
Sirt1基因多态性与有氧耐力表型的关联研究
2018-04-24 10:49:47陈彩珍何子红
中国体育科技 2018年2期
金 晶,冯 燕,卢 健,陈彩珍,何子红
Sirt1基因多态性与有氧耐力表型的关联研究
金 晶1,2,冯 燕1,卢 健1,陈彩珍1,何子红3
1.华东师范大学 体育与健康学院, 上海 200241; 2 .浙江农林大学 体军部, 浙江 杭州 311300; 3.国家体育总局体育科学研究所, 北京 100061
目的:通过分析去乙酰化酶Sirt1多态性与60名中国北方汉族女子专业长跑运动员身体机能指标的关联性,探寻预测有氧耐力表型的分子标记。方法:应用基质辅助激光解吸附电离飞行时间质谱检测技术对Sirt1基因8个SNP分型进行检测,采用联合测试法测得无氧阈和最大摄氧量相关指标,肺功能仪测得肺功能相关指标。结果:1)rs11596401位点不同基因型运动员之间肺活量存在显著性差异,TC基因型运动员显著高于TT型和CC型运动员(<0.05);2)rs12778366-rs11596401-rs4746720组合的单体型(A/B组)与有氧耐力表型(最大摄氧量/体重、无氧阈摄氧量/体重等)有极其显著性差异(<0.01);3)A组基因型的5 000 m和10 000 m成绩极其显著优于B组单体型(<0.01)。结论:1)rs11596401位点的TC基因型可作为预测中国北方汉族女子长跑运动员肺活量的分子标记;2)rs12778366-rs11596401-rs4746720组合的A组单体型可作为预测中国北方汉族女子长跑运动员无氧阈摄氧量/体重、最大摄氧量/体重等指标的分子遗传学标记;3)rs12778366-rs11596401-rs4746720组合的A组单体型可作为中国北方汉族女子5 000 m和10 000 m项目运动员选材的分子遗传学标记。
Sirt1基因多态性;有氧耐力表型;遗传学标记;表观遗传学;肺活量
运动员选材是竞技体育活动的开端,是挑选具有良好运动天赋及竞技潜力的儿童少年或后备力量参加运动训练的起始性工作。世界各国力求使用科学的测试和预测方法,努力提高选材的精准度和成功率,分子遗传学的选材近年来得到广泛认识,因此,探寻与项目特征相符的有效分子标记是运动员选材的重要方式[9]。
Sir2(silent information regulator 2)与Sirt1(silent mating type information regulation2 homolog1)同源性最高,是NAD+依赖的组蛋白去乙酰化酶,以依赖NAD+的方式使组蛋白发生去乙酰化作用,它是目前研究最深的Sirtuin家族成员[18]。Sirt1与衰老及年龄性疾病都具有相关性[24,33],拥有延长细胞寿命的“长寿作用”,文献也报道Sirt1与调控能量代谢[12]和骨骼肌氧化能力有高度相关性[44]。

综上所述,Sirt1与有氧运动能力可能存在很高的相关性,国内报告Sirt1与有氧运动的关联性研究主要集中于相关代谢疾病和动物模型中[3-6],鲜有报道Sirt1与有氧运动能力相关的人体实验研究[2,22]。本研究对Sirt1基因所有标签单核苷酸多态位点进行分型,拟通过研究优秀耐力运动员生理指标(肺活量、运动成绩、身体机能)与Sirt1的基因单核苷酸多态性的相关性分析,探讨其影响有氧耐力表型的相关性,试图探索其作为中国北方汉族女子耐力项目运动员选材遗传学标记的可能性。
1 研究对象与方法
1.1 研究对象
中国北方汉族女子专业长跑运动员,来自备战2012年奥运会的国家长跑集训队以及北京、山东、河南、河北、内蒙古的省(自治区)田径队运动员共60名(运动员等级均为一级以上,其中,国际健将15名、健将7名)。
