
基于RNA-Seq分析褪黑激素对绒山羊皮肤基因表达的影响
2018-04-24 05:11付绍印王建蒙
畜牧与饲料科学 2018年2期
丽 春 ,付绍印 ,王建蒙 ,王 位
(1.内蒙古民族大学动物科学技术学院,内蒙古 通辽 028000;2.内蒙古自治区农牧业科学院,内蒙古 呼和浩特010031;3.内蒙古ATCG生物信息研究所,内蒙古 呼和浩特 010020;4.内蒙古农业大学动物科学学院,内蒙古 呼和浩特 010018)
褪黑激素(melatonin,MT)是哺乳动物松果体分泌的主要产物,是一种吲哚类激素,具有多种生理功能,如抗衰老、抗氧化、抗肿瘤、调节时差、调节免疫、调节毛皮动物绒毛周期和色素沉积等。近年来,对毛皮动物的研究表明,MT与非生绒季节促绒生长和季节性换毛有关[1-6]。绒山羊的绒毛生长受光照影响,呈现明显的周期性变化,其毛囊在一年内经历兴盛、退行、休止3个时期。研究发现,外源褪黑激素处理可显著影响绒山羊绒毛的生长[2,7-9]。 然而截至目前,人们对褪黑激素调控绒毛生长的机理仍然知之甚少。
RNA-Seq,即转录组测序,是基因结构及功能研究的基础和出发点,通过高通量测序RNASeq,能够获得某一物种特定组织或器官在某一状态下的几乎所有转录本序列及表达信息,已被广泛用于基础生物学研究、临床疾病诊断以及药物开发等领域。通过RNA-Seq试验可准确测定基因的表达水平、等位基因转录本的特异表达及可变剪接,以此来阐明生物机体发生生理或病理学变化的分子机制[10]。
目前,从全局水平了解褪黑激素影响绒山羊皮肤毛囊基因表达的研究较少。该研究以处于皮肤毛囊发育生长起始点(5月)的内蒙古绒山羊罕山型个体为研究对象,基于RNA-Seq技术在基因转录层次上对皮下埋植MT个体和非埋植MT个体的皮肤基因表达情况进行比较研究,旨在筛选与MT影响毛囊发育调控相关的候选基因,为进一步阐明MT影响绒山羊绒毛生长的分子机理提供理论依据。
1 材料与方法
1.1 试验材料
根据体重、产绒量、绒长、绒细度,选取饲养条件相同、体况良好的6只罕山型白绒山羊周岁母羊,平均分成2组,即MT埋植组(n=3)和对照组(n=3)。 试验期间,按照 2 mg/(kg·BW)的剂量在埋植组绒山羊个体耳后皮下埋植MT,对照组不埋植MT,埋植后次月于肩胛部采集1 cm2大小的皮肤样品,液氮快速冷冻带回实验室,-80℃保存备用。同一日期24时左右于静脉处采集血液样品。
1.2 试验方法
