
铅胁迫对金丝草生长及生理生化的影响
2018-04-18 07:48韩航陈顺钰薛凌云侯晓龙蔡丽平刘爱琴周垂帆
草业学报 2018年4期
韩航,陈顺钰,薛凌云,侯晓龙, 2, 3*,蔡丽平,3,刘爱琴,3,周垂帆,3
(1.福建农林大学林学院, 福建 福州350002;2.福建农林大学资源与环境学院,福建 福州350002;3.海峡两岸红壤区水土保持协同创新中心,福建 福州350002)
近年来,由于我国矿产资源大规模开采,富含重金属污染物无序排放,农药化肥大量使用等因素,导致土壤重金属污染严重。据环境保护部和国土资源部联合发布全国土壤污染状况调查显示,全国土壤总的点位超标率达16.1%,部分工矿业地区土壤重金属超标范围较大,土壤环境质量堪忧[1]。其中,铅污染因其毒害作用强、影响范围广等而备受关注,成为当前急需解决的环境问题[2]。
土壤重金属污染治理传统的工程、化学等方法存在需扰动土壤、费用昂贵等缺点,难以大规模推广应用。植物修复技术因其绿色环保、经济、操作性强等特点,在土壤重金属污染治理方面有广阔应用前景[3-4],植物修复技术的应用前提是筛选出重金属超富集植物。多年生禾本科植物金丝草(Pogonatherumcrinitum),又名落苏、黄毛草、毛毛草,是课题组在福建尤溪铅锌矿区发现,并通过室内试验验证的一种Pb的超富集植物,可在Pb浓度高达17000 mg·kg-1的铅锌矿区正常生长,对Pb具有较强耐性[5]。金丝草的铅耐性是其长期适应Pb胁迫环境的演化结果,其中可能存在特殊的适应机制。大量研究表明,抗氧化系统及渗透调节物质的调节响应很大程度影响着植物的拮抗能力[6]。黑麦草可通过提高SOD、POD活性和可溶性糖及脯氨酸含量,增强体内清除活性氧和渗透调节能力,降低铅毒害[7]。旱柳在Pb处理(<600 mg·kg-1)下,则通过根长和生物量调节,同时诱导POD、CAT和SOD等酶活响应来适应土壤铅逆境[8]。但目前对金丝草适应土壤铅胁迫的机理尚不清楚,特别是抗氧化系统和渗透调节系统在适应过程中发挥的作用仍有待验证。
鉴于此,以金丝草为研究对象,测定不同梯度铅胁迫下金丝草形态、生物量、抗氧化系统、渗透调节物质等指标,比较金丝草叶片和根系响应差异,探讨逆境下金丝草抗氧化系统及渗透调节物质的响应规律,以期为耐性植物逆境响应机理研究提供科学参考。
1 材料与方法
1.1 试验材料
试验于2016年5月展开,供试金丝草苗来自福建农林大学森环所,选取10 cm高、长势一致的新生幼苗待用。基质采用洗净河沙和黄心土,风干后过2 mm筛,按1∶3混合(参考砂壤土质地)。基本理化性质:pH 5.2,全碳、全氮、全磷、全钾含量分别为:3.37、0.46、0.17和37.62 g·kg-1,速效磷、速效钾含量分别为:0.44、38.19 mg·kg-1,重金属Pb、Zn、Cu含量分别为:1.03、2.14和0.35 mg·kg-1。
1.2 试验设计
试验用盆直径30 cm,高25 cm,基质填充8 kg。参考国家土壤质量标准(GB 15618-1995),设计4个Pb浓度,0、1000、2000和3000 mg·kg-1(文中分别以CK、Pb 1000、Pb 2000、Pb 3000表示)。采用(CH3COO)2Pb溶液进行施加,并钝化30 d。移植金丝草前,一次性施加底肥,每kg土肥料用量N为100 mg、P2O2为80 mg,K2O为100 mg,施加形态为KH2PO4和尿素[9]。每组处理5个重复,每盆10株金丝草幼苗。试验在玻璃温室中进行,温度(25±2) ℃,湿度65%~75%。试验过程中,使用土壤温湿度仪监测土壤水分情况,及时补充土壤水分,保持含水量在田间持水量的70%左右。胁迫试验60 d时,取样测定金丝草叶片和根系相关形态和生理指标。
