
绢蒿荒漠植物种子大小、形状变异及其生态适应特征
2018-04-18 07:48靳瑰丽鲁为华王树林陈乙实汪传建马春晖
草业学报 2018年4期
靳瑰丽,鲁为华,王树林,陈乙实,汪传建,马春晖
(1.新疆农业大学草业与环境科学学院,新疆草地资源与生态重点实验室,新疆 乌鲁木齐 830052;2.石河子大学动物科技学院,新疆 石河子 832003;3.石河子大学信息科学与技术学院,新疆 石河子 832003)
种子是植物进行繁殖更新的物质基础,是植物生活史的关键节点,而种子大小、形状是植物生活史中选择的焦点。种子的大小和形状影响其扩散、萌发、幼苗定居及种群分布。在植物性状中,种子大小和形状处于中心地位,是植物生活史中的核心特征[1]。因此,有关植物种子大小、形状变异及其生态学意义成为国内外的研究热点。国外研究学者已经在种子大小、形状和土壤种子库类型之间的关系[2],种子大小在不同分类水平上的差异,在进化方向上种子大小的变异特征[3]以及种子大小在特殊生境条件下的生态适应意义[4]等方面作了大量的研究工作。随着分子生物学技术的发展,有关种子大小、形状变异内在的分子机制方面也成为目前研究的重点内容,并取得了众多成果[5]。国内关于种子大小变异的研究已经引起生态学家的广泛关注,区域性的特殊生境条件下的种子大小变异特征成为国内学者广泛关注的焦点,主要集中在典型草原[6]、青藏高原[7]、沙地[8]、西北荒漠区[9],上述工作取得了众多成果并极大地丰富了有关种子大小变异的研究内容,并仍然是今后的研究热点。但由于生态系统种类多样,而种子大小变异由植物本身内部的遗传特性和外部环境条件所决定,因此,局部或区域性的研究结果致使种子大小的变异类型和内部发生机制(如种子大小与植物系统发育的关系,种子大小与植物生活型的关系以及进化意义)一直存在争议[10-11]。因此,在多尺度、多对象上对不同生态系统中的植物种子大小变异进行研究并进一步整合,从而得到更具有普遍意义的结果是将来的研究方向。
绢蒿(Seriphidium)荒漠草地广泛分布于天山北坡土质、沙质及盐土荒漠,是组成天山北坡垂直带的植被之一,新疆分布面积约为114.25万hm2[12]。绢蒿荒漠草地上的各种植物在长期适应干旱气候的过程中,形成了特异性的生活史对策,其中表现最为突出的就是种子性状(seed traits)多样性,这种多样性又体现在形态学和生理学特征两个方面。物种内在形态上存在由于结实位置不同而造成的多型性,不同物种间存在复杂的附属结构造成的多样性。而上述有关形态学特征以及由其主导下的生理学特征多样性是植物长期适应环境的结果,又决定着植物在特定环境条件下的生活史特征,因此,有关绢蒿荒漠植物种子性状对环境条件的适应性成为目前研究的热点[13]。但就目前的研究来看,所关注的重点在于干旱环境条件下种子特殊性状的生态学意义方面,而有关该区域内种子性状在进化水平上的变异特征仍然比较缺乏。
基于此,本研究对绢蒿荒漠主要植物种子的大小、形状进行了比较,从传统分类和基于系统进化比较两个方面分析种子大小和形状的变异特征,并重点分析荒漠干旱多变以及长期放牧环境条件下植物种子进化出的特殊的适应对策,旨在了解绢蒿荒漠区植物种子的形态特征、生态功能,从植被恢复潜力的角度探讨绢蒿荒漠植物种子对特殊生境的生态适应策略,为该类草地的利用保护以及恢复措施的制定提供依据。
1 材料与方法
1.1 研究区概况
研究区位于天山北坡中段山前丘陵及平原荒漠区,是北疆的绢蒿荒漠集中分布区,本区属于温带荒漠干旱气候,年平均降水量为90~180 mm,土壤为淡栗钙土、沙质土以及盐漠土,年均温度为5~7 ℃。植被组成主要以绢蒿为主体,另广泛伴生有藜科、十字花科、百合科、禾本科、牻牛儿苗科等多年生、一年生、短命和类短命植物,植被组成结构相对复杂且存在季节动态变化。