1.2 研究方法
1.2.1 肘静脉取血
用Wizard基因组DNA纯化试剂盒(Promega)从全血中提取基因组DNA。
1.2.2 多态位点检测
Sirt1基因全长约为33.72 kb,上下游各扩展2 kb后从hapmap网站(www.hapmap.org)下载Sirt1(中国北方汉族人)的SNPs,应用Haploview软件中的Tagger程序,根据Pair-wise方法,最小2=0.8,Maf≥0.1选择出4个标签SNP-rs12778366、rs11596401、rs4746720、rs1022764。普遍认为,基因外显子的作用是编码氨基酸,从而决定蛋白质的性质。因此,添加4个发生氨基酸改变的SNP-rs2273773(leu332leu)、rs116374368(His357Gln)、rs17855430(Ala91Asp)、rs114182972(Thr712Ala)进行实验。
1.3 基因分型
依据NCBI网站公布的人类Sirt1基因序列(NM_001142498.1),PCR引物和单碱基延伸引物都是用Assay Designer(Sequenom)软件包设计。使用MassARRAY Nanodispenser将最终的分型产物点样到spectroCHIP上,并用基质辅助激光解吸电离飞行时间质谱进行分析。
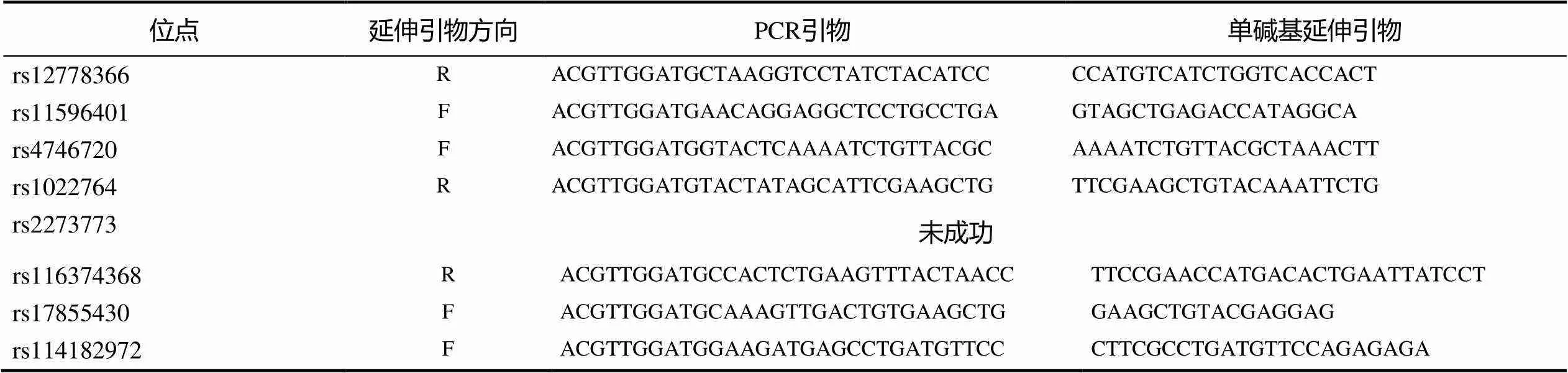
表1 Sirt1基因SNP的引物及单碱基引物信息
注:延伸引物方向F表示正向,R表示反向。
1.4 生理指标的测试方法
采集60名优秀女子长跑运动员的生理指标,包括:5 000 m成绩(s)、10 000 m成绩(s)、马拉松成绩(min)、肺活量(L)、时间肺活量(L)、1 s用力呼出量(L)、每分通气量(L/min)、无氧阈摄氧量/体重(mL/kg/min)、无氧阈心率(次/min)、无氧阈跑速(m/s)、最大摄氧量/体重(mL/kg/min)、最大心率(次/min)、最大跑速(m/s)、最大通气量(L/min)。测试仪器为德国Cortex Metalyzer 3B气体代谢分析仪、德国Cosmos跑台和芬兰Polar心率表。肺功能相关指标使用TOKYO Chestgraph HI-101肺功能仪测得。
1.5 统计学分析
所有数据处理采用SPSS 17.0软件统计包完成。采用卡方检验进行H-W平衡检验,即哈代-温伯格平衡定律(Hardy-Weinberg equilibrium),又称遗传平衡定律,是指对于一个大且随机交配的种群,基因频率和基因型频率在没有迁移、突变和选择的条件下会保持不变,>0.05说明该群体达到遗传平衡,即数据可信。所测生理指标数据先用K-S检验是否符合正态分布。K-S检验是检验单一样本是否来自某一特定分布的方法。检验方法是以样本数据的累计频数分布和特定理论分布比较,若两者间的差距很小,则推论该样本取自某特定分布族。如果符合正态分布,则基因型之间各生理指标的初始值指标采用独立样本检验或单因素方差分析,若不符合正态分布采用非参数检验处理。显著性水平设为<0.05,非常显著性水平设为<0.01。
2 结果
2.1 各位点完成检测样本数
本实验对Sirt1基因的8个SNP进行分型,结果rs114182972、rs1022764、rs2273773未能分型成功;另有rs11637436和rs17855430两个SNP,分型结果均为AA基因型,没有统计的意义;最终对rs12778366、rs11596401和rs4746720的结果进行分析(表2)。
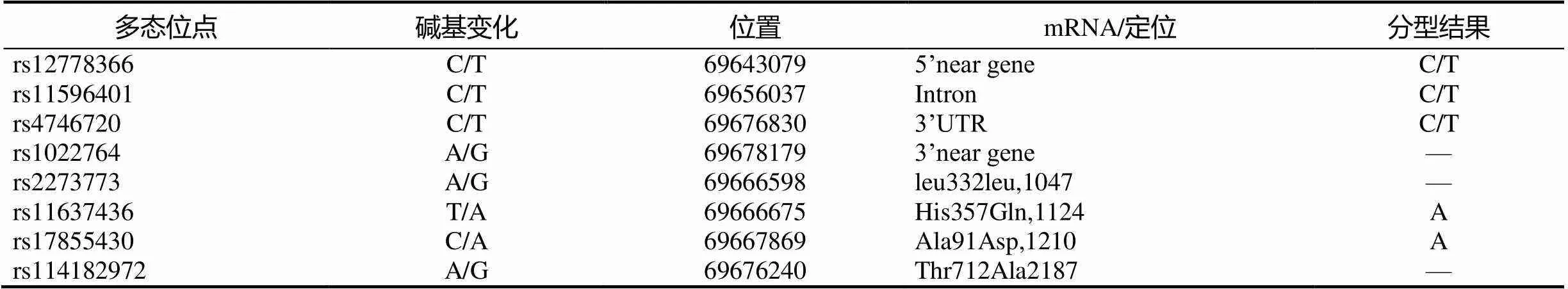
表2 Sirt1基因SNP的分型结果
