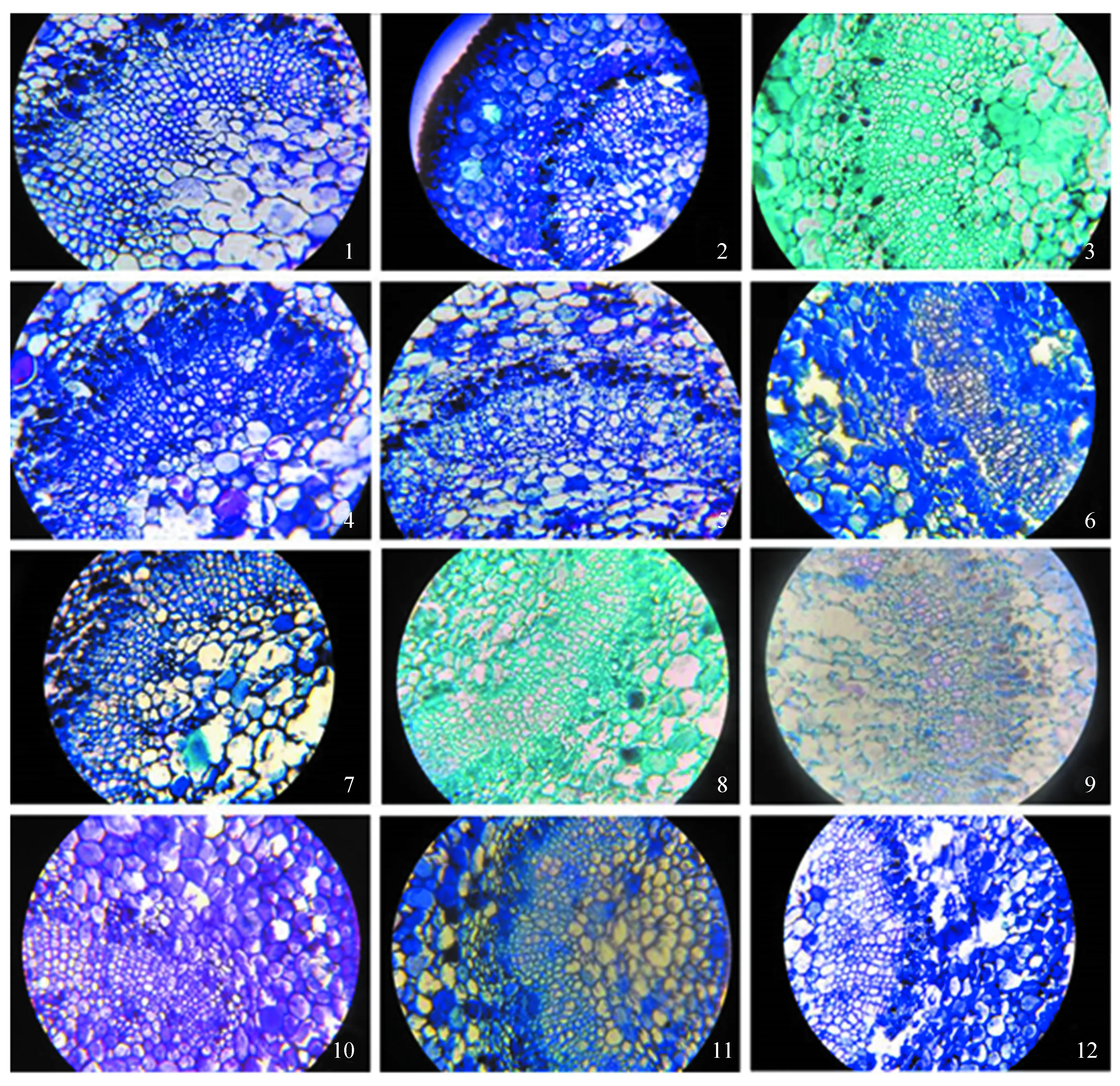
1~12依次是薄叶润楠、芳槁润楠、粉叶润楠、广东润楠、红楠、基脉润楠、龙眼润楠、木姜润楠、润楠、塔序润楠、狭叶润楠、长梗润楠。
图212种润楠属植物叶柄中部维管束观察(×40)
Fig.2 Observation of the middle transverse section of vascular bundle of 12Machilus(×40)
维管束长宽比主要分3种情况 (表1):广东润楠、润楠和长梗润楠维管束长宽比在2~3之间;芳槁润楠、粉叶润楠、红楠、基脉润楠、木姜润楠、塔序润楠和狭叶润楠维管束长宽比在3~4之间;薄叶润楠和龙眼润楠维管束长宽比为4倍以上。
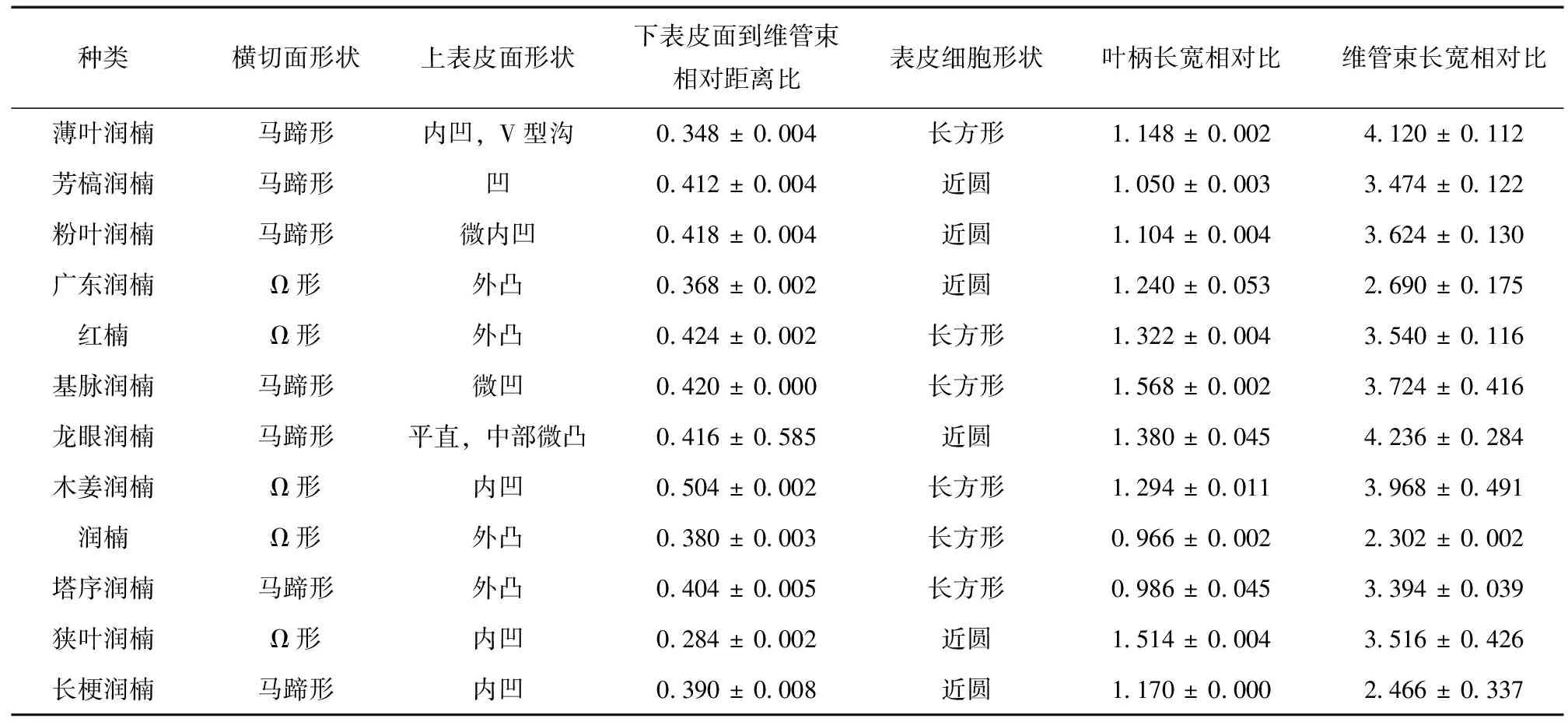
表1 12种润楠属植物参数测定Table 1 Determination of 12 Species of Machilus
2.3 基于12种润楠属植物叶柄解剖特征的聚类分析
根据表1的测定结果,以下表皮到维管束的相对距离比、维管束长宽比、叶柄长宽比、叶柄形状、叶柄表皮细胞形状和上表皮细胞形状6个性状参数对12种润楠属材料进行对聚类分析,定性形状以0 (否) 或1 (是) 表示,用Ntsys软件应用UPGMA分析法进行对比分析,结果见图3。
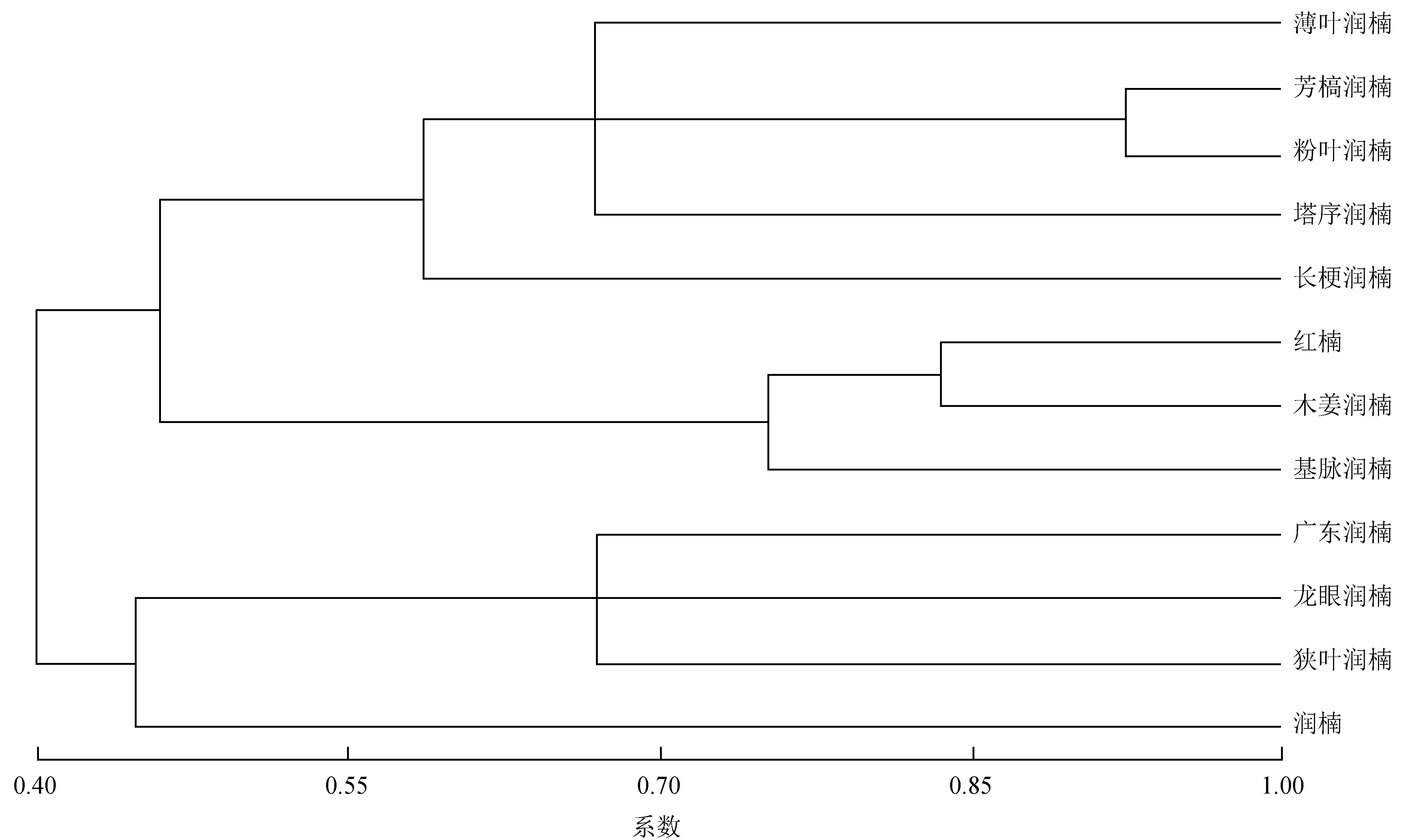