1.2 研究方法
于2014-2015年5-11月进行各类植物成熟种子的采集,以植物繁殖体扩散单元作为完整的采集对象,包括各植物种子和附属结构。同时,在采集过程中关注种子的多型性现象,将一种植物的多型性种子按形态学特征进行分别采集、详细记录并作为不同种对待。各类种子采集完成后带回室内阴干至恒重,将其整理装入褐色牛皮纸信封内保存于4 ℃冰箱内待用。
种子重量测定时,将每个物种的种子混合在一起,从中随机选择3份100粒发育良好的种子样品,并用1/10000的电子天平称重,并换算为单粒重(mg)。利用体视显微镜对种子三维(长、宽和高)指标进行量算,量算时随机选出3粒种子测定长、宽和高,并计算平均值,个别种子高度在体视显微镜下不便量算,利用电子游标卡尺进行量取。测定上述指标时,将某些种子的附属结构(如长芒、冠毛等)去除。将种子按照其单粒重的所在数量级可分为4类:A型(0.01~0.10 mg)、B型(0.10~1.00 mg)、C型(1.00~10.00 mg)和D型(10.00~50.00 mg)。种子三维指标测定完成后,按照Thompson等[2]介绍的方法衡量繁殖体的形状,即将种子形状与球体形状对比,求繁殖体长、宽、高的总体方差,用繁殖体三维方差衡量繁殖体的形状。方差越小,繁殖体越接近圆球形;方差越大,繁殖体越细长或扁平。计算方差前进行数据转换,即先将繁殖体长定为1,然后求出其宽、高与长的比值,用繁殖体长、宽和高的方差来衡量繁殖体的大小,得出种子形状指数。
1.3 数据分析
植物系统进化树采用国际通用的APGIII分类系统,使用Phylomatic 3.0网络版进行绘制,该结果包括各物种在进化水平上的位置及进化枝长。利用Phylocom中的AOT模块对进化树上各节点的进化保守性分别进行检测。为进一步明确种子性状进化是否存在明显的谱系信号,在系统进化树绘制结果的基础上,利用K值法来衡量连续型功能性状的系统发育信号的强度,检测功能性状与物种进化历史的相关性。其计算方法如下:计算系统发育树末端物种性状数据的均方误差(MSEO)与用系统发育树的方差-协方差矩阵(variance-covariance matrix)计算的均方误差(MSE)的比值, 实际的MSEO/MSE相对于其期望值的比值即为K值[14]。如果K值大于1,则表示该功能性状表现出比按布朗运动模型(Brownian motion model)方式进化更强的系统发育信号;如果K值小于1,表示功能性状表现出比按布朗运动模型方式进化更弱的系统发育信号。本研究中的K值及其显著性检验采用R软件包Picante的phylosignal函数计算[15]。用SPSS 20.0软件进行方差分析,多重比较采用Duncan法。
2 结果与分析
2.1 种子大小分布与生活型频度分布
种子的大小具有较大的变化范围,涵盖A、B、C和D 4个级别。按种子大小数量级分类来看,C型种子最多(46种),占总物种数的52.27%,其次是B型种子(32种),占36.36%,此外A型种子4种,占4.54%,D型种子6种,占6.82%(图1)。
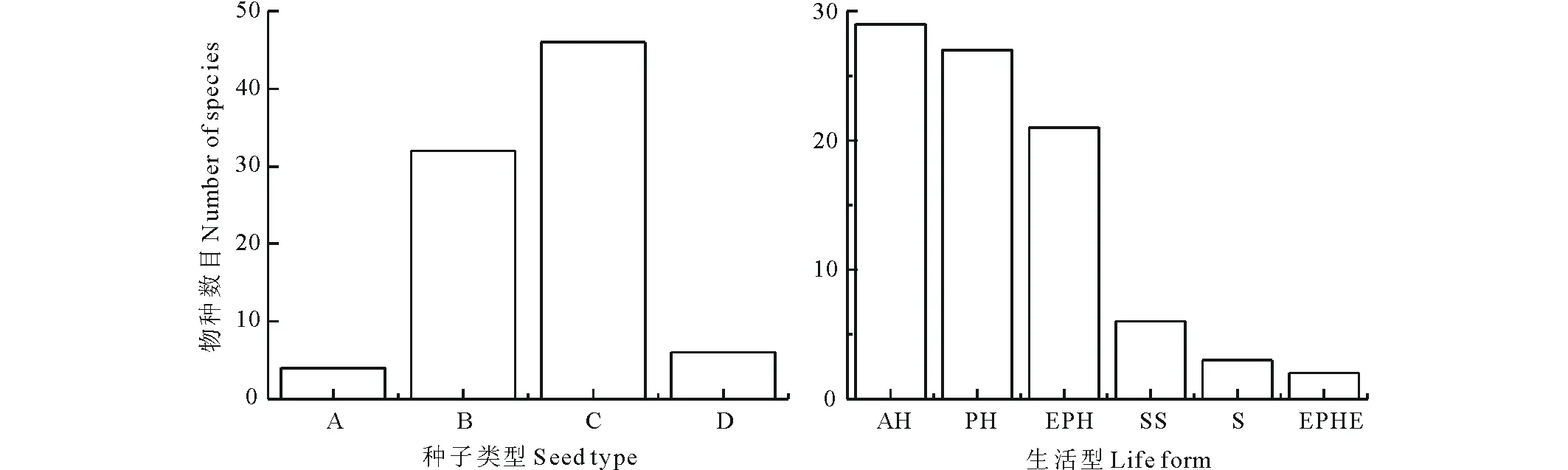
图1 不同种子大小数量级及生活型中的物种数目Fig.1 The number of species in different seed mass ranges and life form among 88 species AH: 一年生草本Annual herb; PH: 多年生草本Perennial herb; EPH: 短命植物Ephemeral plant; SS: 半灌木Semi-shrub; S: 灌木Shrub; EPHE: 类短命植物Ephemeroid plant.