2.2 Sirt1基因SNP位点与生理指标的关联性
2.2.1 Sirt1基因位点多态性与有氧耐力表型的关联分析
经独立样本检验或单因素方差分析(表3、表4、表5),rs12778366(其中检测出一个运动员为CC型,剔除处理)、rs11596401、rs4746720位点不同基因型运动员多态性与其他指标之间均无关联性(>0.05)。

表3 rs12778366多态性与有氧耐力表型的相关性
2.2.2 Sirt1基因单个位点多态性与肺功能的关联分析
经过统计分析(表6),Sirt1基因rs12778366位点不同基因型运动员之间肺功能指标(肺活量、时间肺活量、1 s用力呼出量)无显著性差异(>0.05)。
rs11596401位点不同基因型运动员之间肺活量存在显著性差异,TC基因型运动员显著高于TT型和CC型运动员(<0.05)。TC基因型的时间肺活量和1 s用力呼出量均显著高于CC型(<0.05),而与TT型没有差异性(>0.05)。
rs4746720位点分析结果显示,TT基因型的时间肺活量和1 s用力呼出量极其显著高于TC型运动员(<0.01)。
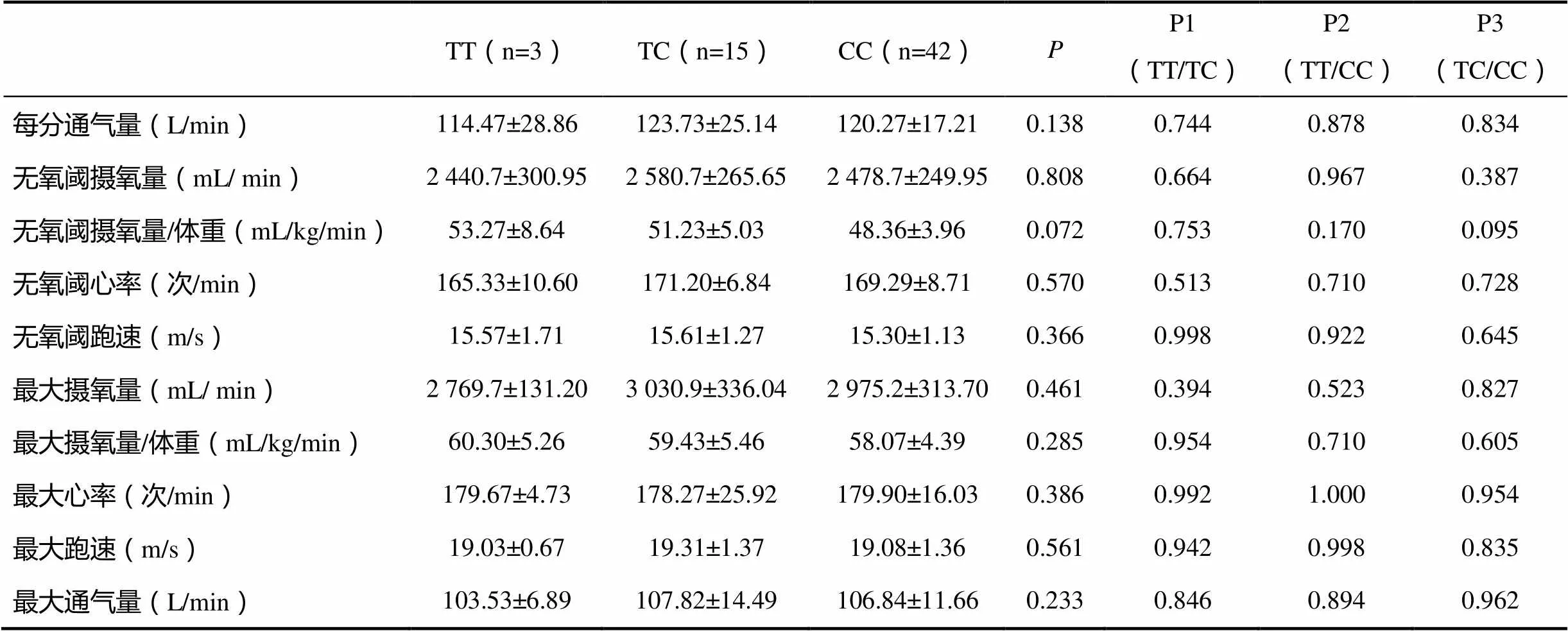
表4 rs11596401多态性与有氧耐力表型的相关性
注:P1表示TT基因型/CT基因型的相关性,P2表示TT基因型/CC基因型的相关性,P3表示CT基因型/CC基因型的相关性,下同。

表5 rs4746720多态性与有氧耐力表型的相关性

表6 Sirt1基因位点多态性与肺功能的相关性
注:*表示有显著性差异<0.05,**表示极显著性差异<0.01。
2.2.3 Sirt1基因多态性与运动成绩的关联分析
经独立样本检验或单因素方差分析(表3、表4、表5),rs12778366、rs11596401、rs4746720位点不同基因型运动员多态性与运动成绩(5 000 m、10 000 m和马拉松成绩)没有相关性(>0.05),数据略。
2.3 Sirt1单体型与机能指标的关联分析
2.3.1 Sirt1基因单体型分型结果
本文对Sirt1基因rs12778366、rs11596401、rs4746720 3个位点多态性组成的单体型在测试的60名运动员中进行分析,共存在11个单体型(表7)。数据显示,rs12778366位点的CC型个体出现的频率极低,仅分型得到1位运动员,其中3个位点都是TC型的单体型(TCTCTC)运动员未出现。
只有极少数拥有天赋的运动员才能获得杰出运动能力,因而,本文将检测样本rs12778366-rs11596401-rs4746720组合的单体型分为n<6人的单体型归为A组(单体型类型a~fn≥6人的单体型归为B组(g~k),进行分析。
2.3.2 Sirt1单体型(A/B组)与有氧耐力表型的关联性
经数理统计,本文发现,Sirt1单体型A组和B组之间与肺功能和有氧耐力表型有很强的相关性。本研究共检测13个生理指标,其中7个指标显示组间有相关性<0.05(图1 a~e)。
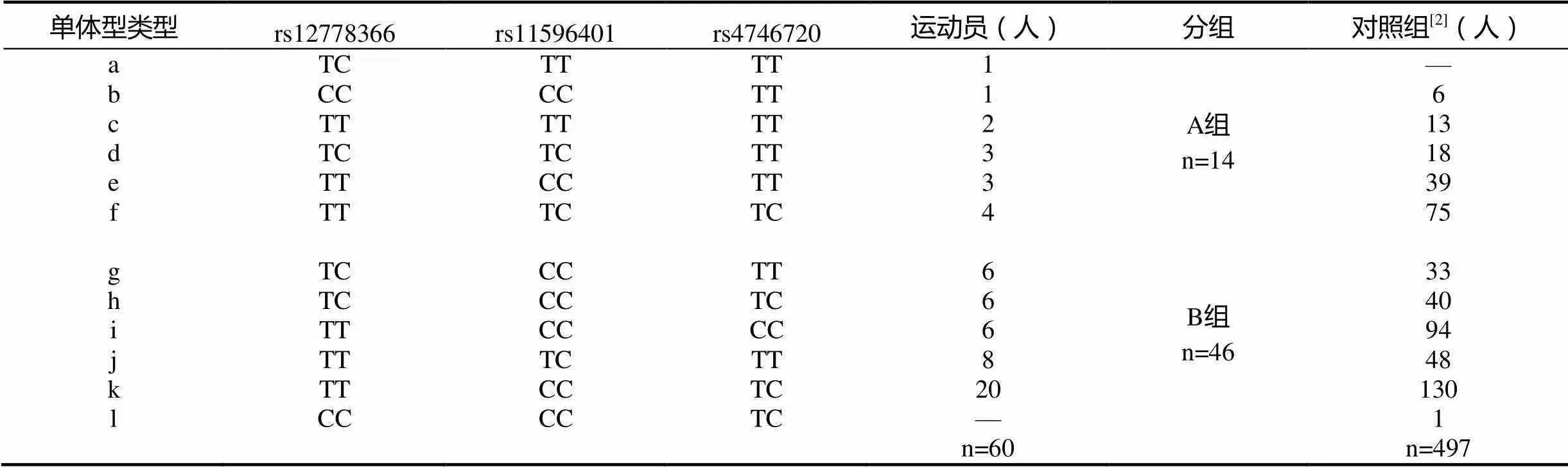
表7 Sirt1基因单体型分型结果
2.3.3 Sirt1单体型(A/B组)与运动成绩的关联性
本研究意外发现Sirt1单体型不同组间(A/B组)与有氧耐力表型的相关性后,尝试着进行与运动成绩的相关性分析。由于运动员的专长和项目不同,本研究5 000 m、 10 000 m和马拉松成绩数据的总人数不同,分别为44人、35人和21人。结果显示,A组5 000 m成绩(947.92±39.9 s)极其显著高于B组(1 012.13±55.52 s),=0.001;10 000 m成绩A组(1 995.5±87.00 s)优于B组运动员(2 107.80±121.18 s),=0.006(图1-f);马拉松成绩上两组间没有差异性(>0.05,图1-g)。
