
川西北高寒沙地不同年限高山柳林下优势植物碳、氮、磷生态化学计量特征
2018-04-18 07:51:08佘淑凤胡玉福舒向阳严星李智王琴何佳贾安都
草业学报 2018年4期
佘淑凤,胡玉福,舒向阳,严星,李智,王琴,何佳,贾安都
(四川农业大学资源学院,四川 成都611130)
生态化学计量学是一门研究生态系统和生态过程中多重化学元素平衡关系的科学,它主要强调活有机体组成元素(特别是C、N和P)的化学计量特征关系[1]。碳(C)是植物体内干物质最主要的构成元素,也是各种生理生化过程的能量来源[2]。氮(N)、磷(P)分别作为蛋白质和核酸的重要组成元素,同时也是植物生长所需基本营养元素。C∶N和C∶P在一定程度上可以反映植物生长速度,并且能够表明植物生长速度与植物N和P的利用效率的相关性[3]。植物叶片N∶P的临界值被认为是判断土壤对植物养分供应状况的指标,因此植物N、P化学计量研究成为揭示植物养分限制状况及其适应策略的手段[4]。研究碳氮磷生态化学计量特征有助于认识植物应对不同生境条件的养分利用策略,对于解释植物与生境间的耦合关系具有重要意义[5]。
林下植被是林地养分循环系统的重要组成,并在维护生态系统多样性和稳定性方面有突出作用[6]。其对灌木的生长、发育及地下过程如分解、土壤养分流动和积累等均产生很大影响[7]。同时大量研究表明,灌木在林下植被的生存、生长、繁殖等过程中也起到了重要的促进作用,尤其是在极端环境条件下[8-11]。林下植被的生长主要受到环境、灌木类型及发育状况等因素的影响[7,12]。灌草的相互作用如竞争、干扰等被认为是构成植物种群结构的主要方式[13]。但随着灌木年限的增长,林地养分循环系统的结构及组成也会随之变化,同时灌木与林下植被的竞争关系日益凸显,进而影响林下植被的生长及养分分配格局。研究林下植被主要元素的分配及化学计量特征随年限产生的变化有助于阐明植物体的内稳性特征,揭示该区植物生长状况和生境养分供应状况。目前,国内外对于植物生态化学计量学已有大量研究,但多集中于湿地[14]或人造林生态系统[15-16],在高寒沙地生态系统的研究主要集中于土壤特性[17-18]方面,关于林下植被生态化学计量特征方面的研究鲜少。
本研究选取川西北高寒沙地不同年限高山柳灌木林下藏沙蒿(Artemisiawellbyi)、裂叶独活(Heracleummillefolium)和镰荚棘豆(Oxytropisfalcata)3种优势草本植物为对象,以植物生态化学计量学为研究手段,探究不同年限高山柳灌木林下藏沙蒿、裂叶独活和镰荚棘豆叶片和根碳、氮、磷化学计量特征及其变化特征,同时研究3种植物在沙地生态系统中的主要养分限制状况,为提高区域植被覆盖率和改善沙地修复的养分供应措施提供理论依据。