所采集种子可分为6种生活型,其中一年生草本(AH)最多,达到29种,占总物种数的32.95%,其次是多年生草本(PH)和短命植物(EPH),分别为27和21种,分别占总物种数的30.68%和23.86%,半灌木(SS)6种,占总物种数的6.82%,灌木(S)3种,占总物种数的3.41%,类短命植物(EPHE)2种,占总物种数的2.27%(图1)。
2.2 种子大小和形状在不同分类水平上的变异性
实地采集的88种植物种子,分别属于12科64属。其中藜科24种,菊科20种,禾本科14种,十字花科13种,豆科7种,紫草科3种,百合科2种,蒺藜科、小檗科、石蒜科、莎草科和车前科各1种(表1)。
种子形状指数为0.00~0.20,细长或扁平种子有42种,球形或接近球形种子有46种。其中豆科、藜科、蒺藜科和小檗科种子多为圆球形,而禾本科、菊科以及车前科多为细长或扁平形(表1)。
种子大小统计数据显示,各科间的种子大小表现出极显著差异(F=3.41,P<0.01)。其中种子单粒较重的5个科是小檗科(19.72 mg)、豆科(8.16 mg)、石蒜科(4.86 mg)、百合科(2.22 mg)、车前科(2.06 mg)。单粒重最小的5个科是紫草科(1.01 mg)、菊科(1.03 mg)、禾本科(1.19 mg)、十字花科(1.40 mg)和藜科(1.88 mg)(表1)。
植物种类较多的科,其种间种子大小差异极显著,其中十字花科(F=2143.01,P<0.01),禾本科(F=7076.42,P<0.01),藜科(F=1343.23,P<0.01),菊科(F=29981.11,P<0.01),豆科(F=9731.21,P<0.01),紫草科(F=7542.34,P<0.01)。
对同种植物异型种子大小进行单因素方差分析,结果表明,异果芥(F=878.9,P<0.01)、紫翅猪毛菜(F=2179.2,P<0.01)、角果藜(F=63.3,P<0.01)、野榆钱菠菜(F=25477.9,P<0.01)、异苞滨藜(F=118960.10,P<0.01)、异子蓬(F=25.52,P<0.01)、小疮菊(F=2718.23,P<0.01)以及异形狭果鹤虱(F=18604.17,P<0.01)。从上述结果可看出,同种植物产生不同种子在形状上均存在极显著差异。
依据Thompson对种子形状分类的方法,对种子形状指数进行了计算,结果见表1。禾本科和菊科为长条或扁平型种子,而十字花科、豆科、紫草科及藜科则多为圆形种子。对种子形状指数进行单因素方差分析,在科分类水平上,各科间种子形状存在极显著差异(F=13.70,P<0.01)。其中禾本科和菊科两类种子形状指数平均值分别为0.15和0.13,十字花科、豆科、紫草科及藜科种子形状指数平均值分别为0.05、0.06、0.06和0.08。
