
温度和土壤水分与Bt棉铃壳中杀虫蛋白表达关系及其氮代谢生理机制
2018-04-18 06:35:49张祥王剑彭盛芮秋治李丽楠陈媛陈源陈德华
中国农业科学 2018年7期
张祥,王剑,彭盛,芮秋治,李丽楠,陈媛,陈源,陈德华
温度和土壤水分与Bt棉铃壳中杀虫蛋白表达关系及其氮代谢生理机制
张祥,王剑,彭盛,芮秋治,李丽楠,陈媛,陈源,陈德华
(扬州大学/江苏省作物遗传生理重点实验室,江苏扬州 225009)
【】明确温度和土壤水分对Bt棉杀虫蛋白含量及其氮代谢活性的影响,为生产中Bt棉抗虫性的安全稳定利用提供理论参考。【】2016—2017年以转Bt抗虫基因抗虫棉常规品种泗抗1号(SK1)和杂交种泗抗3号(SK3)为材料,采用盆栽法,设置29℃、32℃、35℃、38℃ 4个温度水平,土壤最大持水量的80%、70%、60%、50%、40% 5个土壤水分水平,观察温度和土壤水分对Bt棉铃壳杀虫蛋白含量的影响,各处理持续胁迫4 d。2016年主要研究各处理对Bt棉铃壳中杀虫蛋白含量的影响;在此基础上,2017年进一步探讨各处理对铃壳中可溶性蛋白含量、谷氨酸草酰乙酸转氨酶(GOT)、蛋白酶、肽酶活性的影响。【】SK1和SK3杀虫蛋白含量均在32℃、最大持水量为60%时最高,分别达到471.1 ng·g-1FW和351.7 ng·g-1FW。在同一土壤水分条件下,32℃最利于SK1和SK3杀虫蛋白表达;同一温度条件下,最大持水量60%利于SK1和SK3杀虫蛋白表达。对杀虫蛋白含量与温度和土壤水分关系进行二元多项式回归分析发现,Bt棉杀虫蛋白含量(Y)与温度(X2)和土壤水分(X1)呈二元二次方程关系,其中SK1、SK3相关方程分别为Y=-3230.2+17.2X1+199.1X2-0.3X12-3.7X22-0.7X1X2(=0.829**)、Y=-3322.0+40.7X1+145.2X2-0.3X12-2.0X22-0.3X1X2(=0.739**)。SK1的杀虫蛋白表达量最大的温度和土壤水分条件为31.8℃、57.8%,SK3为33.2℃、60.8%。氮代谢相关生理特征表明,SK1和SK3均表现为在32℃和土壤含水量为60%处理下,棉铃中可溶性蛋白含量、谷氨酸草酰乙酸转氨酶(GOT)活性较高,蛋白酶、肽酶活性较低;杀虫蛋白含量与可溶性蛋白和GOT活性呈极显著正相关关系(=0.613**;=0.735**),与蛋白酶活性和肽酶活性呈极显著负相关关系(=-0.724**;=-0.738**)。【】温度和土壤水分通过调控蛋白质分解和合成,共同影响Bt棉杀虫蛋白表达,且与其含量呈二元二次方程关系。