图3基于6个解剖参数的12种润楠属植物聚类分析图
Fig.3 Clustering analysis diagram based on 6 anatomic parameters of 12Machilus
聚类结果表明:12种润楠属植物可分为4类:第1类分别为薄叶润楠、芳稿润楠、粉叶润楠、塔序润楠和长梗润楠;第2类分别为红楠、木姜润楠和基脉润楠;第3类为广东润楠、龙眼润楠和狭叶润楠;第4类为润楠。研究还发现,第1类中的芳稿润楠和粉叶润楠亲缘关系最为紧密,薄叶润楠和塔序润楠次之,长梗润楠与其他4种润楠属植物亲缘关系较远,但均属于毛花组[1],与传统分类学观点相一致;第2类的红楠、木姜润和基脉润楠亲缘关系较近,均属于红楠亚组,这也与传统分类学观点相一致;第3类中的广东润楠为黄心树亚组、龙眼润楠为枇杷叶润楠亚组、狭叶润楠为红楠亚组,这3种不同的亚组归为一类,与传统的分类结果不一致,这可能是由于测定指标较少造成的,关于这3种润楠属植物的系统分类学地位,还需要进一步的实验数据进行确定。润楠作为润楠属植物的模式种,单独作为一类。
根据12种润楠属植物叶柄的解剖特征来看,12种润楠属植物之间确实存在着明显的差异,此可以作为种间鉴别的重要辅助依据,对种间分类学也具有一定的参考意义。
3 结论与讨论