含植物种类较多的科,其种间种子形状差异极显著,其中十字花科(F=144711.22,P<0.01),禾本科(F=126005.20,P<0.01),藜科(F=4786.25,P<0.01),菊科(F=3728.28,P<0.01),豆科(F=71417.33,P<0.01),紫草科(F=336970.15,P<0.01)。
对同种植物异型种子形状指数进行单因素方差分析,结果表明,异果芥(F=1366.72,P<0.01)、紫翅猪毛菜(F=25632.60,P<0.01)、角果藜(F=4608990.12,P<0.01)、野榆钱菠菜(F=565156.82,P<0.01)、异苞滨藜(F=170058.52,P<0.01)、异子蓬(F=1203.81,P<0.01)、小疮菊(F=223066.71,P<0.01)以及异形狭果鹤虱(F=307.37,P<0.01)。从上述结果可看出,同种植物产生不同种子在形状上均存在极显著差异。
2.3 基于系统进化树的AOT法和K值法的种子大小、形状变异
利用Phylomatic 3.0网页版进行各物种系统进化树的构建,形成9个进化水平(图2)。进一步利用Phylocom中的AOT模块对各节点所涉及物种进行性状分析,结果表明,各物种在系统发育树上共形成48个内部节点。在这48个内部节点中,种子大小明显保守的节点有8个,而过度进化分异的节点仅有1个。种子大小进化明显保守的8个节点中,有6个为科间或属间节点,有2个为属内节点。除了这9个节点之外,其余39个节点均表现出进化适度分异。
对种子形状指数的谱系分析结果表明,形状指数在48个内部节点中,进化明显保守的节点为9个,而过度进化分异的节点有12个。形状指数明显保守的9个节点中,有8个为科间或属间以上的分类节点,1个为属内节点。进化过度分异的12个节点中,11个为科间以上节点,属内节点仅有1个。除了这21个节点外,其余27个节点均表现出进化适度分异。

表1 种子大小和种子形状指数Table 1 The characters of seed mass and seed shape index

续表1 Continued Table 1

续表1 Continued Table 1
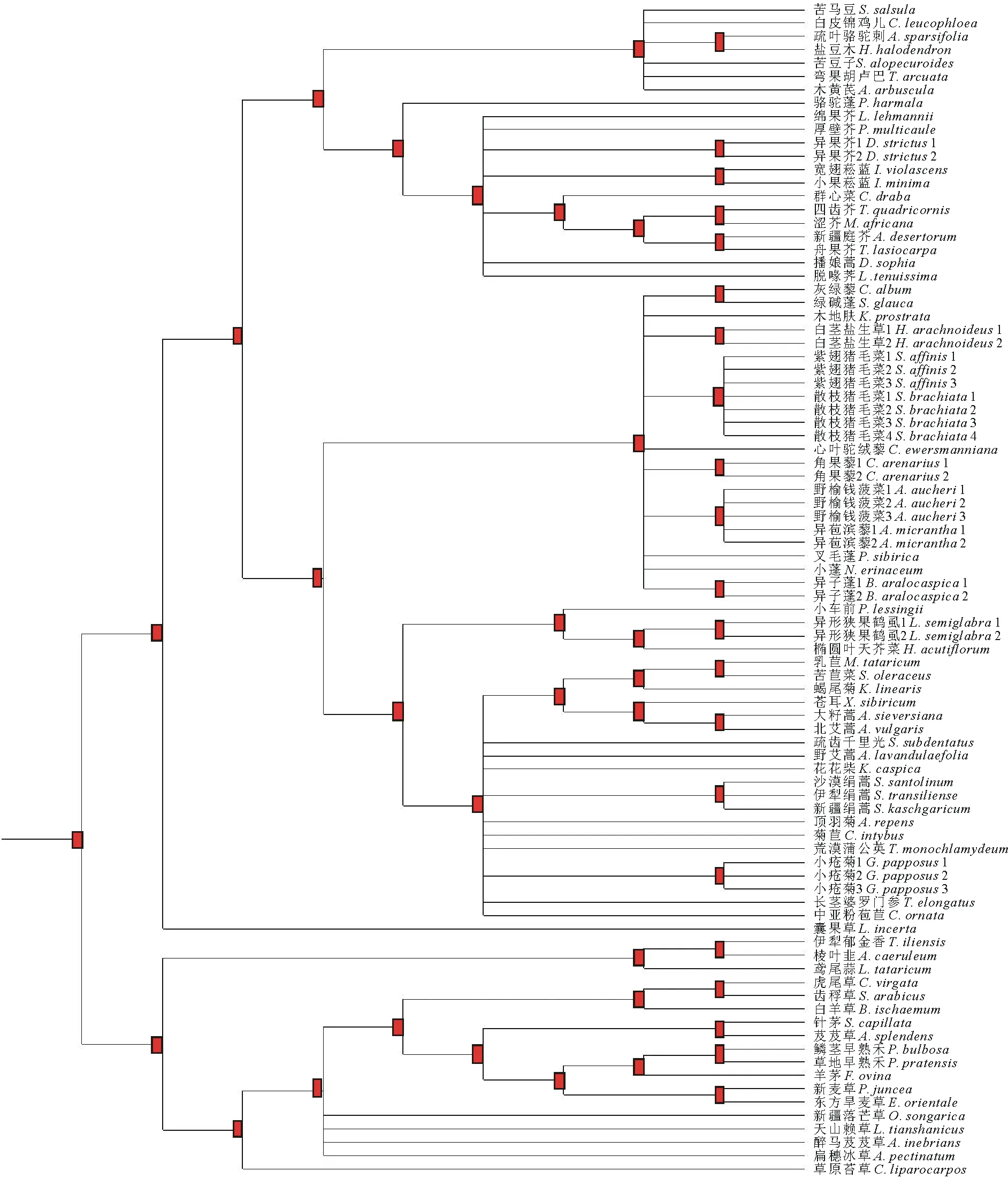
图2 系统进化树Fig.2 The phylogeny tree 利用Phylomatic 3.0网页版生成,小方块为内部节点位置。Created by Phylomatic 3.0, small rectangle is position of 48 internal nodes.