1.2.1 血液中MT含量测定:将肝素钠抗凝的新鲜血液样品2 500 r/min离心10 min(时间可根据血液分层情况适当增加),吸取上层血浆,用放免试剂盒(KIPL3300,Biosource公司)测定血浆中的MT浓度。
1.2.2 总RNA提取和文库构建:皮肤组织总RNA提取按照天根(北京)生化科技有限责任公司提供的动物组织总RNA提取试剂盒(DP501)说明书进行。利用NanoDrop ND-2000检测提取产物浓度,采用琼脂糖电泳以及Agilent 2100检测总RNA完整性。cDNA文库构建参照Illumina(USA)的链特异性转录组文库构建试剂盒(Truseq RNA Sample prep kit v2)说明书进行。
1.2.3 转录组数据分析:6个转录组文库由Illumina平台的HiSeq 2500测序系统完成高通量测序。测序原始数据使用软件Fastx_toolkit过滤接头序列,剔除低质量(质量值Q≤20的碱基数占整条reads的50%以上)以及含有 “N”(比例>2%)的reads等不合格序列。过滤后的clean reads导入FastQC做进一步的质量控制,以充分评估测序数据的质量。然后利用Tophat 2软件将clean data比对到山羊参考基因组上(capra hircus:assembly ARS1),应用cufflinks对样本的每一个基因的表达量进行归一化处理,计算FPKM值。使用cufflinks的cuffdiff模块寻找两两样本之间的差异表达基因,并通过fold_change、P-value对结果进行筛选和统计。然后利用多重假设检验(multiple hypothesis testing)对上述计算出的每一表达差异基因的P值进行校正,过滤出同时满足倍数改变以及P-value显著效应的序列进行后续分析。
对表达具有显著性差异的基因运用在线服务软件 DAVID(http://david.abcc.ncifcrf.gov/)进行 GO和KEGG pathway的富集性分析。
2 结果与分析
2.1 夜间血液中MT含量的变化
罕山绒山羊在一个绒毛生长周期内夜间血浆中MT含量的变化情况见图1。由图1可知,对照组血浆中MT水平在7~26 pg/mL,而埋植组血浆中MT平均浓度高达174.02 pg/mL,平均高出对照组10.9倍。
2.2 RNA-Seq测序结果及比对结果