1 材料与方法
1.1 试验地概况
研究区位于四川省阿坝藏族羌族自治州红原县,地处青藏高原东部边缘、四川省西北部,地理坐标为N 31°51′-33°19′、E 101°51′-103°23′,境域分属长江、黄河两大水系。地势为东南向西北倾斜,地貌具有山原向丘状高原过渡的典型特征,平均海拔在3400 m以上;属高原大陆性寒温带季风气候,干湿季节分明,雨热同期,降水主要集中于5-10月,年降水量达 686.75 mm;空气较为稀薄,年均气压为66.56 KPa;气候寒冷,年均温1.1 ℃,极端低温为-33.5 ℃;日照时间较长,年均日照时长约为2158.7 h,太阳辐射强,年总量为6194 MJ·m-2;冰冻期长,寒冻风化作用强烈,季节性冻土分布普遍。植被以藏沙蒿、沙生苔草(Carexpraeclara)、镰荚棘豆、裂叶独活、线叶嵩草(Kobresiacapillifolia)、赖草(Leymussecalinus)、淡黄香青(Anaphalisflavescens)、黑穗苔草(Carexatrata),木里苔草(Carexmuliensis)、细叶亚菊(Ajaniatenuifolia)等为主,植被组合以亚高山草甸为主,沼泽草甸与沼泽植被较为发达,植物群落外貌鲜艳,富有季相变化。
1.2 试验材料与方法
样品采集于2015年8月,在研究区内选择人为活动影响较小的地区作为样品采集地点。通过对红原县高寒草地的实地勘察,在川西北沙化治理典型示范区内共设置了3个年限(6、18、24年)的高山柳实验样地来代表不同年限的高山柳,选取作为林下植被代表,因高山柳林下草本菊科植物藏沙蒿、豆科植物镰荚棘豆、伞形科植物裂叶独活这3种植物在6、18和24年高山柳林下均为优势植物,能够起到代表和对比作用,将其作为研究对象。各年限分别设置3个10 m×10 m的样方, 在植物样方调查基础下采集完整植物样品, 样方基本信息见表1。植物样品取回后,洗掉植株样品根部泥土后快速风干;然后分叶、根装袋,在105 ℃下10 min杀青,最后80 ℃烘干至恒质量不变,称干质量。采用重铬酸钾容量法测定植物样品的有机碳含量,采用半微量凯氏法测定全氮含量,采用钼锑抗比色法测定全磷含量[19]。
1.3 数据处理与分析
采用Microsoft Excel 2003对数据进行处理、整理和绘图,用SPSS 19.0对数据进行差异显著性检验(LSD法)和相关性分析。
2 结果与分析
2.1 不同年限高山柳林下植被C含量变化