Bt棉;温度;土壤水分;杀虫蛋白;氮代谢
0 引言
【研究意义】1997年起中国开始在生产上大面积推广应用(Bt)棉,自那以来中国的转基因抗虫棉种植面积不断扩大。据统计,Bt转基因抗虫棉的种植面积已经达到中国总植棉面积的80%以上[1-2]。但目前为止,Bt棉的杀虫蛋白表达不稳定的观点受到国内外一致认同。其中环境因素的胁迫对抗虫性表达具有较大的影响,明确环境因素对Bt棉杀虫蛋白表达影响及其机制,可为逆境下Bt棉抗虫性安全应用和合理防治棉铃虫提供科学依据。【前人研究进展】在不良的环境条件胁迫下,Bt棉显著表现为抗虫性下降或不抗虫的现象[3-5]。温度是影响Bt棉抗虫性表达的重要因素[6-7]。Zhang等[8]、夏兰芹等[9]研究表明高温逆境可导致Bt杀虫蛋白含量急剧降低。Wang等[10]进一步明确38℃是导致Bt棉棉蕾中Bt杀虫蛋白表达量显著下降的临界温度。在水分方面,王留明等[11]的研究表明,在花铃期进行的田间淹水涝渍或土壤缺水干旱处理可导致棉株各个器官中Bt蛋白表达含量显著降低,其中缺水干旱能够更强地抑制器官中Bt杀虫蛋白的合成。Benedict等[12]发现土壤含水量下降,引起土壤水分压力降低,从而导致Bt棉植株中总可溶性蛋白及Bt杀虫蛋白含量减低。【本研究切入点】前人的研究或推测主要是以单个因子为主,而棉花生育过程并非仅受简单的单因素影响,而是受自然环境条件中温度和土壤水分等数个条件互作综合影响,因此单因子往往不能完全反映自然温度(高温或低温)和土壤水分(水分过多或过少)互作条件下对Bt棉抗虫性表达的影响。此外,虽然前人主要研究了环境因子中温度和土壤水分对Bt棉杀虫蛋白表达影响,但并未能明确它们与杀虫蛋白含量的数量关系,这限制了上述研究成果对生产中的实际指导作用。【拟解决的关键问题】本文拟通过人工模拟棉花生育过程中经常出现的温度与土壤水分互作条件,探讨Bt棉杀虫蛋白表达量的变化,明确Bt棉杀虫蛋白含量与温度和土壤水分数量关系及其生理机制,从而为逆境条件下,Bt棉抗虫性安全应用提供理论和实践依据。
1 材料与方法
1.1 试验材料与试验设计
试验于2016—2017年在扬州大学江苏省作物遗传生理国家重点实验室培育点盆栽试验场进行。选择来源于中国的转Bt基因棉品种泗抗1号(常规种)和泗抗3号(杂交种)为材料,由江苏省宿迁市农业科学院提供,抗虫基因均为。
试验采用盆栽法,所用盆钵直径40 cm,盆高50 cm,每盆装20 kg过筛的砂壤土(取自试验田)。采用育苗移栽方式种植,于4月6日播种,出苗41 d后,选择长势一致的壮苗移栽至盆钵中,每盆1株。其他生长管理措施按大田高产栽培要求进行。