选取绒毛生长兴盛前期(5月)埋植组(样品编号:MT-1、MT-2、MT-3)与对照组(样品编号:CK-1、CK-2、CK-3)皮肤样品进行转录组文库构建及测序。6个转录组文库共获得了32.03 GB的原始数据,通过质量控制最终得到了31.79 GB数据用于后续生物信息分析,平均每个样品测序数据量达到了5.3 GB。测序质量及GC含量见表1。将获得的6个样品的转录组数据通过Tophat 2比对到山羊参考基因组,序列比对结果见表2,比对成功率为(78.75±2.17)%。
2.3 差异表达基因的筛选
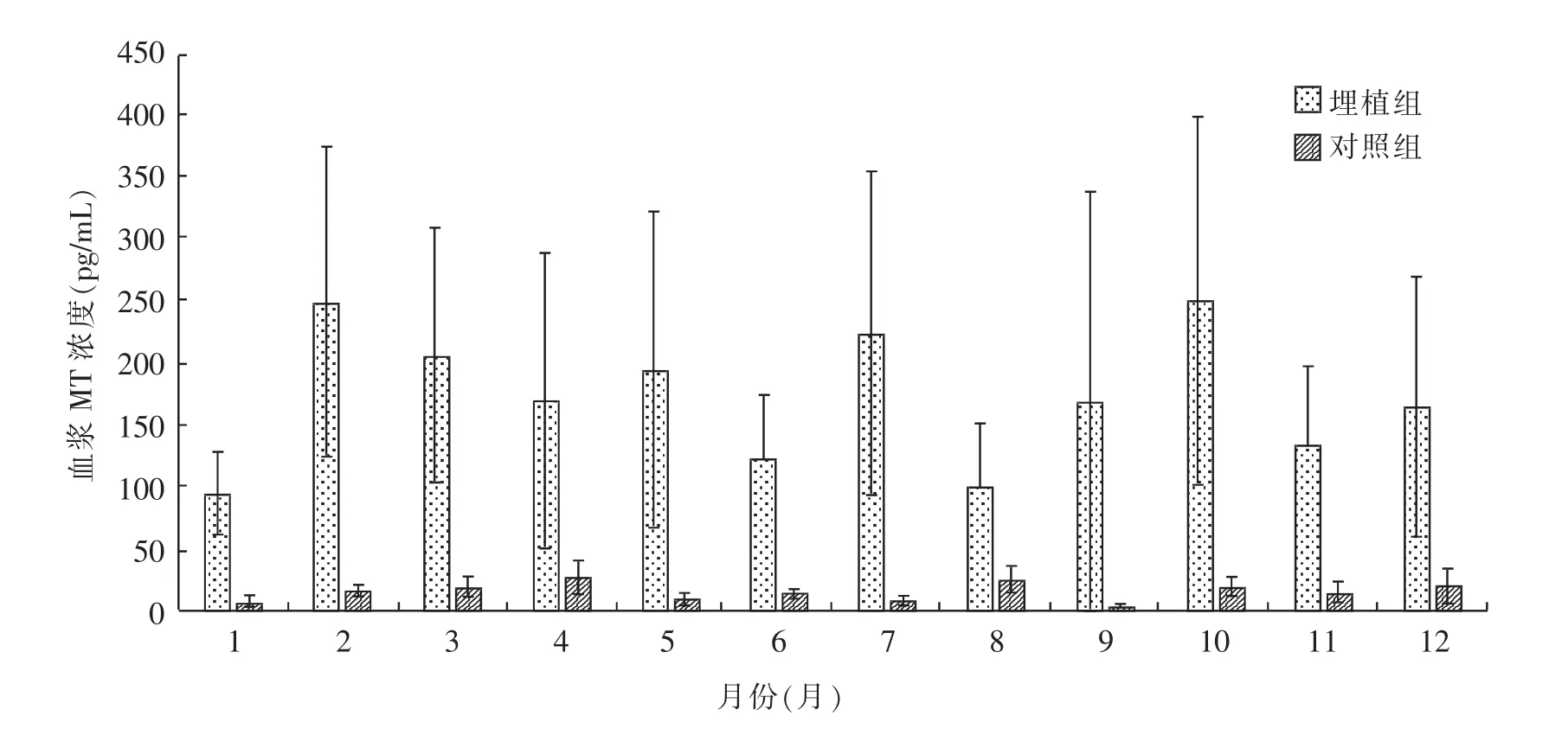
图1 埋植组与对照组绒山羊血浆MT浓度在1年内的变化趋势

表1 6个转录组文库测序结果
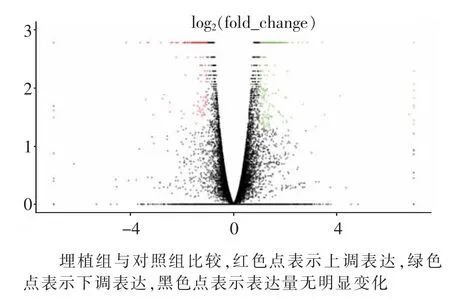
图2 基因表达量火山图
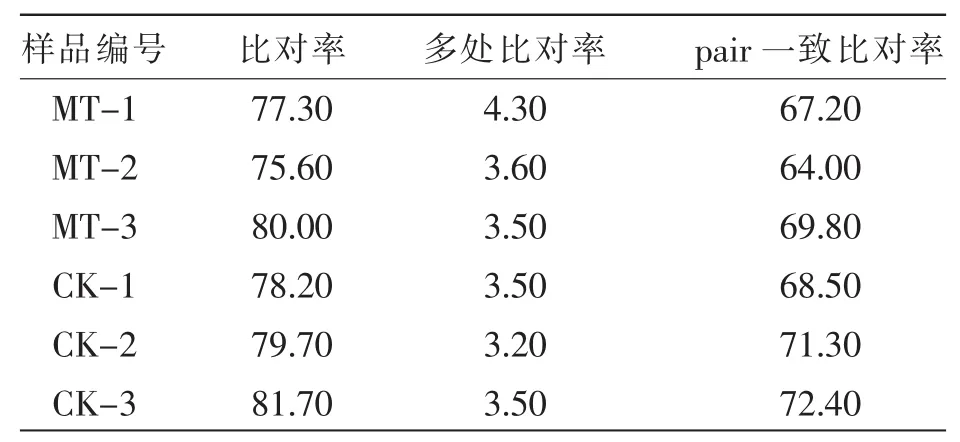
表2 测序数据比对到参考基因的结果 %
FPKM是指每一百万个比对上的片段中比对到外显子的每1 000个碱基上的片段个数,该值反映了基因的表达水平,可以用于不同样品之间基因表达差异的比较。在该研究中,埋植组表达的基因数目为14 726个(FPKM>0.2),对照组表达的基因数目为14 943个(FPKM>0.2)。通过差异基因筛选(log2|(FC)|>1,P-value<0.01),共获得了 462个差异基因,其中上调表达基因208个,下调表达基因254个(见图2)。表3列出了埋植MT以后上下调前10位的基因。
2.4 差异基因的富集分析
将上述差异基因使用在线软件DAVID进行GO及KEGG pathway的功能富集。KEGG富集分析结果如图3所示。其中,横轴代表pathway显著性水平(P-value)的-log2 P值,纵轴代表KEGG数据库中对应的pathway条目名称,条形图上的数字表示参与该pathway的差异基因数目。由图3可知,差异表达基因主要富集的信号通路为:代谢途径(metabolic pathways)、不饱和脂肪酸生物合成(biosynthesis of unsaturated fatty acids)、 脂肪酸代谢(fatty acid metabolism)、癌症信号通路(pathways in cancer)、鳞状细胞癌信号通路(squamous cell carcinoma)、黑色素合成(melanogenesis)、Wnt信号通路(Wnt signaling pathway)、补体和凝血级联反应(complement and coagulation cascades)。