叶片C含量表现为藏沙蒿>裂叶独活>镰荚棘豆,三者的C含量范围分别为252.98~322.60 mg·g-1、 66.73~71.34 mg·g-1和203.20~299.29 mg·g-1
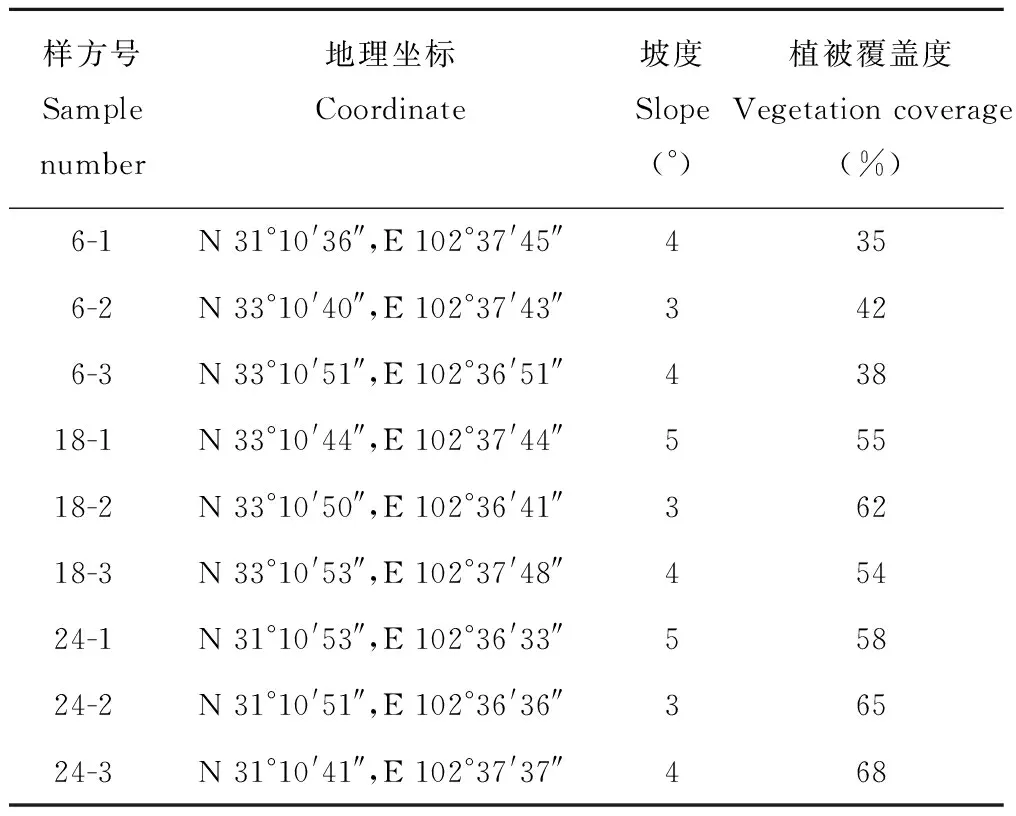
表1 样方基础信息Table 1 Basic information of samples
(图1A)。林下植被叶片C含量随年限增加呈下降趋势,但不同植物间表现出不同变化趋势。藏沙蒿叶片C含量在6年达到最大,后降低并保持稳定,在18和24年间无显著差异;镰荚棘豆叶片C含量在6、18、24年间均无显著差异;裂叶独活叶片C含量随年限增加先增加,在18年时达到最大值后降低,且变化显著(图 1A)。高山柳林下植被根中C含量随年限增加无显著变化,根部C含量表现为裂叶独活>藏沙蒿>镰荚棘豆,其中裂叶独活为387.95~400.95 mg·g-1,藏沙蒿为150.39~180.68 mg·g-1,镰荚棘豆为143.87~164.21 mg·g-1(图 1B)。
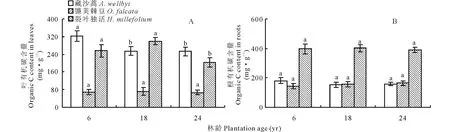
图1 不同年限高山柳林下植被C含量变化 Fig.1 Changes in the understory plants C content in different plantations age of S. cupularis 不同字母表示同一植物不同林龄差异显著(P<0.05),下同。Different lowercase letters in the same plantation indicate significant differences among the plantation age (P<0.05). The same below.
2.2 不同年限高山柳林下植被N含量变化
叶片N含量表现为裂叶独活>藏沙蒿>镰荚棘豆,三者的 N含量范围分别为37.40~52.77 mg·g-1、22.77~47.96 mg·g-1和 13.29~34.70 mg·g-1(图2A)。年限对林下植被叶片N含量影响较显著,且3种植物呈不同的变化趋势。随年限增加,藏沙蒿叶片N含量先下降,在18年达到最低点后略上升;镰荚棘豆随高山柳年限的增长而显著增加(P<0.05),在24年达到最大值;裂叶独活在6、18、24年均为三者最高,且随年限增长先下降,在18年达到最低点后再上升(图 2A)。高山柳林下植被中,镰荚棘豆根部N含量显著高于藏沙蒿和镰荚棘豆,三者分别为20.69~70.04 mg·g-1、20.33~37.47 mg·g-1和1.19~43.46 mg·g-1(图 2B)。