1.3 测定方法
株高、叶片指标和生物量测定:在胁迫60 d收获时,每盆随机选取5株金丝草使用直尺测量株高,以金丝草最长叶尖计,计算均值后为一次重复,每个处理组共测定5盆;叶长、叶宽和叶面积采用北京Yaxin-1241叶面积仪测定,每盆随机选取5株金丝草,选取金丝草自上至下第3~9片顺序中任一叶片,共5片,其中叶长以叶身计算,叶宽以叶身中部最宽处计算,计算均值后为一次重复,每个处理组共测定5盆;收获金丝草洗净后,将地上部和根系分开,分别测定鲜重。然后105 ℃杀青30 min,75 ℃烘干至恒重后分别记录地上和根系部分干重,并计算根冠比。
抗氧化酶与MDA含量、抗氧化能力、渗透调节物质测定:每盆金丝草收获后,分别取叶片和根系,洗净混匀后各称取0.2 g金丝草叶片和根系,加入5 mL预冷的pH 7.0的磷酸缓冲液,研磨成匀浆,于4 ℃下10000 r·min-1冷冻离心20 min,取上清液放4 ℃冰箱冷藏待测。超氧化物歧化酶(SOD)、过氧化物酶(POD)、过氧化氢酶(CAT)和丙二醛(MDA)含量分别采用氮蓝四唑法、愈创木酚法、紫外吸收法测定和硫代巴比妥酸法测定[10]。抗超氧阴离子自由基活力(anti superoxide anion radical activity, ASAFR)和总抗氧化能力(total antioxidant capacity, T-AOC)均使用南京建成生物工程研究所提供的试剂盒(准确称取组织重量,按重量体积比加入9倍的PBS制成10%的组织匀浆,2500 r·min-1,离心10 min,分别按试剂盒步骤操作加入各试剂后,1 cm光径,波长550和520 nm处分别测定吸光度得ASAFR、T-AOC);可溶性蛋白(soluble protein, SP)含量采用考马斯亮蓝法测定[11],可溶性糖(soluble sugar, SS)含量采用蒽酮比色法测定[11],脯氨酸(proline, Pro)含量采用茚三酮比色法测定[12]。
1.4 数据统计分析
数据均为5次重复试验结果,使用SPSS 20.0进行处理,采用单因素方差分析(One-way ANOVA)对试验数据进行分析比较。采用Origin 8.5软件制图。
2 结果与分析
2.1 铅胁迫对金丝草生长的影响
由图1可知,Pb 1000处理下,金丝草株高、叶长和叶宽较对照无显著变化。随胁迫浓度增加,金丝草株高、叶长、叶宽均低于对照,抑制效果显著(P<0.05)。其中,Pb 2000和Pb 3000处理下金丝草株高分别降低了13.4%和26.8%,叶长分别降低22.6%和26.9%,叶宽分别降低23.3%和18.4%。此外,铅处理下金丝草叶面积均显著低于对照(P<0.05)。

图1 不同浓度铅胁迫对金丝草地上部生长的影响Fig.1 The growth of aboveground parts of P. crinitum under Pb stress 不同小写字母表示差异显著(P<0.05),下同。The different letters mean the significant differences at P<0.05, the same below.