由于单纯从AOT法计算结果中无法准确判定种子性状与进化历史的整体相关性,因此,采用K值法进一步对其种子性状的谱系特征进行分析(表2)。K值法的计算结果表明,种子大小和种子形状的K值均为0.27,但种子大小并无显著的谱系信号(P>0.05),说明这一性状与物种进化历史无关。而种子形状K值小于1,并表现出极显著的谱系信号(P<0.01),充分说明亲缘关系越近的物种其性状越相似,其进化过程与系统进化历史密切相关。
2.4 种子性状对荒漠环境的生态适应类型及表现
所研究的88种植物均在荒漠或半荒漠生境中广泛分布,在长期的干旱和波动环境条件的塑造下,种子性状会产生对荒漠环境明显的进化适应。对88种种子的适应性特征进行总结,结果见表3。
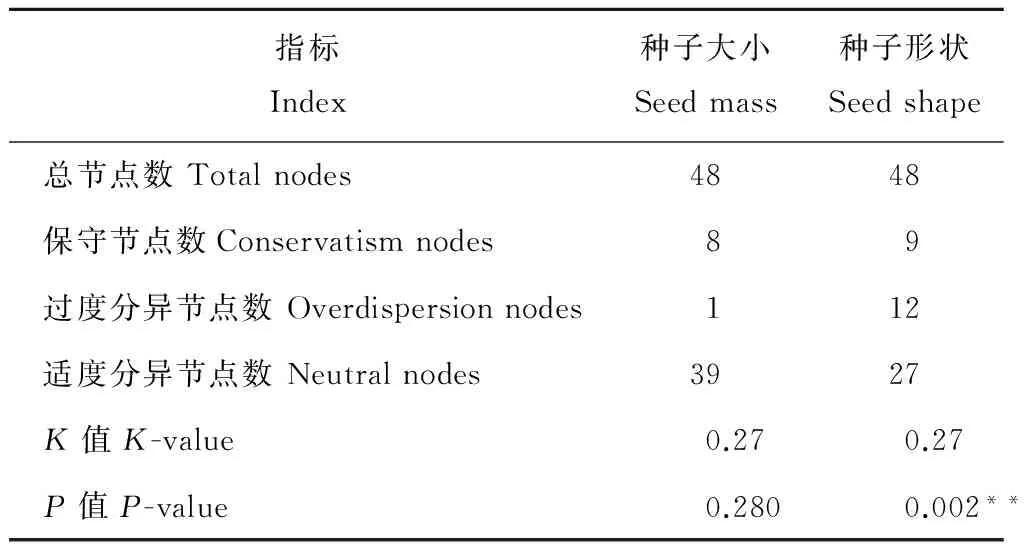
表2 基于AOT法和K值法的性状分析Table 2 Trait analysis with AOT and K-value means
注:**表示达到极显著水平(P<0.01)。
Note: ** indicates significance at the level of 0.01. 绢蒿荒漠植物在荒漠或半荒漠条件下主要产生了6种适应方式,包括种子多型性、产生粘液繁殖体、具助力传播结构、特殊生活型、家畜消化道传播以及结实方式适应。种子多型性是植物在极端环境条件下产生的极为特殊的策略,本研究所涉及的植物中,能够产生2种以上种子的有10种植物。种子表面分泌粘液是荒漠区植物另外一种重要的适应策略,本研究所涉及的植物中,能够产生该种适应方式的有11个物种,其中以十字花科和菊科蒿属植物为主。在荒漠区,植物结实后的种子远距离传播是开拓新生境的重要手段,可以很好地避免与母株或同胞产生竞争,因此植物本身也会进化出相应的助力传播结构,包括具芒、果翅、苞片、角果开裂弹射、冠毛或绒毛、钩刺、膜质果囊、种荚膜质并肿胀成膀胱状、种子圆形且流动性好等多样化的结构,为种子顺利传播到新生境创造了极为有利的条件,本研究所涉及的植物中有39种,是6种适应方式中物种数量最多的一类。短命和类短命植物是为充分利用荒漠区春季丰沛降水所产生的特殊适应类型, 这种生活型上的适应在新疆荒漠区也极为常见,其典型特征是在夏季来临前完成生活史,短命类植物种子成熟后死亡,而类短命则进入休眠状态,本研究所涉及的短命植物17种,类短命植物有4种。除了上述4种适应方式外,绢蒿荒漠植物还对家畜放牧过程产生了一定的适应,其中最明显的表现就是种子可通过家畜消化道进行传播,这类植物具有营养体或种子本身适口性好、营养价值高,种子和植物营养器官接触紧密,直接被家畜采食或通过采食营养器官而携带进入消化道,然后通过家畜的游走和粪便排泄实现种子的远距离传播。同时,在家畜消化道作用下,可打破某些植物(尤其是豆科植物)种子的休眠,提高了种子的萌发率,这对荒漠地区植物的新生境拓殖意义重大。
3 讨论
绢蒿荒漠是中亚荒漠的重要组成部分,由于受到中亚哈萨克斯坦气候型的影响,在此类生境中生活的植物已经产生了明显的适应性对策。从生活型上来讲,多以短命和类短命植物为主,并能以这两类生活型植物形成重要的植物层片[16]。同时,植物种子大小和形状也产生了极为鲜明的适应对策,一般而言,荒漠区种子往往采取产生大量小种子的r对策[17],本研究所涉及的物种中,种子大小在10.00 mg以下的占全部物种的93.24%,绝大多数为小型种子。
有关同一区系植物功能性状比较是众多学者关注的焦点问题,比较生物学和比较生态学在世界和我国已经有很长的历史,对于物种水平上的比较通常利用常规的统计方法[18],本研究中利用方差分析对种子大小和形状两个指标在科间、科内种间以及同一植物异型种子间分别进行了比较,结果显示不同分类水平上的种子大小和形状均存在极显著差异,前人的相关研究也得到了类似结论[17]。但是,在功能性状的比较过程中,常规的统计方法由于没有将物种之间的系统进化关系考虑进来,因而可能会影响到比较结果的精确性,而物种性状的系统比较方法可以圆满解决上述问题[18], 并且在近年来被众多学者接受认同[19]。本研究利用Phylomatic 3.0网页版对88种植物进行了系统进化树的绘制,反映出所研究物种的进化历史,然后用Phylocom中的AOT模块和Picante软件包中的K值法对性状特征值与进化历史之间的相关性进行了分析。AOT分析可得到各个内部节点的进化保守(非保守)特征,而K值法可判定所有物种整体上的进化谱系信号,二者结合可对不同节点和全部物种进化特征进行全面分析。种子大小进化的相对保守性在很多文献中都已经被明确指出[20],但在本研究中种子大小却并没有显著的谱系信号。