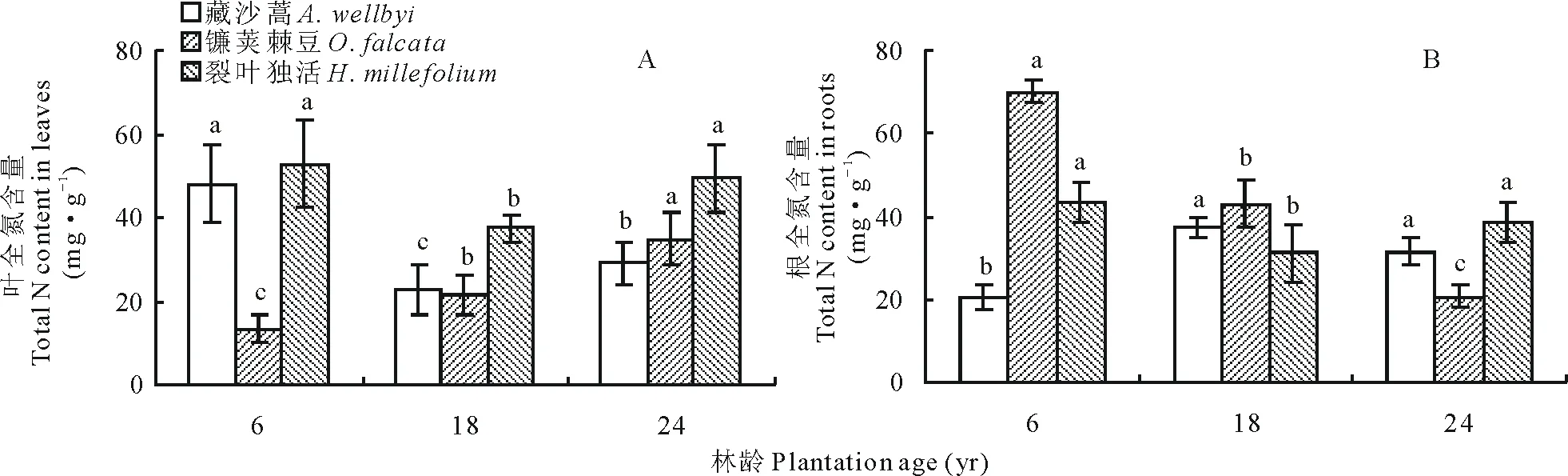
图2 不同年限高山柳林下植被N含量变化Fig.2 Changes in the understory plants N content in different plantations age of S. cupularis
2.3 不同年限高山柳林下植被P含量变化
叶片P含量随年限增长变化显著,裂叶独活、藏沙蒿和镰荚棘豆分别为1.90~2.52 mg·g-1、1.08~3.05 mg·g-1和2.01~2.29 mg·g-1(图 3A)。随着高山柳种植年限增长,其林下植被的叶和根P含量变化均呈相反趋势。藏沙蒿叶全P含量随年限增长先降低再升高,根中P含量随年限增长先升高再降低;镰荚棘豆叶全P含量随年限增长呈增长趋势,其根则逐渐降低;裂叶独活叶全P含量随年限增长呈逐渐降低趋势,其根随年限增长而逐渐升高。其中藏沙蒿叶全P含量随年限增长变化较大,而镰荚棘豆和裂叶独活无显著变化(P>0.05)(图 3B)。

图3 不同年限高山柳林下植被P含量变化Fig.3 Changes in the understory plants P content in different plantations age of S. cupularis
2.4 不同年限高山柳林下植被C∶N
叶片C∶N表现为藏沙蒿>裂叶独活>镰荚棘豆,三者分别为6.73~11.17、4.12~8.00和1.92~5.22(图 4A)。年限对3种草本植物叶片C∶N均有显著影响(P<0.05),藏沙蒿在6年最小,先随年限增长而增加,在18年达到最大值后下降;裂叶独活随年限增长先增加,在18年达到最大值后下降至最小值;镰荚棘豆则随年限增长一直降低,24年时达到最低(图 4)。高山柳林下植被根部C∶N随年限增长变化显著,藏沙蒿随年限增长先由最大值8.89下降至最低值4.01后增加;镰荚棘豆随年限增长而增加,在24年达到最大值7.94;裂叶独活随年限增长先增加至最大值12.86后降低(图 4B)。
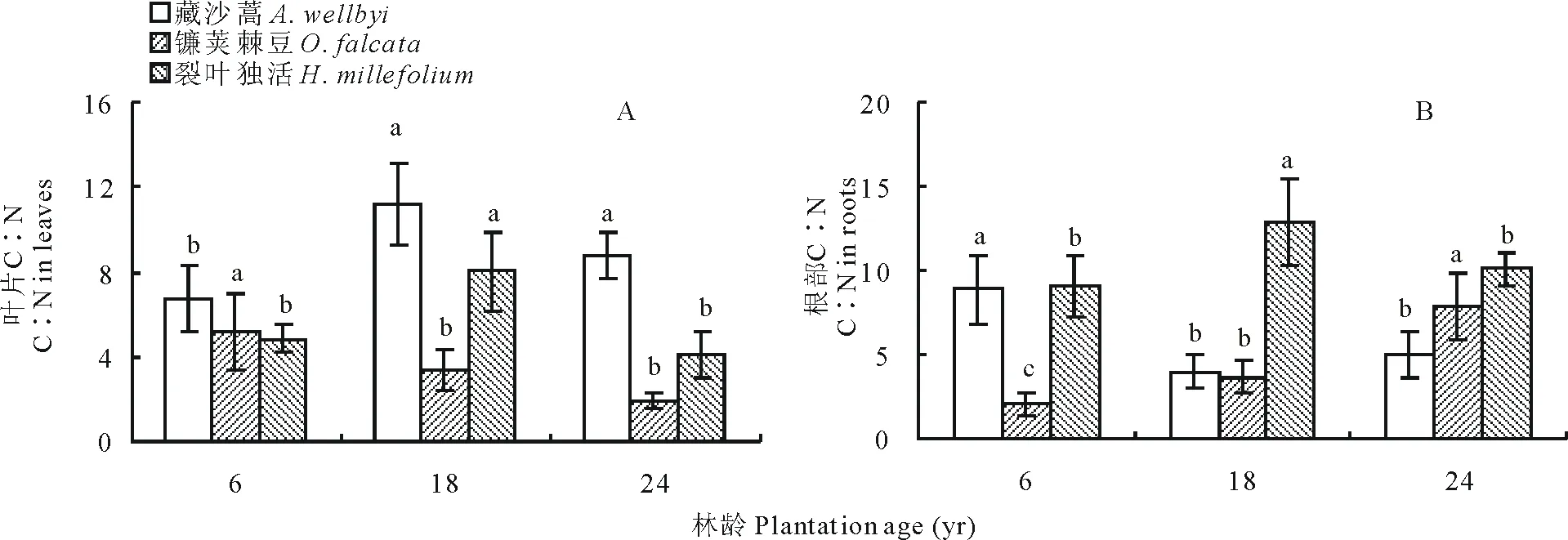
图4 不同年限高山柳林下植被含量C∶N变化Fig.4 Changes in the understory plants C∶N in different plantations age of S. cupularis
2.5 不同年限高山柳林下植被C∶P