2.2 铅胁迫对金丝草生物量的影响
由表1可知,低浓度铅胁迫下(Pb 1000),金丝草地上和地下部分鲜重、干重及全株总质量均高于对照,其中地上部分干重增长13.4%,根干重增长14.7%。随胁迫浓度增加,金丝草生物量逐渐受到抑制。Pb 3000处理下,金丝草地上和地下部鲜重、干重及全株质量均显著低于对照(P<0.05),全株质量下降约62.8%。且铅处理下金丝草根冠比均大于对照,金丝草根系生物量占比逐渐增加。综上,金丝草生物量指标在铅胁迫下总体呈“低促高抑”趋势。

表1 不同浓度铅胁迫对金丝草生物量的影响Table 1 The biomass of P. crinitum under Pb stress
注:同列不同小写字母表示差异显著(P<0.05),下同。
Note:The different letters within the same column mean the significant differences atP<0.05, the same below.
2.3 铅胁迫对金丝草抗氧化酶活性的影响
由图2(A)可知,铅胁迫对金丝草叶片SOD活性抑制作用显著(P<0.05),但胁迫浓度的增加对叶片SOD活性影响较小;金丝草根系SOD活性呈先增后减趋势,Pb 2000处理下达最大值。且对照和Pb处理下金丝草叶片SOD活性均高于根系。由图2(B)可知,铅处理对金丝草叶片POD活性有一定促进作用,Pb 1000和Pb 2000处理下,叶片POD活性分别比对照增长17.9%和19.0%,但与对照无显著差异(P>0.05);不同铅胁迫处理金丝草根系POD活性均小于对照。同时,铅胁迫条件下金丝草叶片POD活性均大于根系,而对照则相反。由图2(C)可知,随铅胁迫浓度的增加,金丝草叶片和根系CAT活性均呈先减后增趋势,Pb 1000和Pb 2000处理下,叶片和根系CAT活性低于对照,Pb 3000处理下较对照无显著差异。此外,对照及Pb 1000处理下金丝草叶片CAT活性大于根系,而Pb 2000 和Pb 3000则相反。
2.4 铅胁迫对金丝草抗氧化能力和MDA含量的影响
由图3(A)可知,铅胁迫下金丝草叶片抗超氧阴离子自由基活力(ASAFR)显著低于对照(P<0.05),不同铅处理间无显著差异(P>0.05);金丝草根系在Pb 1000处理时高于对照,在Pb 3000处理时,显著低于对照(P<0.05),呈先增后减趋势。且除Pb 3000处理外,金丝草根系抗超氧阴离子自由基活力高于叶片。
植物总抗氧化能力(T-AOC)是植物体内酶促、非酶促等抗氧化物质总的抗氧化能力,受诸多因素影响,逆境下常表现为非线性、复杂性的规律[13]。由图3(B)可知,土壤铅胁迫对金丝草叶片总抗氧化能力(T-AOC)有一定促进作用,Pb 3000处理时比对照增长约76.3%,显著高于对照及其他铅处理(P<0.05);金丝草根系T-AOC在Pb 3000处理下显著降低(P<0.05),Pb 1000和Pb 2000与对照无显著差异(P>0.05);不同Pb处理和对照金丝草叶片T-AOC均大于根系。故推测,金丝草在Pb 1000~2000处理下,叶片和根系总抗氧化能力处于稳态,在Pb 3000处理下,由于叶片和根系抗氧化系统响应水平不同,导致根系T-AOC显著下降,叶片则显著增加。
由图3(C)可知,铅处理下叶片MDA含量呈增长趋势,Pb 2000和Pb 3000处理时,显著增加(P<0.05);根系MDA含量在铅胁迫下均高于对照,其中Pb 1000处理显著大于对照(P<0.05),而Pb 2000和Pb 3000与对照无显著差异(P>0.05);对照及Pb处理下金丝草叶片MDA含量均高于根系。
2.5 铅胁迫对金丝草渗透调节物质的影响