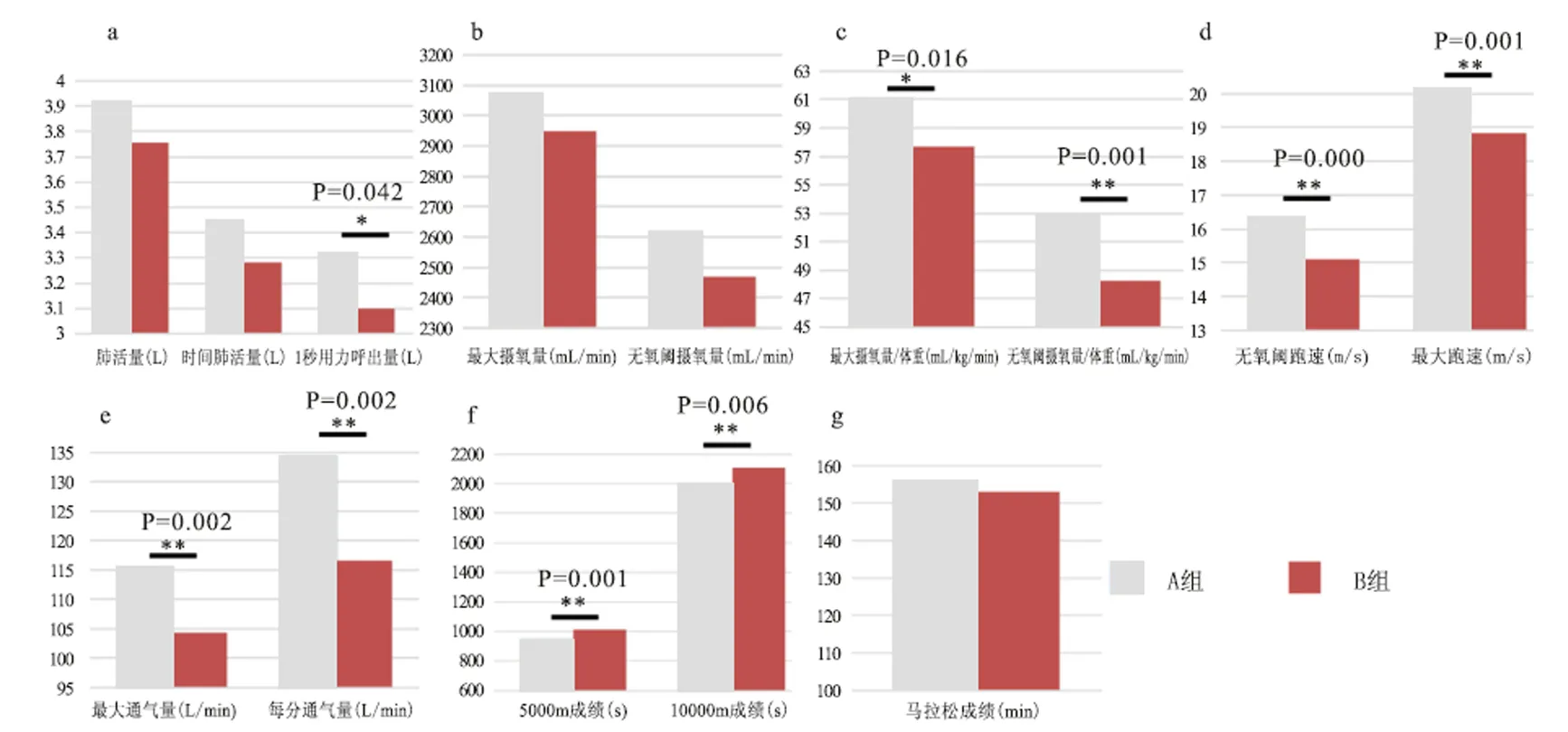
图1 Sirt1单体型(A/B组)与机能指标的相关性分析
Figure1. Correlation between Sirt1 haplotype (A/B) and Function Index
注:a:单体型与肺功能指标的相关性,A组的1 s用力呼出量显著高于B组(<0.05),其他指标没有显著性相关;b:两组间最大摄氧量和无氧阈摄氧量没有差异性;c:A/B组间最大摄氧量/体重有显著性差异(<0.05),无氧阈摄氧量/体重有极其显著型差异(<0.01);d:最大跑速和无氧阈跑速有极其显著性差异(<0.01);e:最大通气量和每分通气量在两组间有极其显著性差异=0.002;f:A组单体型的5 000 m成绩(A组n=13/B组n=31)和10 000 m成绩(A组n=13/B组n=22)极显著优于B组单体型成绩(<0.01);g:马拉松成绩两组间(A组n=11/B组n=10)没有显著性差异。*表示有显著性差异<0.05,**表示有极显著性差异<0.01。
3 讨论
检索Pubmed网站鲜有报告关于Sirt1基因的多态性与运动能力表型的关联性研究,主要集中在肥胖研究[29,36]、体脂和血压[41],长寿[24,30]和疾病[27,46]。本文报告单个多态位点与肺功能指标有关联性,虽然其他位点也显示肺功能上部分基因型的优越性,如rs4746720的TT基因型时间肺活量和1 s用力呼出量都极其显著高于TC基因型,然而,所有位点多态基因型中rs11596401的TC基因型肺活量(4.01±0.43 L)为最高参数,rs11595401位点TC型的肺活量显著高于TT基因型和CC基因型运动员(<0.05,表6),因此认为,该位点的TC基因型可作为预测中国北方汉族女子长跑运动员肺活量的分子标记。观察时间肺活量和1s用力呼出量显示rs11596401的TC基因型显著高于CC型(<0.05),但TT型与TC/CC没有差异,只能说明TC基因型在这两个指标上略高于其他基因型,并未达到统计学意义。本文统计Sirt1基因中rs12778366、rs11596401、rs4746720位点不同基因型运动员多态性与有氧运动能力和运动成绩,结果均显示没有相关性,因而可以判断,单个多态位点与有氧运动能力没有相关性。
本研究中对3个位点不同基因型组成的单倍体与有氧耐力表型进行分析,分型共得到11种单倍体(表7 a~k)。研究发现,k表示的TTCCTC基因型有20例,占据1/3的实验样本总数,猜测是否这种基因型的个体更容易拥有杰出的运动能力,即成为优秀耐力运动员。因此分析对照组(本文以文献[2]作为先期研究基础,因此不重复例举)和运动员之间进行卡方检验,并未发现k基因型个体在运动员组和对照组之间有差异性。单倍体的分型中还发现rs12778366的CC基因型出现频率较低,与之前报告的结果相一致[2,22]。观察到rs12778366的T等位基因个体分型共有11/12种,笔者对先前样本量统计未发现1例TCTCTC基因型,认为汉族人群中可能没有该基因型个体。
人类有氧运动能力受到环境与训练的敏感性在极大程度上是由遗传因素所决定的,而且越接近于训练的极限,越依赖于基因遗传表达[37]。本研究的测试者均为我国优秀女子有氧耐力运动员,假设只有极少数的个体才拥有杰出运动能力的遗传物质,因此,本文将少于6例的单倍体划为A组,反之划分为B组,对有氧耐力表型和肺活量等指标进行关联性分析。结果发现,A组单倍体基因型在最大摄氧量/体重、无氧阈摄氧量/体重、1 s用力呼出量、无氧阈跑速等有氧耐力的指标上均显著/极其显著高于B组。单体型是核苷酸特定的组合形式,较单一的多态位点含有更多的SNPs的遗传信息,与复杂形状的相关分析后,单体型分析比单个位点分析具有更高的统计效能,是目前认为比较有效的多位点联合分析方法,特别适用于多基因调控表型的研究[45]。