3 讨论
MT对绒毛生长影响的研究由来已久。Rust等[11]报道,使用外源 MT 可促使白鼬(Mustela er-minea)冬皮提早成熟;Kim 等[12]研究发现,局部注射MT可能会影响人类头发的生长。在绒山羊绒毛周期性生长方面,研究者发现,持续埋植MT可导致山羊体表出现常年长绒现象[13-15]。内蒙古绒山羊于每年7—8月体表开始长绒,持续一年埋植MT可导致其翌年5月时体表长出夏季绒毛[13-14]。该研究从2014年12月开始对罕山型白绒山羊持续埋植MT,翌年4月脱绒后,埋植组绒山羊在5月时体表就已长绒,而对照组在7月才开始长绒。为此,选择5月的皮肤组织作为研究对象,进行RNA-Seq测序和分析。分析结果表明,埋植组绒山羊皮肤组织中表达倍数改变在2倍以上的差异基因达到了462个,其中上调表达基因208个,下调表达基因254个;经KEGG富集分析发现,差异表达基因主要富集的信号通路为:代谢途径、不饱和脂肪酸生物合成、脂肪酸代谢、癌症信号通路、鳞状细胞癌信号通路、黑色素合成、Wnt信号通路以及补体和凝血级联反应。Wnt/β-catenin信号通路作为表皮和间充质间相互作用的主要调节器,能够调节毛囊的发育[16-17],在胚胎毛囊发育过程
中可能是BMP的遗传上游通路[18]。Wnt信号通路中的 FZD10、WNT11、WNT4、PLCB1、DAAM2、WNT2B等基因在埋植MT后皮肤组织中的表达量发生了明显的变化,因此,MT可能通过调节这些基因的表达变化,进而提前激活皮肤毛囊的活性,促使绒山羊二次生绒。

表3 MT埋植后上、下调前10位的基因

图3 差异基因KEGG富集分析结果
4 结论
该研究应用生物信息学方法对6只处于皮肤毛囊发育生长起始点(5月)的罕山型白绒山羊皮肤组织的高通量测序结果进行了分析,筛选获得了462个在MT埋植组与对照组中差异表达的基因,其中,上调表达208个,下调表达254个。KEGG富集性分析表明,差异表达基因主要参与:代谢途径、不饱和脂肪酸生物合成、脂肪酸代谢、癌症信号通路、鳞状细胞癌信号通路、黑色素合成、Wnt信号通路以及补体和凝血级联反应等。Wnt信号可能在褪黑激素调控皮肤毛囊发育中发挥重要作用。
:
[1]孔庆松,刘志平,景松岩,等.褪黑激素促进水貂冬皮早熟的研究[J].野生动物,1995(4):22-26.