高山柳林下植被叶片C∶P表现为藏沙蒿>裂叶独活>镰荚棘豆,藏沙蒿和裂叶独活随年限增长变化显著(P<0.05),藏沙蒿随年限增长由6年时最低值105.65增加至18年时最大值235.66而后再降低;裂叶独活同藏沙蒿一样由最低值101.67先增加至最高值147.58而后降低;镰荚棘豆随年限增长差异不显著,有下降趋势(图5A)。根部C∶P随年限增长变化显著,藏沙蒿随年限增长先下降,在18年降至最小值64.50后增加;裂叶独活随年限增长一直下降,在6年为最大值196.80,24年时为最小值141.68;而镰荚棘豆随年限增长一直增加,在24年达到最大值78.32(图 5B)。
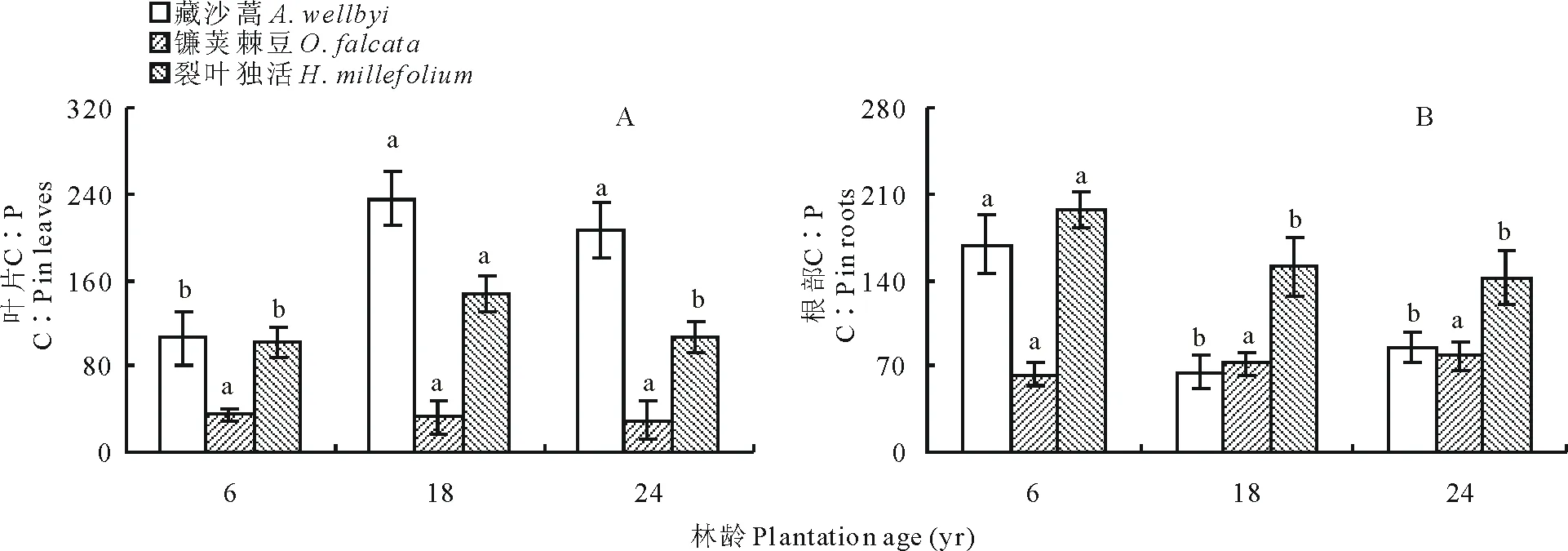
图5 不同年限高山柳林下植被含量C∶P变化Fig.5 Changes in the understory plants C∶P in different plantations age of S. cupularis
2.6 不同年限高山柳林下植被N∶P
叶片N∶P表现为裂叶独活>藏沙蒿>镰荚棘豆,三者分别为18.44~25.89、15.70~23.69和6.60~15.17(图 6A)。年限对3种草本植物均有显著影响(P<0.05),藏沙蒿和镰荚棘豆随年限增长均显著增加,由6年最低值增加至24年最高值;裂叶独活则随年限增长先下降而后增加至最高值(图 6A)。根部N∶P随年限增长变化显著(P<0.05),镰荚棘豆随年限增长显著下降,由6年最高值30.44降至24年最低值9.87;藏沙蒿随年限增长则呈先下降后上升的趋势;裂叶独活随年限增长也由最高值21.58先降至最低值11.76而后增加(图 6B)。

图6 不同年限高山柳林下植被含量N∶P变化Fig.6 Changes in the understory plants N∶P in different plantations age of S. cupularis
3 讨论
植物叶片C∶N、C∶P可以表征植物吸收营养元素同化C的能力,在一定程度上可以反映出单位养分供应量所能达到的生产力、植物对营养的利用效率以及植物的生长速度[20]。Sterner等[21]提出的生长率假说认为,生物体的快速生长需要大量的核糖体RNA合成蛋白质,由于核糖体RNA中含有大量P,从而使得生长率高的生物具有较低的C∶N和C∶P。本研究结果表明,在18和24年高山柳林下植被C∶P明显高于6年,其原因可能是造林过程中灌木和多年生草本植物对N、P的摄取导致土壤中全N和全P含量减少,进而限制了植物吸收摄取。这一研究结果与崔宁洁等[22]对不同林龄马尾松生态化学计量特征研究结果相一致,其研究结果表明,该地区马尾松生长早期土壤相对贫瘠,可供植物利用的N、P养分较少,从而限制了马尾松的生长发育。较高的C∶N和C∶P也表明植物对于N、P元素利用效率较高,这也证实了植物往往以较高的元素利用效率应对贫瘠的养分状态[23]。