其主要原因可能是在荒漠和半荒漠多变环境条件下,虽然不同植物所采取的适应策略存在差别,但在结实上都存在由于物种和环境条件不同繁殖分配上的两种权衡[21],一是种子大小和种子数量二者之间的权衡,二是种子本身和其附属结构之间的权衡,两种权衡造成的最终结果使种子大小产生更大的可变性[22]。但是,种子形状与种子大小在进化过程中完全不同,由于种子形状指数综合了种子长、宽、高的三维信息,即使种子大小会由于资源量波动而出现变化,其长、宽、高会由于种子大小而产生变化,但三者的变化具有同步性,最终用三者方差大小反映的种子形状指数不会发生显著变化,因此,使种子形状具有很强的稳定性。本研究中的种子形状具有显著的谱系信号,充分说明种子形状与系统进化的历史密切相关,这也是经典植物分类学中将植物种子形状特征作为分类依据的主要原因。

表 3 88种植物种子对荒漠环境的适应类型及表现Table 3 Adaptation types and expression of 88 plants or seeds to desert ecoenvironment
荒漠区植物种子的适应对策一直是众多学者关注的焦点,已经对同种植物种子多型性[23]、种子粘液[24]、助力传播的附属结构[25]、生活型[26]以及动物消化道传播[27]等方面进行了大量的研究工作。绢蒿荒漠特殊的气候和资源条件使植物产生了相应的适应策略,本研究所涉及的所有植物种子都会存在1~2种以上的适应对策,涉及种子结实类型、萌发以及传播方式等方面。
种子多型性是同种植物产生多种类型种子的一种生物学现象,不同种子在萌发行为上存在显著差别,多出现在藜科、十字花科及菊科植物中,该类植物通常通过产生多样化的种子而实现在多变环境条件中的“多头下注”,从而保证种群在多变环境中的成功繁衍[28]。粘液结构种子多出现在菊科、十字花科及车前科植物中[29],该结构可在短时间内吸收水分而形成保水层,同时可粘结土壤、沙粒使种子大粒化种子表面粘液层,为种子萌发创造极为有利的条件[30]。很多附属结构的存在为种子多渠道传播提供了保障,附属结构是种子在长期适应进化过程中形成的,是植物在适应环境过程中繁殖分配“权衡”的具体表现,由于其对种子传播过程意义重大,因此也被认为是种子的重要组成部分,同时也增加了种子在适应环境过程中的可塑性[31]。除了与助力传播有关的附属结构外,有些植物种子表面光滑或者呈圆形,增加了种子的流动性,也可帮助种子实现远距离传播[27],本研究中的豆科植物种子就属于此类。总之,绢蒿荒漠中大部分植物均会采取产生助力传播结构而实现种子的有效传播。对荒漠区波动的水分条件适应就产生了短命和类短命植物这两个特殊生活型,其简短的生活史和水分条件完美契合是这两类植物生生不息的关键,本研究中的十字花科、鸢尾科以及禾本科中的某些植物都属于这两类,这两类植物也在植物群落方面增加了植被层片结构的丰富度,同时也为春季放牧提供了大量的可食牧草。人们很早就关注到动物消化道传播对种子扩散过程的贡献,本研究中有24种植物满足消化道传播的条件,并且这种传播在放牧条件下是更为有效的种子扩散方式。绢蒿荒漠在牧业生产中通常作为春秋放牧地,秋季放牧时间和种子成熟时间重合,就为该传播方式提供了时间上的便利,这也是荒漠区植物对放牧过程的一种典型适应。
4 结论
88种植物中,绝大多数种子质量为0.10~10.00 mg,呈现明显的正态分布。88种植物分属6种生活型,其中一年生草本、多年生草本和短命植物占到88%。种子形状指数为0.00~0.20,细长或扁平种子有42种,球形或接近球形种子有46种。种子大小和形状指数在不同科间、科内种间以及同种植物异型种子间均存在极显著差异。
88种植物在系统发育树上共形成48个内部节点。种子大小明显保守的节点有8个,而过度进化分异的节点仅有1个,其余39个节点均表现出进化适度分异。形状指数明显保守的节点为9个,而过度进化分异的节点有12个,其余27个节点表现出进化适度分异。种子大小无明显的谱系信号,与系统进化历史无关,产生了与环境条件相适应的进化,因此,种子大小会由于环境条件波动表现出较强的可塑性。而种子形状具明显的谱系信号,与系统进化历史密切相关,因此,种子形状会由于三维变化的同步性表现出较强的稳定性。
88种植物种子绝大多数能够产生与荒漠、半荒漠气候以及放牧压力相适应的对策,主要有种子多型性、产生种子粘液层、具有助力传播结构、特殊生活型以及动物消化道传播等6种适应类型,很多植物在复杂环境条件下可产生多种适应类型,可为种子萌发、传播及种群繁衍创造良好条件。
参考文献References:
[1] Harper J L, Moore K G. The shapes and sizes of seeds. Annual Review of Ecology & Systematics, 1970, 1(1): 327-356.