由图4(A)可知,铅处理下金丝草叶片可溶性蛋白含量均高于对照,Pb 2000处理下达最大值,较对照增加36.4%,显著大于对照(P<0.05);金丝草根系可溶性蛋白含量则先减后增,在Pb 2000和Pb 3000处理高于对照;金丝草叶片可溶性蛋白含量在铅处理及对照中均高于根系。由图4(B)可知,金丝草叶片可溶性糖含量在铅处理下呈先减后增趋势,在Pb 3000处理时达最大值,显著高于对照(P<0.05);Pb 1000和Pb 2000处理下金丝草根系可溶性糖含量分别比对照增加17.7%和19.0%,显著增长(P<0.05),在Pb 3000处理下受明显抑制。且叶片可溶性糖含量总体高于根系。由图4(C)可知,金丝草叶片脯氨酸含量总体高于根系。铅胁迫下金丝草叶片脯氨酸含量呈先减后增趋势,在Pb 3000处理下达最大值,与对照无显著差异(P<0.05);金丝草根系脯氨酸含量在Pb 1000处理时达最大值,随铅胁迫浓度增加,显著降低(P<0.05)。

图3 不同浓度铅胁迫对金丝草抗氧化能力的影响Fig.3 Effect of Pb stress on the antioxidant capacity of P. crinitum
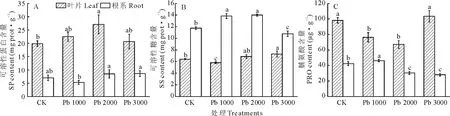
图4 不同浓度铅胁迫对金丝草渗透调节物质的影响Fig.4 Effect of Pb stress on the osmotic adjustment substances of P. crinitum
3 讨论
植物生长形态和生物量变化是植物适应环境生长的最终结果。沈高峰[14]发现,铅胁迫(≤1500 mg·kg-1)18 d时,高羊茅(Festucaarundinacea)和多年生黑麦草(Loliumperenne)鲜重分别为对照的67.44%和68.09%。李西等[15]也发现,土壤铅胁迫会抑制假俭草(Eremochloaophiuroides)、狗牙根(Cynodondactylon)与结缕草(Zoysiajaponica)地上和根系生物量。本研究中,除Pb 1000处理外,其他铅处理下金丝草株高、叶长、叶宽均显著低于对照,抑制作用明显,与前人研究基本一致。同时,金丝草地上与地下部分鲜重、干重及全株生物量在Pb 1000处理时达最大值。在Pb 2000处理时,与对照无显著差异,在Pb 3000处理时,显著降低,符合“低促高抑”规律。但铅处理下金丝草根冠比逐渐增大,这可能是金丝草增强根系生物量占比,提高逆境下对养分、水分的汲取能力来主动适应铅胁迫的结果。
重金属铅胁迫会抑制植物生长、导致植株矮化、叶片面积变小,降低植物鲜重和干重。同时,植物在铅胁迫下体内会产生多种活性氧(ROS),造成膜脂质过氧化毒害、抑制酶活、削弱细胞正常渗透调节能力等[16]。为了避免和降低重金属毒害作用,植物有一套复杂的防御机制。其中诱导抗氧化酶系统响应和提高渗透调节物质水平来清除植株体内活性氧自由基、维持细胞正常渗透压和水势,是耐性植物逆境生存的重要手段[17]。SOD、POD、CAT是植物抗氧化系统重要组成部分,具有清除O2-·、H2O2的作用,逆境下不同植物抗氧化酶响应敏感程度和响应规律略有不同[18]。有研究发现,香根草苗在水培Pb胁迫下,随Pb2+浓度的增加(0~8 mmol·L-1),香根草苗SOD活性显著增加,POD和CAT活性则在高浓度(>4 mmol·L-1)Pb处理时,逐渐受到抑制[19]。灰绿藜和小藜幼苗SOD及POD活性则分别在200、100 mg·kg-1铅浓度下达到最高,CAT的活性则在100 mg·kg-1铅浓度时达到最大[20]。本研究中,除金丝草叶片SOD活性和根系POD活性受Pb胁迫明显抑制外,金丝草叶片和根系其他酶活并未表现出单一增减规律。如金丝草根系SOD活性无显著变化,叶片POD活性趋势为先增后减。这可能是在一定阈值内铅胁迫对金丝草抗氧化酶活性有刺激作用,但随胁迫浓度增加,金丝草酶活系统受损,酶活降低,与李铮铮等[21]结论类似。且同一浓度铅处理下,金丝草叶片与根系酶活响应水平不同,Pb 1000~3000处理下金丝草叶片SOD、POD活性均高于根系,推测是金丝草根系与污染土壤直接接触,根系生长受阻、根系过氧化作用对酶活抑制更强造成的。综上,金丝草主要通过叶片和根系抗氧化酶差异化响应,提高叶片和根系CAT活性,维持根系SOD活性、叶片POD活性稳态,协同配合来降低过氧化毒害。