Aerts等[24]研究表明,植物对 N、P 养分缺乏的适应可体现在植物N∶P化学计量比的变化上,因此,N∶P化学计量比可作为当前限制性养分判断的指标之一。Drenovsky等[25]研究表明,植物N∶P<14反映植物生长受N限制,N∶P >16反映植物受P限制,14
4 结论
在川西北高山沙地修复过程中,随着灌木高山柳年限的增加,其林下植被藏沙蒿、镰荚棘豆和裂叶独活叶片和根C、N、P含量及其比值发生了显著变化。不同高山柳林下植被N含量差异显著,随着年限增长叶片N含量增加而根部N含量逐渐减少;林下植被P含量呈不同的变化趋势。随着高山柳年限增长,灌木和草本植物对土壤养分的不断摄取,林下植被叶片C∶N和C∶P较高,植被生长受到限制,但同时也反映出在养分供给缺乏的条件下植物对养分的利用效率较高;林下植被N∶P随年限增加而增加,其中不同年限林下藏沙蒿和裂叶独活叶片N∶P均大于16,镰荚棘豆由6.60增长到15.17,由受N限制转为受N和P的共同限制,而林下植被根部N∶P随年限增加而降低,但仍表现出主要受P限制,因而该地区林下植被的生长主要受到P的限制,为提高区域植被覆盖率,改善沙漠化情况应合理使用P肥以改善养分供应状况,且对于年限较大的高山柳林下植物补充P尤为重要。
参考文献References:
[1] Chrzanowski T H, Kyle M, Elser J J,etal. Element ratios and growth dynamics of bacteria in an oligotrophic Canadian shield lake. Aquatic Microbial Ecology, 1996, 11(2): 119-125.
[2] Qi D H, Wen Z M, Wang H X,etal. Stoichiometry traits of carbon, nitrogen, and phosphorus in plants of different functional groups and their responses to micro-topographical variations in the hilly and gully region of the Loess Plateau, China. Acta Ecologica Sinica, 2016, 36(20): 6420-6430.