图1b和c分别显示了最大摄氧量和无氧阈摄氧量的绝对值和相对值,研究报告最大摄氧量在A/B组之间没有差异性(>0.05),最大摄氧量/体重和无氧阈摄氧量/体重组间有极其显著的差异(<0.01)。最大摄氧量的结果与Howley等研究相一致,在训练的初始阶段,最大摄氧量与成绩呈线性正相关,在高水平的运动群体这种平行关系消失[23],本研究对象均为我国高水平长跑运动员,所以之间没有相关性。最大摄氧量的利用率是耐力运动员取得成功的关键因素之一[26],最大摄氧量与体重成正相关,最大摄氧量的相对值(最大摄氧量/体重)更能体现运动员的有氧能力(A组相对值显著高于B组运动员),无氧阈摄氧量/体重指标亦然。因此认为,rs12778366-rs11596401-rs4746720组合的A组单体型可作为预测中国北方汉族女子长跑运动员无氧阈摄氧量/体重、最大摄氧量/体重等指标的分子遗传学标记。
本文测试的13个指标中有7个指标均出现显著性,显示A组中的单体型基因型可能具有较高的有氧耐力表型,可能可以作为预测有氧耐力运动员的分子标记。为了探究这种可能性,本文试探性的分析A/B组单体型基因型与运动成绩之间是否存在关联性。结果发现,5 000 m和10 000 m成绩,A组单体型成绩极其显著高于B组(<0.01),与有氧耐力表型中出现的有利因素相一致(图1)。马拉松成绩没有差异性,马拉松运动是人类挑战极限的运动,除了对身体素质和技能有高要求外,其成绩还受到外界环境影响[17]。另外,将A组基因型在运动员组和对照组进行卡方检验没有发现有显著性的分布差异,也就是说,在运动员组和对照组中A组单体型分布情况基本相似。换句话说,A组中的单体型个体在人群中的分布没有差异性,而经过训练后更有利于获得训练的敏感性和潜能(遗传基因)的激发,进而使其获得较高的生理机能和运动成绩(5 000 m和10 000 m),成为更为“优秀”的长跑运动员。综上所述,拥有较高有氧耐力表型的基因型(A组个体)同时也具有较好的运动成绩,因此本研究认为,A组(表7a~f)单倍体可作为我国汉族女子长跑运动员选材的基因标志。
研究的结果虽然是“偶然”的发现,但也并不“意外”。1)人作为生物有机体的存在,无时无刻不在进行新陈代谢(氧化还原反应),Sirt1通过诱导抗氧化酶,如超氧化物歧化酶和过氧化氢等,调控氧化应激反应[16],Sirt1基因与人群长寿的相关性已经被证实[24, 33],间接的说明Sirt1调控长寿ROS(活性氧)的侵扰与增强氧化能力的关联[32]。2)在骨骼肌细胞中,Sirt1主导NAD+的含量并调控氧化应激反应[32]。运动时通过激活AMPK,进而大量激发Nampt[21],运动员具有较高水平的Nampt且提示与有氧耐力的重要指标最大摄氧量有相关性[13],不同基因型的单体型有不同Sirt1基因的转录和表达水平,可以解释A/B组中出现有氧表型(最大摄氧量/体重、无氧阈摄氧量/体重等)现象。3)Sirt1基因的去乙酰化作用在骨骼肌中的表观遗传学的研究热点[38],有氧耐力表型是否可通过表观遗传的方式传递有待考证。