于盛铃期设温度与土壤水分2个试验因子,其中温度设4个水平(24 h恒温):29℃、32℃、35℃、38℃;土壤水分设5个水平:土壤含水量分别为土壤最大持水量的80%、70%、60%、50%、40%,分别用80%、70%、60%、50%、40%表示。每个处理重复3次,供试土壤最大田间持水量为23.5%。2016年主要研究温度和土壤水分对Bt杀虫蛋白含量影响,2017年在2016年结果基础上进一步研究其对氮代谢相关指标影响。
应用人工气候室(北京易盛泰和科技有限公司,北京)控制温度水平(精度±0.5℃),采用菲利普T5灯和美国GE农用钠灯控制光源,光照30 000 Lux。各处理在盛铃期前10 d(2016年和2017年均为7月30日),标记棉株内围第1—2果节当日花。同时控制浇水,如遇下雨,将处理盆钵移入室内。使用WET土壤三参数速测仪监测土壤水分,用称重法控制土壤水分,即当监测发现土壤水分低于设计值时,分别于6:00、14:00、22:00进行定量补水。在到达预期水分指标后,将盆栽棉株移入人工气候箱进行处理。处理持续4 d,胁迫结束后分别取样标记的相同日龄的铃,液氮冷冻后进行相关指标测定。
1.2 测定内容和方法
1.2.1 Bt杀虫蛋白含量 采用酶联免疫分析法(ELISA)测定,试剂盒由中国农业大学提供。测定方法参见文献[13]。
1.2.2 氮代谢相关酶活性和物质含量 测定铃壳中谷氨酸草酰乙酸转氨酶(GOT)、蛋白酶、肽酶活性、可溶性蛋白含量。GOT、蛋白酶、肽酶活性用邹琦[14]方法测定,可溶性蛋白含量用张宪政[15]方法测定。
1.3 数据处理与分析
采用Excel 2007进行数据处理和作图,使用SPSS10.0进行统计分析。利用MATLAB软件进行杀虫蛋白与温度和土壤水分二元二次方程(Y=-b0+b1X1+ b2X2+b3X12+b4X22+b5X1X2)模拟及作图。
2 结果
2.1 温度和土壤水分对生理指标影响方差分析
从表1的双因素方差分析结果可以看出,单独温度、水分以及温度与水分交互作用均对供试品种铃壳中杀虫蛋白和可溶性蛋白含量,GOT、蛋白酶和肽酶活性具有显著或极显著影响。
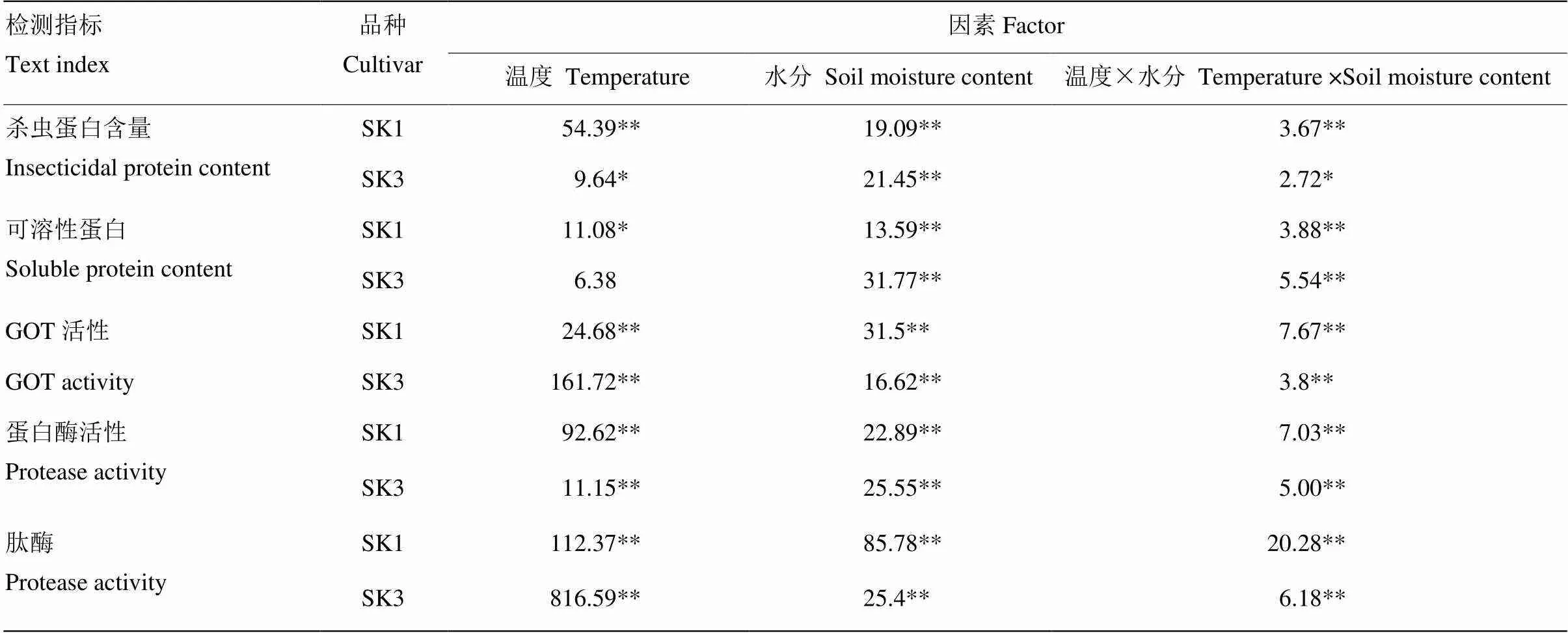
表1 双因素方差分析
值代表均方和;*和**分别代表显著相关(<0.05)和极显著相关(<0.01);SK1和SK3分别代表泗抗1号和泗抗3号