戚德辉, 温仲明, 王红霞, 等. 黄土丘陵区不同功能群植物碳氮磷生态化学计量特征及其对微地形的响应. 生态学报, 2016, 36(20): 6420-6430.
[3] Niklas K J, Cobb E D. N, P, and C stoichoimetry ofEranthishyemalis(Ranunculaceae) and the allometry of plant growth. American Journal of Botany, 2005, 92(8): 1256-1263.
[4] Li C J, Lei J Q, Xu X W,etal. The stoichiometric characteristics of C, N, P for artificial plants and soil in the hinterland of Taklimakan Desert. Acta Ecologica Sinica, 2013, 33(18): 5760-5767.
李从娟, 雷加强, 徐新文, 等. 塔克拉玛干沙漠腹地人工植被及土壤CNP的化学计量特征. 生态学报, 2013, 33(18): 5760-5767.
[5] Hu W F, Zhang W L, Zhang L H,etal. Stoichiometric characteristics of nitrogen and phosphorus in major wetland vegetation of China. Chinese Journal of Plant Ecology, 2014, 38(10): 1041-1052.
胡伟芳, 章文龙, 张林海, 等. 中国主要湿地植被氮和磷生态化学计量学特征. 植物生态学报, 2014, 38(10): 1041-1052.
[6] Vander Schaaf C L. Estimating understory vegetation response to multi-nutrient fertilization in Douglas-fir and ponderosa pine stands. Journal of Forest Research, 2008, 13(1): 43-51.
[7] Nilsson M C, Wardle D A. Understory vegetation as a forest ecosystem driver: evidence from the northern Swedish boreal forest. Frontiers in Ecology and the Environment, 2005, 3(8): 421-428.
[8] He Q, Cui B, Bertness M D,etal. Testing the importance of plant strategies on facilitation using congeners in a coastal community. Ecology, 2012, 93(9): 2023-2029.
[9] Cavieres L A, Badano E I, Sierra-Almeida A,etal. Positive interactions between alpine plant species and the nurse cushion plantLaretiaacaulisdo not increase with elevation in the Andes of central Chile. New Phytologist, 2006, 169(1): 59-69.
[10] Armas C, Pugnaire F I. Ontogenetic shifts in interactions of two dominant shrub species in a semi-arid coastal sand dune system. Journal of Vegetation Science, 2009, 20(3): 535-546.
[11] Zhou G X, Wang G J, Li L,etal. C, N, P stoichiometry characteristic of Chinese fir root, branch and leaf. Hunan Forestry Science and Technology, 2015, 42(1): 15-18.
周国新, 王光军, 李栎, 等. 杉木根, 枝和叶的 C, N, P 生态化学计量特征. 湖南林业科技, 2015, 42(1): 15-18.
[12] Barbier S, Gosselin F, Balandier P. Influence of tree species on understory vegetation diversity and mechanisms involved-a critical review for temperate and boreal forests. Forest Ecology and Management, 2008, 254(1): 1-15.
[13] Zhang G, Zhao L, Yang Q,etal. Effect of desert shrubs on fine-scale spatial patterns of understory vegetation in a dry-land. Plant Ecology, 2016, 217(9): 1141-1155.
[14] Wang W Q, Xu L L, Zeng C S,etal. Carbon, nitrogen and phosphorus ecological stoichiometric ratios among live plant-litter-soil systems in estuarine wetland. Acta Ecologica Sinica, 2011, 31(23): 7119-7124.
王维奇, 徐玲琳, 曾从盛, 等. 河口湿地植物活体-枯落物-土壤的碳氮磷生态化学计量特征. 生态学报, 2011, 31(23): 7119-7124.
[15] Wang S Q, Yu G R. Ecological stoichiometry characteristics of ecosystem carbon, nitrogen and phosphorus elements. Acta Ecologica Sinica, 2008, 28(8): 3937-3947.
王绍强, 于贵瑞. 生态系统碳氮磷元素的生态化学计量学特征. 生态学报, 2008, 28(8): 3937-3947.
[16] Fan H, Wu J, Liu W,etal. Linkages of plant and soil C∶N∶P stoichiometry and their relationships to forest growth in subtropical plantations. Plant and Soil, 2015, 392(1/2): 127-138.