但许多实验已经证明Sirt1能促进有氧运动能力:槲皮黄酮是Sirt1的诱导因子[31],经过槲皮黄酮的喂养和跑台训练,结果显示,Sirt1和PGC-α的mRNA高表达,且短期喂食的小鼠运动能力提升,增加运动时间约36%[14];Suwa等对小鼠进行不同强度的14天跑台训练,比目鱼肌和跖肌中发现Sirt1的表达提高[42];马拉松训练后Sirt1的mRNA含量升高,依赖于Sirt1的转录因子如FOXO3a和目标基因GADD45的mRNA也升高[35]。4)在有氧能力的基因组学研究实验中发现,有许多基因都与最大摄氧量具有相关性[28,43],Bouchard C等[8]应用22对常染色体的324611SNPs进行关联研究,结果发现,39个SNPs与△最大摄氧量关联,并且报道最强关联位点是rs6552828,其位于乙酰辅酶A合成酶长链1的第一内含子上。乙酰辅酶A是三羧酸循环中重要的原料,循环中合成α酮戊二酸和琥珀酸过程大量消耗NAD+[38],Sirt1是依赖于NAD+其也参与线粒体氧化反应[47]。综上所述,Sirt1与有氧能力有很强烈的相关性,所以判断Sirt1在有氧运动能力可能有很大遗传度,因此,本文得到的结果也具有“必然性”。
当今世界竞技体育的水平已经日益接近人类的生理极限,只有极具天赋的运动员经过系统科学的训练才会达到世界顶尖水平。如在运动选材中增加基因检测的方法,能够提高运动员成功的机率[9]。基因多态性与杰出运动能力的关联性始终受到样本量稀缺的困扰,而对于有氧能力与基因多态性的相关性研究,主要是通过Meta数理分析等方法[20]和年度总结阳性结果报道[34,40]。本文所涉及的样本为我国汉族优秀女子耐力运动员,样本量的稀少是存在的现实问题,如果盲目扩大样本量(降低纳入标准或增加其他人种的数据)将影响结果的可靠性,今后仍需经过几批人甚至几代人坚持不懈的努力,不断完善和丰富我国优秀长跑运动员基因库[1]。本文显示,Sirt1与有氧耐力存在相关性,另有文献对骨骼肌卫星细胞进行重塑过程中不同阶段(激活、增殖、分化)的乙酰化水平进行测定,发现有很大的差异性[39],因此,认为去乙酰化作用是具有时相性。为了进一步证实Sirt1对有氧耐力表型的作用,需要对有氧运动过程中各阶段Sirt1基因转录和表达水平进行实时检测。
4 结论
1. rs11596401位点的TC基因型可作为预测中国北方汉族女子长跑运动员肺活量的分子标记。
2. rs12778366-rs11596401-rs4746720组合的A组单体型可作为预测中国北方汉族女子长跑运动员无氧阈摄氧量/体重、最大摄氧量/体重等指标的分子遗传学标记。
3. rs12778366-rs11596401-rs4746720组合的A组单体型可作为中国北方汉族女子5 000 m和10 000 m项目运动员选材的分子遗传学标记。
[1] 胡扬. 基因多态性和竞技体育金牌战略[J]. 体育科学, 2013, 33(12): 14-15.
[2] 金晶,何子红,胡扬,等. ERRα和SIRT1基因多态性位点在优秀耐力运动员中的分布特征及其功能研究[J]. 中国运动医学杂志, 2013, 32(10): 868-874.
[3] 林琴琴,耿元文,田振军. 间歇有氧运动激活miR-21/SIRT1/NF-κB通路改善心梗大鼠肾功能研究[J]. 体育科学, 2017, 37(7): 44-49, 82.
[4] 燕小妮. SIRT1在有氧运动改善APOE--/小鼠肝脏脂代谢异常中的作用研究[J]. 体育科学, 2015,35(8): 40-44.
[5] 张坦,崔迪,张喆,等. 游泳运动对糖尿病小鼠骨骼肌AMPK/ SIRT1/NF-κB炎症信号通路的影响[J]. 体育科学,2016,36(9):40-47.
[6] 赵军,赵美琴,史长征,等. 基于SIRT1轴探讨不同强度有氧运动对NAFLD大鼠脂肪性肝炎的影响[J]. 体育科学,2014,34(11): 50-59, 96.