另一方面,抗超氧阴离子自由基活力、总抗氧化能力和MDA含量高低反映了Pb胁迫下植物细胞膜脂过氧化程度,也体现出植物体内各类抗氧化物质总的抗氧化能力[13]。本研究中,随铅处理浓度增加,金丝草叶片和根系抗超氧阴离子自由基活力显著低于对照(除Pb 1000处理下金丝草根系外),金丝草叶片和根系MDA含量均高于对照,铅胁迫毒害渐显。但金丝草叶片和根系T-AOC在Pb 1000~2000处理下处于稳态,叶片T-AOC在Pb 3000处理下显著增长。这表明铅胁迫在导致金丝草叶片和根系膜脂过氧化损伤的同时,也会诱导金丝草体内抗氧化酶、抗氧化物等抵御机制响应,使得金丝草叶片和根系总抗氧化能力处于高位水平,增强了金丝草整体拮抗能力。
此外,可溶性蛋白、可溶性糖和脯氨酸是植物体内重要的渗透调节物质,具有保持逆境下植物体正常含水量和膨压的功能,是植物抗氧化系统中非酶物质重要的组成部分[22-23]。土壤铅胁迫可能诱导了金丝草细胞内部基因表达等复杂机制的响应,使得铅处理下叶片和根系的可溶性蛋白、可溶性糖含量处于较高水平,间接促进了金丝草体内活性氧清除,平衡了细胞质与液泡间渗透势,使得金丝草体内细胞代谢等生理活动可正常运作,这可能是金丝草主要的抗逆机制之一。
4 结论
金丝草对土壤铅胁迫有较强耐性,金丝草通过提高叶片POD活性、根系SOD活性等不同器官抗氧化酶差异化响应,促进金丝草叶片和根系可溶性蛋白和可溶性糖含量增加,维持金丝草体内总抗氧化能力处于较高水平,增强了金丝草抵御过氧化毒害和细胞逆境渗透调节能力,较好地适应了土壤铅胁迫。但本研究同时发现,在Pb 3000胁迫下金丝草POD和可溶性蛋白等指标降低,MDA含量有增加趋势,说明不同抗氧化酶响应Pb胁迫的作用有一定阈值,金丝草适应高浓度Pb胁迫可能还存在除抗氧化系统外的其他机理。
参考文献References:
[1] Wang Y J, Liu C, Zhou D M,etal. A critical view on the status quo of the farmland soil environmental quality in China: discussion and suggestion of relevant issues on Report on the national general survey of soil contamination. Journal of Agro-Environment Science, 2014, 33(8): 1465-1473.
王玉军, 刘存, 周东美, 等. 客观地看待我国耕地土壤环境质量的现状——关于《全国土壤污染状况调查公报》中有关问题的讨论和建议. 农业环境科学学报, 2014, 33(8): 1465-1473.
[2] Nemati K, Abu Bakar N K, Abas M R,etal. Speciation of heavy metals by modified BCR sequential extraction procedure in different depths of sediments from Sungai Buloh, Selangor, Malaysia. Journal of Hazardous Materials, 2011, 192(1): 402-410.