Value indicates mean square; *, ** indicate significance at 0.05 and 0.01 levels, respectively. SK1 and SK3 indicate Sikang 1 and Sikang 3, respectively
2.2 温度和土壤水分对杀虫蛋白含量影响
图1表明,在同一温度条件下,Bt棉棉铃中杀虫蛋白含量均随着土壤水分含量的增加而呈先增加后下降的趋势。总体而言,SK1和SK3均表现为土壤水分含量为最大持水量的60%时,棉铃中杀虫蛋白含量最高。在同一土壤水分条件下,棉铃中杀虫蛋白含量也随着温度的升高呈先上升再下降趋势,在32℃达峰值。从温度和土壤水分综合效应来看,在32℃和土壤含水量为60%条件下时,SK1和SK3棉铃杀虫蛋白含量最高,分别为471.1 ng·g-1FW和351.7 ng·g-1FW,且总体显著高于其他处理,说明此环境条件最有利于SK1和SK3杀虫蛋白表达。

图中小写字母表示同一品种各处理间在5%水平差异显著。下同
对Bt棉铃壳中杀虫蛋白含量(Y)与土壤水分含量(X1)和温度(X2)进行建模拟合发现,SK1和SK3杀虫蛋白含量与土壤水分含量和温度均呈二元二次方程关系(图2),拟合方程分别为:Y=-3230.2+17.2X1+ 199.1X2-0.3X12-3.7X22-0.7X1X2;Y=-3322.0+40.7X1+ 145.2X2-0.3X12-2.0X22-0.3X1X2。相关系数分别为0.829、0.739,均达到极显著水平,说明上述方程拟合效果很好。根据上述方程计算,最有利于SK1和SK3棉铃中Bt蛋白表达的温度和土壤水分含量分别为31.2℃、最大持水量的57.8%和33.2℃、最大持水量的60.8%。
2.3 温度和土壤水分对Bt棉氮代谢生理特征影响
2.3.1 可溶性蛋白含量 方差分析表明(表1),温度和土壤水分均对SK1和SK3棉铃中可溶性蛋白表达具有显著或极显著影响,且两者存在着极显著互作关系。由图3可知,在同一温度条件下,Bt棉棉铃中可溶性蛋白含量均随着土壤水分含量的增加而呈先增加后下降的趋势。总体而言,SK1和SK3均表现为土壤水分含量为最大持水量的50%—70%时,棉铃中可溶性蛋白含量较高。在同一土壤水分条件下,棉铃中可溶性蛋白含量也随着温度的升高呈先上升后下降规律,在32℃达峰值。从温度和土壤水分综合效应来看,2个品种均在32℃和土壤含水量为60%处理时,棉铃中可溶性蛋白含量较高,分别为2.87 mg·g-1FW和2.64 mg·g-1FW。

图2 温度和土壤水分与Bt棉铃壳的杀虫蛋白的关系(2016)

图3 温度和水分对Bt棉可溶性蛋白含量影响(2017)