[7] BAKHTIARI N, HOSSEINKHANI S, SOLEIMANI M,. Short-term ursolic acid promotes skeletal muscle rejuvenation through enhancing of SIRT1 expression and satellite cells proliferation[J]. Biomed Pharmacother, 2016, 78: 185-196.
[8] BOUCHARD C, SARZYNSKI M A, RICE T K,. Genomic predictors of the maximal O(2) uptake response to standardized exercise training programs[J]. J Appl Physiol (1985), 2011, 110(5): 1160-1170.
[9] BROOKS M A, TARINI B A. Genetic testing and youth sports[J]. JAMA, 2011, 305(10): 1033-1034.
[10] CANTO C, GERHART-HINES Z, FEIGE J N,. AMPK regulates energy expenditure by modulating NAD+ metabolism and SIRT1 activity[J]. Nature, 2009, 458(7241): 1056-1060.
[11] CANTO C, JIANG L Q, DESHMUKH A S,. Interdepende-nce of AMPK and SIRT1 for metabolic adaptation to fasting and exercise in skeletal muscle[J]. Cell Metab, 2010, 11(3): 213-219.
[12] CANTO C, MENZIES K J, AUWERX J. NAD(+) metabolism and the control of energy homeostasis: A balancing act between mitochondria and the nucleus[J]. Cell Metab, 2015, 22(1): 31-53.
[13] COSTFORD S R, BAJPEYI S, PASARICA M,. Skeletal muscle NAMPT is induced by exercise in humans[J]. Am J Physiol Endocrinol Metab, 2010, 298(1): E117-E126.
[14] DAVIS J M, MURPHY E A, CARMICHAEL M D,. Quercetin increases brain and muscle mitochondrial biogenesis and exercise tolerance[J]. Am J Physiol Regul Integr Comp Physiol, 2009, 296(4): R1071-R1077.
[15] DIAZ-RUIZ A, GONZALEZ-FREIRE M, FERRUCCI L,. SIRT1 synchs satellite cell metabolism with stem cell fate[J]. Cell Stem Cell, 2015, 16(2): 103-104.
[16] DONATO A J, MORGAN R G, WALKER A E,. Cellular and molecular biology of aging endothelial cells[J]. J Mol Cell Cardiol, 2015, 89(Pt B): 122-135.
[17] EL H N, TAFFLET M, BERTHELOT G,. Impact of environmental parameters on marathon running performance[J]. PLoS One, 2012, 7(5): e37407.
[18] FULCO M, SCHILTZ R L, IEZZI S,. Sir2 regulates skeletal muscle differentiation as a potential sensor of the redox state[J]. Mol Cell, 2003, 12(1): 51-62.
[19] GERHART-HINES Z, RODGERS J T, BARE O,. Metabolic control of muscle mitochondrial function and fatty acid oxidation through SIRT1/PGC-1alpha[J]. EMBO J, 2007,26(7):1913-1923.
[20] GRAFF M, SCOTT R A, JUSTICE A E,. Genome-wide physical activity interactions in adiposity - A meta-analysis of 200,452 adults[J]. PLoS Genet, 2017, 13(4): e1006528.
[21] HARDIE D G. AMP-activated/SNF1 protein kinases: conserved guardians of cellular energy[J]. Nat Rev Mol Cell Biol,2007,8(10):774-785.
[22] HE Z H, HU Y, LI Y C,. PGC-related gene variants and elite endurance athletic status in a Chinese cohort: a functional study[J]. Scand J Med Sci Sports, 2015, 25(2): 184-195.
[23] HOWLEY E T, BASSETT D J, WELCH H G. Criteria for maximal oxygen uptake: review and commentary[J]. Med Sci Sports Exerc, 1995,27(9):1292-1301.
[24] HUANG J, SUN L, LIU M,. Association between SIRT1 gene polymorphisms and longevity of populations from Yongfu region of Guangxi.[J]. Zhonghua Yi Xue Yi Chuan Xue Za Zhi, 2013,30(1):55-59.
[25] IMAI S. The NAD world: a new systemic regulatory network for metabolism and aging--Sirt1, systemic NAD biosynthesis, and their importance[J]. Cell Biochem Biophys,2009,53(2):65-74.
[26] JOYNER M J, COYLE E F. Endurance exercise performance: the physiology of champions[J]. J Physiol,2008,586(1):35-44.
[27] KALEMCI S, EDGUNLU T G, KARA M,. Sirtuin gene polymorphisms are associated with chronic obstructive pulmonary disease in patients in Mugla province[J]. Kardiochir Torakochirur-gia Pol,2014, 11(3): 306-310.
[28] KELLER P, VOLLAARD N B, GUSTAFSSON T,. A transc-riptional map of the impact of endurance exercise training on skeletal muscle phenotype[J]. J Appl Physiol (1985),2011,110(1): 46-59.
[29] KILIC U, GOK O, ELIBOL-CAN B,. SIRT1 gene variants are related to risk of childhood obesity[J]. Eur J Pediatr,2015, 174(4):473-479.
[30] KILIC U, GOK O, ERENBERK U,. A remarkable age-related increase in SIRT1 protein expression against oxidative stress in elderly: SIRT1 gene variants and longevity in human[J]. PLoS One, 2015, 10(3): e117954.
[31] LAPPALAINEN Z. Sirtuins: a family of proteins with implica-tions for human performance and exercise physiology[J]. Res Sports Med, 2011, 19(1): 53-65.
[32] LE MOAL E, PIALOUX V, JUBAN G,. Redox control of skeletal muscle regeneration[J]. Antioxid Redox Signal,2017,27 (5):276..
[33] LIN R, YAN D, ZHANG Y,. Common variants in SIRT1 and human longevity in a Chinese population[J]. BMC Med Genet, 2016, 17: 31.
[34] LOOS R J, HAGBERG J M, PERUSSE L,. Advances in exercise, fitness, and performance genomics in 2014[J]. Med Sci Sports Exerc, 2015, 47(6): 1105-1112.