[3] Sánchez-Chardi A. Biomonitoring potential of five sympatric Tillandsia, species for evaluating urban metal pollution (Cd, Hg and Pb). Atmospheric Environment, 2016, 131: 352-359.
[4] Hou X L, Cai L P, Han H,etal. Effect of lead stress on the chorophyll fluorescence characteristics and antioxidative enzyme activities ofPaspalumnotatum. Acta Prataculturae Sinica, 2017, 26(3): 142-148.
侯晓龙, 蔡丽平, 韩航, 等. 铅胁迫对百喜草叶绿素荧光特性及酶活性的影响. 草业学报, 2017, 26(3): 142-148.
[5] Hou X L, Chang Q S, Liu G F,etal. Two lead-hyperaccumulator:PogonatherumcrinitumandLsacheglobosa. Chinese Journal of Environmental Engineering, 2012, 6(3): 989-994.
侯晓龙, 常青山, 刘国锋, 等. Pb超富集植物金丝草 (Pogonatherumcrinitum), 柳叶箬(Lsacheglobosa). 环境工程学报, 2012, 6(3): 989-994.
[6] Mahar A, Wang P, Ali A,etal. Challenges and opportunities in the phytoremediation of heavy metals contaminated soils: A review. Ecotoxicology & Environmental Safety, 2016, 126: 111-121.
[7] Liu H Q, Han J C, Liu H P,etal. Influence of lead gradient stress on the physiological and biochemical characteristics of perennial ryegrass (Loliumperenne) seedlings. Acta Prataculturae Sinica, 2012, 21(6): 57-63.
刘慧芹, 韩巨才, 刘慧平, 等. 铅梯度胁迫对多年生黑麦草幼苗生理生化特性影响. 草业学报, 2012, 21(6): 57-63.
[8] Zhu J, Wang P, Jia S S,etal. Tolerance, accumulation, translocation and stress response ofSalixmatsudanaKoidz to lead. Acta Scientiae Circumstantiae, 2016, 36(10): 3876-3886.
朱健, 王平, 夹书珊, 等. 旱柳(SalixmatsudanaKoidz)对Pb的耐性、富集、转运与胁迫响应研究. 环境科学学报, 2016, 36(10): 3876-3886.
[9] Sun J Y. Varietal difference in Cd tolerance and mechanism in cabbage. Nanjing: Nanjing Agricultural University, 2011.
孙建云. 甘蓝(BrassicaoleraceaL.)耐镉性的品种差异及其机理研究. 南京: 南京农业大学, 2011.
[10] Zou Q. Experimental guidance of plant physiology and biochemistry. Beijing: China Agriculture Press, 1995.
邹琦. 植物生理生化实验指导. 北京: 中国农业出版社, 1995.
[11] Li H S. Plant physiological and biochemical experimental principles and techniques. Beijing: Higher Education Press, 2000.
李合生. 植物生理生化实验原理和技术. 北京: 高等教育出版社, 2000.
[12] Wang X K, Huang J L. Principles and techniques of plant physiology and biochemistry experiment. Beijing: Higher Education Press, 2015.
王学奎, 黄见良. 植物生理生化实验原理与技术. 北京: 高等教育出版社, 2015.
[13] Gülçin. Antioxidant activity of food constituents: an overview. Archives of Toxicology, 2012, 86(3): 345-391.
[14] Shen G F. Effects of simulated acid rain and Pb combined stress on growth and antioxidative physiology of two kinds of turf grass. Journal of Anhui Agricultural Sciences. 2012, 40(22): 11283-11286.
沈高峰. 模拟酸雨和Pb复合胁迫对2种草坪草生长及抗氧化生理的影响. 安徽农业科学, 2012, 40(22): 11283-11286.