[2] Thompson K, Hodgson J G. Seed size and shape predict persistence in soil. Functional Ecology, 1993, 7(2): 236-241.
[3] Westoby M, Jurado E, Leishman M. Comparative evolutionary ecology of seed size. Trends in Ecology & Evolution, 1992, 7(11): 368-372.
[4] Venable D L, Brown J S. The selective interactions of dispersal, dormancy, and seed size as adaptations for reducing risk in variable environments. American Naturalist, 2010, 131(3): 360-384.
[5] Holley J C, Moreau C S, Laird J G,etal. Subcaste-specific evolution of head size in the ant genusPheidole. Biological Journal of the Linnean Society, 2016, 118(3): 472-485.
[6] Zhong Y K, Bao Q H, Sun W,etal. The influence of mowing on seed amount and composition in soil seed bank of typical steppe Ⅲ size and weight of seeds of 120 plant species. Acta Scientiarum Naturalium Universitatis Neimongol (Natural Science Edition), 2001, 32(3): 280-286.
仲延凯, 包青海, 孙维, 等. 割草干扰对典型草原土壤种子库种子数量与组成的影响 Ⅲ 120种植物种子的大小与重量. 内蒙古大学学报(自然科学版), 2001, 32(3): 280-286.
[7] Qi W, Guo S Q, Cui X L,etal. Variation of seed mass and SLA of 63 plant species in different habitats at various altitudes on the eastern Qinghai-Tibet Plateau. Acta Prataculturae Sinica, 2012, 21(6): 42-50.
齐威, 郭淑青, 崔现亮, 等. 青藏高原东部4科植物种子大小和比叶面积随海拔和生境的变异. 草业学报, 2012, 21(6): 42-50.
[8] Liu Z M, Jiang D M, Yan Q L,etal. Study on dispersal biology of common species of flora of the Horqin Steppe. Acta Prataculturae Sinica, 2005, 14(6): 23-33.
刘志民, 蒋德明, 阎巧玲, 等. 科尔沁草原主要草地植物传播生物学简析. 草业学报, 2005, 14(6): 23-33.
[9] Wang L, Liu T, Han Z Q,etal. Variations and spatial distribution pattern of seed mass in the Gurbantunggut Desert. Acta Ecologica Sinica, 2014, 34(23): 6797-6806.
王琳, 刘彤, 韩志全, 等. 古尔班通古特沙漠植物种子大小变异的空间格局. 生态学报, 2014, 34(23): 6797-6806.
[10] Moles A T, Ackerly D D, Webb C O,etal. A brief history of seed size. Science, 2005, 307: 576-580.
[11] Zhang S T, Du G Z, Chen J K. The present situation and prospect of studies on evolutionary ecology of seed size variation. Acta Ecologica Sinica, 2003, 23(2): 353-364.
张世挺, 杜国祯, 陈家宽. 种子大小变异的进化生态学研究现状与展望. 生态学报, 2003, 23(2): 353-364.
[12] Xu P. Utilization of grassland resources in Xinjiang. Urumqi: Xinjiang Science and Technology Health, 1993.
许鹏. 新疆草地资源及其利用. 乌鲁木齐: 新疆科技卫生出版社, 1993.
[13] Yang J J, Wan J J, Wali Narikes,etal. Seed morphology and effect of sheep rumen digestion on germinability of 28 grass of Tianshan. Acta Prataculturae Sinica, 2015, 24(2): 104-115.
杨洁晶, 万娟娟, 娜丽克斯·外里, 等. 28种植物种子形态学性状及其萌发对绵羊瘤胃消化的反应. 草业学报, 2015, 24(2): 104-115.
[14] Jin D M. Leaf traits of woody species in east China forests and their relationships with climate. Beijing: University of Chinese Academy of Sciences, 2010.
金冬梅. 中国东部森林木本植物的叶性状及其与气候的关系. 北京: 中国科学院大学, 2010.
[15] Kembel S W, Cowan P D, Helmus M R,etal. Picante: R tools for integrating phylogenies and ecology. Bioinformatics, 2014, 26(11): 1463-1464.
[16] Lu W H. Study on regeneration characteristics of degradedSeriphidiumdesert grassland under short term fencing. Urumqi: Xinjiang Agriculture University, 2010.
鲁为华. 短期围栏封育下退化绢蒿荒漠草地更新特征研究. 乌鲁木齐: 新疆农业大学, 2010.
[17] Wang L. Seed mass variation and its adaptive significances in Gurbantunggut Desert, China. Shihezi:Shihezi University,2013.
王琳. 古尔班通古特沙漠种子大小变异的适应意义. 石河子: 石河子大学, 2013.