[35] MARFE G, TAFANI M, PUCCI B,. The effect of marathon on mRNA expression of anti-apoptotic and pro-apoptotic proteins and sirtuins family in male recreational long-distance runners[J]. BMC Physiol,2010,10:7.
[36] PEETERS A V, BECKERS S, VERRIJKEN A,. Association of SIRT1 gene variation with visceral obesity[J]. Hum Genet, 2008, 124(4): 431-436.
[37] PRUD'HOMME D, BOUCHARD C, LEBLANC C,. Sensit-ivity of maximal aerobic power to training is genotype-dependent[J]. Med Sci Sports Exerc,1984,16(5):489-493.
[38] RYALL J G, CLIFF T, DALTON S,. Metabolic reprogram-ming of stem cell epigenetics[J]. Cell Stem Cell,2015,17(6):651-662.
[39] RYALL J G, DELL'ORSO S, DERFOUL A,. The NAD(+)-dependent SIRT1 deacetylase translates a metabolic switch into regulatory epigenetics in skeletal muscle stem cells[J]. Cell Stem Cell, 2015, 16(2): 171-183.
[40] SARZYNSKI M A, LOOS R J, LUCIA A,. Advances in Exercise, Fitness, and Performance Genomics in 2015[J]. Med Sci Sports Exerc, 2016, 48(10): 1906-1916.
[41] SHIMOYAMA Y, SUZUKI K, HAMAJIMA N,. Sirtuin 1 gene polymorphisms are associated with body fat and blood pressure in Japanese[J]. Transl Res, 2011, 157(6): 339-347.
[42] SUWA M, NAKANO H, RADAK Z,. Endurance exercise increases the SIRT1 and peroxisome proliferator-activated recept-or gamma coactivator-1alpha protein expressions in rat skeletal muscle[J]. Metabolism, 2008, 57(7): 986-998.
[43] TIMMONS J A, KNUDSEN S, RANKINEN T,. Using molecular classification to predict gains in maximal aerobic capacity following endurance exercise training in humans[J]. J Appl Physiol (1985),2010, 108(6):1487-1496.
[44] TONKIN J, VILLARROYA F, PURI P L,. SIRT1 signaling as potential modulator of skeletal muscle diseases[J]. Curr Opin Pharmacol,2012,12(3):372-376.
[45] TRIPODI G, MODICA R, STELLA A,. Haplotype analysis of carnitine transporters and left ventricular mass in human essential hypertension[J]. J Ren Nutr, 2005, 15(1): 2-7.
[46] WANG Y, YAN X Y, WU J C,. Analysis of silent information regulator 1 (SIRT1) gene polymorphisms in antituberculosis- drug-induced hepatotoxicity in a prospective cohort study[J]. Int J Clin Pharmacol Ther,2016,54(10):775-781.
[47] ZSCHOERNIG B, MAHLKNECHT U. Sirtuin1:regulating the regulator[J].Biochem Biophys Res Commun,2008,376(2):251-255.
Association Study of Sirt1 Genes Polymorphisms with Aerobic Performance Phenotypes
JIN Jing1,2, FENG Yan1, LU Jian1, CHEN Cai-zhen1, HE Zi-hong3
1.East China Normal University, Shanghai 200241, China; 2.Zhejiang Agricultrure and Forestry University, Hangzhou 311300, China; 3.China Institute of Sport Science, Beijing 100061, China.
Objective: This study aimed to explore the genetic markers for aerobic phenotypes by analyzing the relationship between Sirt1 gene polymorphisms and physical performance in 60 female long-distance runners of China Han nationality. Methods: Application of matrix assisted laser desorption ionization time of flight mass spectrometry detection technology to test the Sirt1 gene eight SNPs. The anaerobic threshold and maximal oxygen intake were measured by joint test, pulmonary function indexes were tested by pulmonary function meter. Results: 1)significant difference between different genotypes of athletes (rs 11596401) (<0.05), they were higher in TC genotype athletes than those in CC and TT genotype athletes.2)rs12778366 - rs11596401 - rs4746720 group A/B haplotype was extremely significant difference between aerobic endurance phenotype (Maximal oxygen uptake / body weight, anaerobic threshold oxygen uptake / body weight) (<0.01). 3)The 5000m and 10000m scores of genotype A were significantly better than those of B group haplotypes (<0.01). Conclusion: 1)The TC genotype of rs11596401 can be used as a molecular marker to predict the vital capacity of Chinese female long distance running athletes in northern China. 2)rs12778366-rs11596401-rs4746720 group A haplotype, that they can be predicted that China's northern Han Chinese woman runner anaerobic threshold of oxygen uptake/body weight, VO2Maxweight/body weight index of molecular genetic markers. 3)rs12778366 - rs11596401 - rs4746720 group A haplotype can be predicted that molecular genetic markers for athletes selection of 5000m and 10000m in northern Chinese Han women.
G804.2
A
1002-9862(2018)02-0073-07
10.16470/j.csst.201802009
2017-10-24;
2018-02-11
国家体育总局体育科学研究所基本业务费(16-01)
金晶,男,讲师,在读博士研究生,主要研究方向为运动与衰老, Email:jinjing12345@163.com。
猜你喜欢
中老年保健(2022年3期)2022-11-21 09:40:36
中老年保健(2022年2期)2022-08-24 03:21:54
中老年保健(2022年4期)2022-08-22 03:01:18
汽车实用技术(2022年7期)2022-04-20 11:45:06
现代园艺(2017年21期)2018-01-03 06:41:32
中国康复理论与实践(2015年10期)2015-12-24 05:42:44
医学研究杂志(2015年5期)2015-06-10 06:43:26
中国体育教练员(2015年2期)2015-04-16 04:57:43
河南科技(2015年3期)2015-02-27 14:20:52
现代检验医学杂志(2015年5期)2015-02-06 01:42:20