2.3.2 GOT活性 温度和土壤水分对棉铃中GOT活性影响与可溶性蛋白趋势基本一致(图4)。表1表明,单独温度和土壤水分均极显著影响SK1和SK3棉铃中GOT活性,且两者存在着极显著互作关系。进一步分析发现,在同一温度条件下,Bt棉棉铃中GOT活性均随着土壤水分含量的增加而呈先增加后下降的趋势。总体而言,SK1和SK3均表现为土壤水分含量为最大持水量的60%时,棉铃中GOT活性较高。在同一土壤水分含量条件下,GOT活性也随着温度的增加呈先上升,至32℃达最高,后下降的趋势。从温度和土壤水分综合效应来看,2个供试品种均在32℃和土壤含水量为60%条件下棉铃中GOT活性最高,分别达2.14 mol·g-1·30min-1和2.44 mol·g-1·30min-1,且显著高于其他处理。综合图3和图4而言,32℃和土壤含水量为60%处理最利于SK1和SK3氮代谢合成能力提高。
2.3.3 蛋白酶活性 方差分析(表1)表明,单独温度和土壤水分均极显著影响SK1和SK3棉铃中蛋白酶活性,且两者存在着极显著互作关系。但温度和土壤水分对棉铃中蛋白酶活性影响与可溶性蛋白、GOT活性变化趋势相反(图5)。其中,在同一温度条件下,Bt棉棉铃中蛋白酶活性均随着土壤水分含量的增加而呈先降低后上升的趋势。总体而言,SK1和SK3均表现为土壤水分含量为最大持水量的50%—60%时,棉铃中蛋白酶活性最低。在同一土壤水分含量条件下,棉铃中蛋白酶活性也随着温度的升高呈先上升再下降趋势。在不同土壤水分条件下,均以32℃条件下活性较低。从温度和土壤水分综合效应来看,2个品种均在32℃和土壤含水量为60%处理时,棉铃中蛋白酶活性最低,分别仅为0.41 mg·g-1·h-1和0.05 mg·g-1·h-1,且显著低于其他处理。
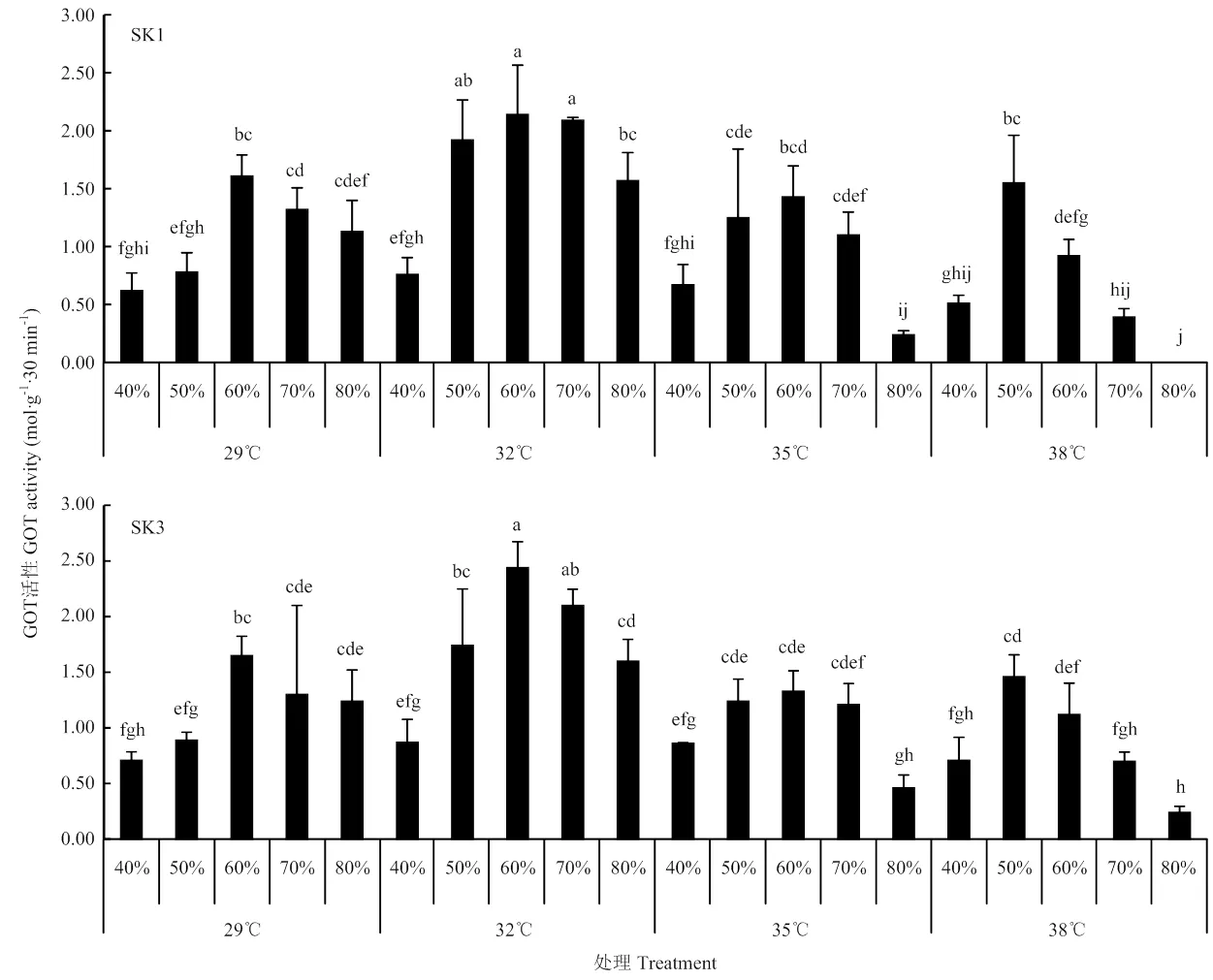
图4 温度和水分对Bt棉GOT活性影响(2017)
2.3.4 肽酶活性 温度和水分对棉铃中肽酶活性影响与蛋白酶活性变化趋势相似(图6)。单独温度和土壤水分均极显著影响SK1和SK3棉铃中肽酶活性,且两者存在着极显著互作关系(表1)。在同一温度条件下,Bt棉棉铃中肽酶活性均随着土壤水分含量的增加而呈先降低后上升的趋势。总体而言,SK1和SK3均表现为土壤水分含量为最大持水量的50%—60%时,棉铃中肽酶活性最低。在同一土壤水分含量条件下,29℃和32℃条件下,肽酶活性总是保持在一个较低的水平。从温度和土壤水分综合效应来看,2个品种均在32℃和土壤含水量为60%处理时,棉铃中肽酶活性最低,且显著低于其他处理。综合图5和图6结果表明,32℃和土壤含水量为60%时SK1和SK3氮代谢分解能力最低。