叶片性状受种源分布[13]、光照、营养[14]等外界条件影响较大,而叶柄内部结构比较稳定,可以作为植物分类的主要参考指标。叶柄解剖的比较研究在高等植物分类中具有重要的作用,对于种间亲缘关系的研究也非常有帮助[15]。王涛等[16]对11种鼠尾草属 (Salvia) 的叶柄进行石蜡切片观察,发现叶柄内部结构上鼠尾草种间表现出了较大的差异,其中维管束数量以及木质部、韧皮部厚度可以作为鼠尾草属植物鉴定和分类学研究提供有力依据。宋文婵等[17]对8种蒲公英属 (Taraxacum) 植物的叶柄进行解剖观察,发现叶柄中部横切面形状、维管束类型可以作为种间分类鉴定的重要依据。Salmaki等[18]对山芝麻属 (Helicteres) 叶形态解剖的研究结果表明叶柄的横切面、径轴长、极轴长以及维管束数目可以作为种间分类的主要判定依据。
通过对12种润楠属植物叶柄的石蜡切片观察,表明叶柄解剖特征可作为种间鉴别特征的辅助判断标准。润楠属植物分类研究多以成年株为对象,从茎、叶、花、果等特征进行描述[1-2],并辅以叶表皮特征[5-6]、孢粉学特征[8]以及分子手段[11-12]进行分类验证,但均缺乏组织形态学的证据,对于幼苗的分类鉴定存在很大难度。本研究以2年生实生苗的叶柄为研究对象,采用石蜡切片的方法,从显微结构上进行观测,本实验研究结果表明叶柄横截面形状、上部叶柄形状、叶柄表皮细胞有形状、维管束长宽比、叶柄长宽比以及下表皮到维管束相对距离比等指标较为稳定,可以作为种间分类的依据,这也为其他润楠属植物的种间鉴别提供了参考。