[15] Li X, Wu Y J, Sun L X,etal. Growth and physiological responses of three warm-season turfgrasses to lead stress. Acta Prataculturae Sinica, 2014, 23(4): 171-180.
李西, 吴亚娇, 孙凌霞. 铅胁迫对三种暖季型草坪草生长和生理特性的影响. 草业学报, 2014, 23(4): 171-180.
[16] Bankaji I, Caçador I, Sleimi N. Physiological and biochemical responses ofSuaedafruticosato cadmium and copper stresses: growth, nutrient uptake, antioxidant enzymes, phytochelatin, and glutathione levels. Environmental Science and Pollution Research, 2015, 22(17): 13058-13069.
[17] Tian B H, Zhang Y J, Zhang L P,etal. Effects of cadmium or chromium on growth and NADPH oxidase and antioxidant enzyme system of foxtail millet seedlings. Journal of Agro-Environment Science, 2016, 35(2): 240-246.
田保华, 张彦洁, 张丽萍, 等. 镉/铬胁迫对谷子幼苗生长和NADPH氧化酶及抗氧化酶体系的影响. 农业环境科学学报, 2016, 35(2): 240-246.
[18] Chen J W. Study on response and potential phytoremediation ofBidenspilosaL. in cadmium and lead stress. Chongqing: Southwest University, 2013.
谌金吾. 三叶鬼针草(BidenspilosaL.)对重金属Cd、Pb胁迫的响应与修复潜能研究. 重庆: 西南大学, 2013.
[19] Han L, Zhang X P, Liu B R,etal. Response ofVetiveriazizanioidesto Pb2+stress. Chinese Journal of Applied Ecology, 2005, 16(11): 2178-2181.
韩露, 张小平, 刘必融. 香根草对重金属铅离子的胁迫反应研究. 应用生态学报, 2005, 16(11): 2178-2181.
[20] Liu J H, Yao Z G. Effects of Pb stress on seed germination and seedling growth of two weed species seeds. Agricultural Science & Technology, 2016, 17(12): 2726-2731.
刘俊华, 姚志刚. 重金属铅胁迫对2种杂草种子萌发和幼苗生长的影响. 农业科学与技术 (英文版), 2016, 17(12): 2726-2731.
[21] Li Z Z, Wu J, Tang Y,etal. Effect of Pb, Zn and their interactions on the chlorophyll content and antioxidant enzyme systems ofHouttuyniacordatathumb. Acta Ecologica Sinica, 2007, 27(12): 5441-5445.
李铮铮, 伍钧, 唐亚, 等. 铅、锌及其交互作用对鱼腥草(Houttuyniacordata)叶绿素含量及抗氧化酶系统的影响. 生态学报, 2007, 27(12): 5441-5446.
[22] Marcu D, Damian G, Cosma C,etal. Gamma radiation impact on lettuce (Lactucasativavar.Capitata) growth and development, mitotic cell division and electron spin resonance analysis of induced free radicals. Environmental Engineering & Management Journal, 2014, DOI:10.1007/s10867-013-9322-z.
[23] Zhang J Y. The effect of Pb and Cd on physiological and biochemical indexes ofMonsteradeliciosaLiebm. in the short-term conditions. Journal of Soil and Water Conservation, 2016, 30(2): 340-345.
张家洋. 重金属铅镉短期胁迫对蓬莱蕉生理生化指标的影响. 水土保持学报, 2016, 30(2): 340-345.
猜你喜欢
艺术家(2021年4期)2021-05-19
哈哈画报(2018年4期)2018-09-10
中成药(2018年8期)2018-08-29
中成药(2018年2期)2018-05-09
小火炬·智漫悦读(2017年7期)2017-09-29
足球周刊(2016年15期)2016-11-02
华人时刊(2016年13期)2016-04-05
金色年华(2016年8期)2016-02-28
医学研究杂志(2015年9期)2015-07-01
中国当代医药(2015年9期)2015-03-01