[18] Jin D M, Li Y, Sun S C. Principles and applications of phylogenetic comparative methods: an overview. Acta Ecologica Sinica, 2008, 28(3): 1247-1256.
金冬梅, 李妍, 孙书存. 系统比较方法的原理及应用. 生态学报, 2008, 28(3): 1247-1256.
[19] Cao K, Rao M D, Yu J Z,etal. The phylogenetic signal of functional traits and their effects on community structure in an evergreen broad-leaved forest. Biodiversity Science, 2013, 21(5): 564-571.
曹科, 饶米德, 余建中, 等. 古田山木本植物功能性状的系统发育信号及其对群落结构的影响. 生物多样性, 2013, 21(5): 564-571.
[20] Qi W. The distribution, variation and evolution of seed size on the eastern Qinghai-Tibet plateau. Lanzhou: Lanzhou University, 2010.
齐威. 青藏高原东缘种子大小的分布、变异和进化规律研究. 兰州: 兰州大学, 2010.
[21] Harper J L, Clatworthy J N. The evolution and ecology of closely related species living in the same area. Evolution, 1961, 15(2): 209-227.
[22] Ren J Y. The study of variation in seed and seedling adaptive strategies ofBetulaandQuercuswith elevation in Qinling Mountains. Xi’an: Northwest University, 2014.
任坚毅. 秦岭桦属栎属植物种子与幼苗适应性策略的海拔变异. 西安: 西北大学, 2014.
[23] Lu J, Tan D, Baskin J M,etal. Fruit and seed heteromorphism in the cold desert annual ephemeralDiptychocarpusstrictus(Brassicaceae) and possible adaptive significance. Annals of Botany, 2010, 105(6): 999-1014.
[24] Song M F, Liu H L, Zhang D Y,etal. Screening and comparison on myxospermy of 120 species occurring on the Junggar desert. Journal of Desert Research, 2009, 29(6): 1134-1140.
宋明方, 刘会良, 张道远, 等. 准噶尔荒漠120种植物粘液繁殖体的甄别与比较. 中国沙漠, 2009, 29(6): 1134-1140.
[25] Bai C K, Cao B, Li G S. Correlations of plant seed dispersal pattern with genome size and 1000-seed mass. Chinese Journal of Ecology, 2013, 32(4): 832-837.
白成科, 曹博, 李桂双. 植物种子传播途径与基因组值和千粒重的相关性. 生态学杂志, 2013, 32(4): 832-837.
[26] Liu Z M, Li X H, Li R P,etal. A comparative study on diaspore shape of 70 species found in the sandy land of Horqin. Acta Prataculturae Sinica, 2003, 12(5): 55-61.
刘志民, 李雪华, 李荣平, 等. 科尔沁沙地70种植物繁殖体形状比较研究. 草业学报, 2003, 12(5): 55-61.
[27] Yang J J, Wali Narikes, Lü Y P,etal. Effect of herbivore’s gut passage on seed germination of legume: A Meta analysis. Chinese Journal of Ecology, 2015, 34(10): 2833-2842.
杨洁晶, 娜丽克斯·外里, 吕艳萍, 等. 草食动物对豆科植物种子萌发的消化道作用效应的Meta分析. 生态学杂志, 2015, 34(10): 2833-2842.
[28] Wei Y. Seed polymorphism and germination behavior of several species from Chenopodiaceae inhabiting the Junggar Basin. Urumqi: Xinjiang Agriculture University, 2007.
魏岩. 准噶尔荒漠几种藜科植物的种子多型性及其萌发行为研究. 乌鲁木齐: 新疆农业大学, 2007.
[29] Ma J L, Liu Z M. Myxospermy and its ecological function. Chinese Journal of Ecology, 2006, 25(11): 1400-1404.
马君玲, 刘志民. 粘液繁殖体及其生态功能. 生态学杂志, 2006, 25(11): 1400-1404.
[30] Luo H, Yan X H, Zhou B,etal. Water absorbance features of seed mucilage of an invasive plant,Plantagovirginica, and its germination response to drought stress. Chinese Journal of Ecology, 2015, 34(8): 2155-2160.
罗辉, 闫小红, 周兵, 等. 入侵植物北美车前种子粘液的吸水特性及其对干旱胁迫的萌发响应. 生态学杂志, 2015, 34(8): 2155-2160.
[31] Yu S L, Fang W W. New advances in seed geography. Chinese Journal of Plant Ecology, 2012, 36(8): 918-922.
于顺利, 方伟伟. 种子地理学研究的新进展. 植物生态学报, 2012, 36(8): 918-922.
猜你喜欢
中学生天地(A版)(2022年11期)2022-11-25
小哥白尼(趣味科学)(2022年5期)2022-08-15
绿色中国(2019年14期)2019-11-26
新世纪智能(英语备考)(2018年11期)2018-12-29
小学生学习指导(低年级)(2018年9期)2018-09-26
读者·校园版(2018年1期)2017-12-16
儿童故事画报·智力大王(2016年7期)2017-02-08
小天使·五年级语数英综合(2016年12期)2016-12-09
文史春秋(2016年3期)2016-12-01
浙江大学学报(农业与生命科学版)(2015年4期)2015-12-15