叶柄形态解剖的观察呈现出的新特征对推断润楠属下种间分类非常有帮助。此外,通过实验观察,发现部分润楠属植物叶柄中分布有油细胞和粘液细胞,这与楠属 (Phoebe) 植物叶片中存在油细胞和粘液细胞[19]的结果相吻合。实验中发现,油细胞和粘液细胞的数量以及分布在不同润楠属植物中存在一定差异,比如广东润楠和狭叶润楠,存在明显的粘液细胞,这也许可以作为属内分布的一个辅助标准。
石蜡切片过程中,发现部分润楠属植物叶柄切片并不完整,对于这些种的石蜡切片方法也做了部分改进,但效果不理想,这可能与叶柄中所含特定化学成分有关,从而导致图片效果不太理想。目前很多文献对润楠属部分植物进行了化学成分研究的报道,其主要成分包括丁内酷类、木脂素类,黄酮类,生物碱类,倍半萜类,三萜类和其他结构类型[20-21],挥发油及其生物活性也有报道[22],其中丁内脂类和木脂素类化合物是润楠属植物的特征性代表成分。在石蜡切片观察中,发现不同种之间存在染色差异,并且部分种存在特殊类型细胞,比如芳槁润楠在紧邻韧皮层的皮层中,存在数量较多的厚壁细胞,与其他种类有明显不同。因此,在石蜡切片基础上,还应进行组织化学方面的观察,以完善补充种间系统分类学证据。
[参 考 文 献]
[1] 中国科学院中国植物志编辑委员会.中国植物志: 第31卷[M]. 北京: 科学出版社, 1982.
[2] 中国树木志编辑委员会. 中国树木志: 第1卷[M]. 北京: 中国林业出版社, 1983.