[2]MARTINET L,ALLAIN D,WEINER C.Role of prolactin in the photoperiodic control of moulting in the mink(Mustela visori)[J].J Endocrinol,1984,103(1):9-15.
[3]王林枫,杨改青,杨耀胜,等.光照和褪黑激素对非生绒期绒山羊激素分泌和绒毛生长的影响[J].中国畜牧兽医,2008,35(12):29-32.
[4]CONG Y Y,DENG H W,FENG Y L,et al.Melatonin implantation from wintersolstice could extend the cashmere growth phase effectively [J].Small Ruminant Research,2011,99(1):48-53.
[5]柳建昌.含褪黑素微型胶囊促使银狐毛皮提早成熟的效应[J].动物学杂志,1995,30(4):48-51.
[6]李维克.外源性褪黑激素对蓝狐、蓝霜狐及乌苏里貉毛皮性状影响的研究 [D].哈尔滨:东北林业大学,2009.
[7]KLOREN W R L,NORTON B W,WATERS M J.Fleece growth in Australian cashmere goats.Ⅰ.The effects of nutrition and age on fleece growth,prolactin and thyroxine concentration [J].Australian Journal of Agricultural Research,1993,44(5):1003-1021.
[8]程建波,张子军,章孝荣,等.褪黑激素对绒山羊绒毛生长的影响及其作用机理研究进展[J].贵州农业科学,2011,39(3):166-168.
[9]贾志海,安民.山羊绒生长机理研究进展[J].草食家畜,1996(1):2-6.
[10]COSTA V,ANGELINI C,De FEIS I,et al.Uncovering the complexity of transcriptomes with RNA-Seq [J].Journalof Biomedicine and Biotechnology,2010:853916.
[11]RUST C C,MEYER R K.Hair color,molt,and testis size in male,short-tailed weasels treated with melatonin[J].Science,1969,165(3896):921-922.
[12]KIM T K,KLESZCZYNSKI K,JANJETOVIC Z,et al.Metabolism of melatonin and biological activity of intermediates of melatoninergic pathway in human skin cells[J].FASEB Journal,2013,27(7):2742-2755.
[13]丽春.褪黑激素影响山羊绒生长的分子机理[D].呼和浩特:内蒙古农业大学,2012.
[14]常子丽.持续埋植褪黑素对绒山羊绒毛生长特性和相关基因影响的研究[D].呼和浩特:内蒙古农业大学,2010.
[15]岳春旺,张微,孔祥浩,等.褪黑激素对内蒙古白绒山羊产绒性能的影响[J].中国畜牧杂志,2007,43(7):32-34.
[16]FUCHS E.Scratching the surface of skin development[J].Nature,2007,445(7130):834-842.
[17]YU B D,MUKHOPADHYAY A,WONG C.Skin and hair:models for exploring organ regeneration[J].Human Molecular Genetics,2008(17):54-59.
[18]HUELSKEN J,VOGEL R,ERDMANN B,et al.Betacatenin controls hair follicle morphogenesis and stem cell differentiation in the skin[J].Cell,2001,105(4):533-545.
猜你喜欢
今日农业(2022年14期)2022-09-15
北方牧业(2021年16期)2021-11-29
感染、炎症、修复(2021年1期)2021-07-28
新农业(2020年5期)2020-06-01
心电与循环(2020年1期)2020-02-27
课堂内外(高中版)(2019年8期)2019-09-02
中华细胞与干细胞杂志(电子版)(2019年1期)2019-05-18
益寿宝典(2018年21期)2018-09-10
新农村(2018年35期)2018-04-02
江苏农业科学(2017年5期)2017-04-15