[17] Hu Y F, Peng J J, Deng L J,etal. Influences of fencing and planting branchy tamarisk on soil particles composition and mineral nutrients in desertization land in northwestern Sichuan Province. Chinese Journal of Soil Science, 2015, (1): 54-61.
胡玉福, 彭佳佳, 邓良基, 等. 围栏种植红柳对川西北高寒沙地土壤颗粒组成和矿质养分的影响. 土壤通报, 2015, (1): 54-61.
[18] Feng H L, Hu Y F, Shu X Y,etal. Influences of adding needle mushroom edible fungi residues on vegetation and soil enzyme activity of sandy soil in northwest Sichuan. Journal of Soil, 2016, (5): 161-165.
冯慧翎, 胡玉福, 舒向阳, 等. 金针菇菌渣对川西北高寒沙地植被及土壤酶活性的影响. 水土保持学报, 2016, (5): 161-165.
[19] He J S, Han X G. Ecological stoichiometry: Searching for unifying principles from individuals to ecosystems. Chinese Journal of Plant Ecology, 2010, 34(1): 2-6.
贺金生, 韩兴国. 生态化学计量学: 探索从个体到生态系统的统一化理论. 植物生态学报, 2010, 34(1): 2-6.
[20] Liu W D, Su J R, Li S F,etal. Stoichiometry study of C, N and P in plant and soil at different successional stages of monsoon evergreen broad-leaved forest in Pu’er, Yunnan Province. Acta Ecologica Sinica, 2010, (23): 6581-6590.
刘万德, 苏建荣, 李帅锋, 等. 云南普洱季风常绿阔叶林演替系列植物和土壤 C, N, P 化学计量特征. 生态学报, 2010, (23): 6581-6590.
[21] Sterner R W, Elser J J. Ecological stoichiometry: the biology of elements from molecules to the biosphere. Princeton: Princeton University Press, 2002.
[22] Cui N J, Liu X B, Zhang D J,etal. The distribution pattern of carbon, nitrogen and phosphorus and the stoichiometry characteristics ofPinusmassonianaplantation in different age. Ecology and Environmental Science, 2014, 23(2): 188-195.
崔宁洁, 刘小兵, 张丹桔, 等. 不同林龄马尾松(Pinusmassoniana)人工林碳氮磷分配格局及化学计量特征. 生态环境学报, 2014, 23(2): 188-195.
[23] Bowman W D. Accumulation and use of nitrogen and phosphorus following fertilization in two alpine tundra communities. Oikos, 1994, 70(2): 261-270.
[24] Aerts R, Chapin F S. The mineral nutrition of wild plants revisited: a re-evaluation of processes and patterns. Advances in Ecological Research, 1999, 30(8): 1-67.
[25] Drenovsky R E, Richards J H. Critical N∶P values: predicting deficiencies in desert shrublands. Plant and Soil, 2004, 259(1): 59-69.
[26] Dong W Y, Nie L S, Li J Y,etal. Effects of nitrogen forms on the absorption and distribution of nitrogen inPopulustomentosaseedlings using the technique of15N tracing. Journal of Beijing Forestry University, 2009, (4): 97-101.
董雯怡, 聂立水, 李吉跃, 等. 应用15N示踪研究毛白杨苗木对不同形态氮素的吸收及分配. 北京林业大学学报, 2009, (4): 97-101.
[27] Zhang C H, Wang Z M, Ju W M,etal. Spatial and temporal variability of soil C/N ratio in Songnen Plain Maize Belt. Environmental Science, 2011, 32(5): 1407-1414.
张春华, 王宗明, 居为民, 等. 松嫩平原玉米带土壤碳氮比的时空变异特征. 环境科学, 2011, 32(5): 1407-1414.
猜你喜欢
鸭绿江(2021年35期)2021-11-11 15:25:02
学生天地(2020年31期)2020-06-01 02:32:08
疯狂英语·初中天地(2019年3期)2019-07-28 07:45:48
疯狂英语·初中天地(2019年2期)2019-07-28 07:39:44
疯狂英语·初中天地(2019年1期)2019-07-28 07:32:40
军事文摘(2018年24期)2018-12-26 00:57:56
反射疗法与康复医学(2017年7期)2017-01-16 01:11:44
反射疗法与康复医学(2017年7期)2017-01-16 01:11:42
西部广播电视(2015年7期)2016-01-16 03:45:18
天然产物研究与开发(2014年1期)2014-04-27 14:15:08