[3] 林夏珍. 浙江润楠属植物的数量分类[J]. 林业科学, 2007, 43(11): 151-156.
[4] 卜鹏滨, 里艳茹, 姜明, 等. 粗壮润楠中的木脂素类成分[J]. 中国中药杂志, 2013, 38(11): 1740-1746.
[5] 庄雪影, 张粤, 孙同. 香港润楠属植物叶表皮形态及分类学意义[J]. 华南农业大学学报, 2002, 23(1): 52-54.
[6] 钟义, 夏念和. 国产润楠属植物的叶表皮特征及其系统学意义[J]. 热带亚热带植物学报, 2010, 18(2): 109-121.
[7] 沈雪梅. 17种樟科润楠属植物的数量分类与分子分类研究[D]. 上海: 华东师范大学, 2015.
[8] 王玉国, 韦发. 樟科两个近缘属润楠属和楠木属花粉形态的比较研究[J]. 广西植物, 2003, 23(1): 27-30, 96-99.
[9] 戴磊, 冯志坚, 李文峰. 4种润楠属植物精油成分分析[J]. 福建林业科技, 2013, 40(1): 49-51.
[10] 李文锋. 广东樟属和润楠属植物精油成分分析[D]. 广州: 华南农业大学, 2010.
[11] 陈俊秋, 李朗, 李捷, 等. 樟科润楠属植物ITS序列贝叶斯分析及其系统学意义[J]. 云南植物研究, 2009, 31(2): 117-126.
[12] 刘玉香, 宋晓琛, 江香梅. 润楠ISSR-PCR优化反应体系建立及引物筛选[J]. 林业科技开发, 2013, 27(5): 24-28.
[13] 陈丽君, 刘明骞, 廖柏勇, 等. 苦楝不同种源叶片性状变异研究[J]. 西南林业大学学报, 2016, 36(5): 16-20.
[14] 尚旭岚, 李琼琼, 邓波, 等. 光照和施肥对青钱柳幼苗叶片性状与解剖结构的影响[J]. 西南林业大学学报, 2014, 34(6): 9-15.
[15] Simpson M G. Plant Systematic[M]. Pittsburgh: Academic Press, 2006.
[16] 王涛, 刘世勇, 王龙, 等. 11种鼠尾草属叶片和叶柄解剖特征及其分类学意义[J]. 中国中药杂志, 2014, 39(14): 2629-2634.
[17] 宋文婵, 宁伟, 赵鑫, 等. 8种蒲公英属植物叶柄解剖学特征与分类学意义[J]. 西北植物学报, 2011, 31(3): 468-474.
[18] Salmaki Y, Zarre S, Lindqvist C, et al. Comparative leaf anatomy of Stachys (Lamiaceae: Lamioideae) in Iran with a discussion on its subgeneric classification[J]. Plant Systematics and Evolution, 2011, 294(1): 109-125.
[19] 周存宇, 万小丽, 张建, 等. 五种楠属植物叶片油细胞和黏液细胞的比较[J]. 湖北农业科学, 2015(18): 4506-4508, 4518.
[20] Cheng W, Zhu C, Xu W, et al. Chemical constituents of the bark ofMachiluswangchianaand their biological activities[J].J Nat Prod,2009,72(12):2145-2152.
[21] Li Y, Cheng W, Zhu C, et al. Bioactive neolignans and lignans from the bark ofMachilusrobusta[J]. J Nat Prod, 2011, 74(6): 1444-1452.
[22] Ho C, Liao P, Su Y. Composition and antimicrobial activities of the leaf essential oil of Maehilus zuihoensis from Taiwan[J]. Brazilian Journal of Pharmacognosy, 2012, 22(2): 277-283.
猜你喜欢
河北农业大学学报(2021年5期)2021-11-10 13:11:28
植物研究(2021年2期)2021-02-26 08:40:10
新疆农业科学(2020年9期)2020-10-13 12:37:42
河南农业科学(2019年9期)2019-09-24 09:10:48
天津农业科学(2015年11期)2015-12-03 00:03:49
儿童故事画报·发现号趣味百科(2015年7期)2015-10-23 00:59:48
江苏农业科学(2015年4期)2015-06-15 21:11:35
热带作物学报(2014年3期)2014-04-29 18:17:49
安徽农学通报(2014年7期)2014-04-29 03:00:24
中国果业信息(2014年11期)2014-01-23 13:43:33