图5 温度和水分对Bt棉蛋白酶活性影响(2017)
2.4 杀虫蛋白含量与氮代谢相关酶活性相关性
图7表明,不同温度和土壤水分条件下,2个类型品种盛铃期棉铃中杀虫蛋白含量与可溶性蛋白和GOT活性呈极显著正相关关系(=0.613**;=0.735**),与蛋白酶活性和肽酶活性呈极显著负相关关系(=-0.724**;=-0.738**)。以上结果说明在不同温度和土壤水分条件下,棉铃中杀虫蛋白含量与氮代谢密切相关,增强棉铃中蛋白合成能力,降低蛋白分解能力有利于Bt棉杀虫蛋白表达量增加。
3 讨论
3.1 协调温度和土壤水分可利于Bt棉杀虫蛋白表达
前人研究表明,外界环境因子对Bt棉杀虫蛋白表达具有明显影响[16-17],但其研究或推测主要是以单个因子进行的,而棉花生育过程中是受温度和土壤水分等多种因素的综合影响,因此单因子往往不能完全反映自然环境中温度和土壤水分亏缺互作条件下对Bt棉抗虫性表达的影响。Chen等[18]研究高温逆境对Bt棉Bt杀虫蛋白表达量影响的结果表明,在温度为37℃时,花铃期Bt棉叶片中杀虫蛋白表达量在48 h后显著下降。进一步研究发现,这样的结果是发生在相对湿度为50%的高温低湿条件下,当相对湿度提高至70%,温度仍为37℃时,Bt杀虫蛋白含量下降明显减少,仅为50%相对湿度时的一半,说明温度与湿度对Bt抗虫蛋白表达的影响存在互作效应。Zhou等[19]则明确认为温度与土壤水分互作可明显影响Bt棉抗虫性表达,说明温度与土壤水分对Bt棉杀虫蛋白表达的影响存在互作效应。本文研究结果也证实温度和土壤水分对Bt棉棉铃中杀虫蛋白表达具有显著或极显著的互作作用。其中,在本试验温度和土壤水分范围内,泗抗1号和泗抗3号在32℃和土壤含水量为最大持水量的60%条件下,棉铃中杀虫蛋白含量最高。进一步对泗抗1号和泗抗3号铃壳的杀虫蛋白含量与温度、土壤水分含量之间的关系进行回归分析和建立模型发现,杀虫蛋白与温度和土壤水分含量呈二元二次方程关系。而且拟合的方程相关系数均达到极显著水平(=0.829**、0.739**),说明该方程拟合效果很好,计算明确最有利于SK1和SK3棉铃中Bt蛋白表达的温度和土壤水分含量分别为31.2℃、最大持水量的57.8%和33.2℃、最大持水量的60.8%。因此,在生产中,我们可利用该方程计算出不同温度和土壤水分情况下Bt棉杀虫蛋白表达量,明确Bt棉自身对田间害虫的控制效果,进而为田间农药施用量提供依据。此外,田间管理中人们还可以利用开沟排水或合理灌溉等方法缓解高温和干旱的影响,从而利于Bt棉自身杀虫性的表达。
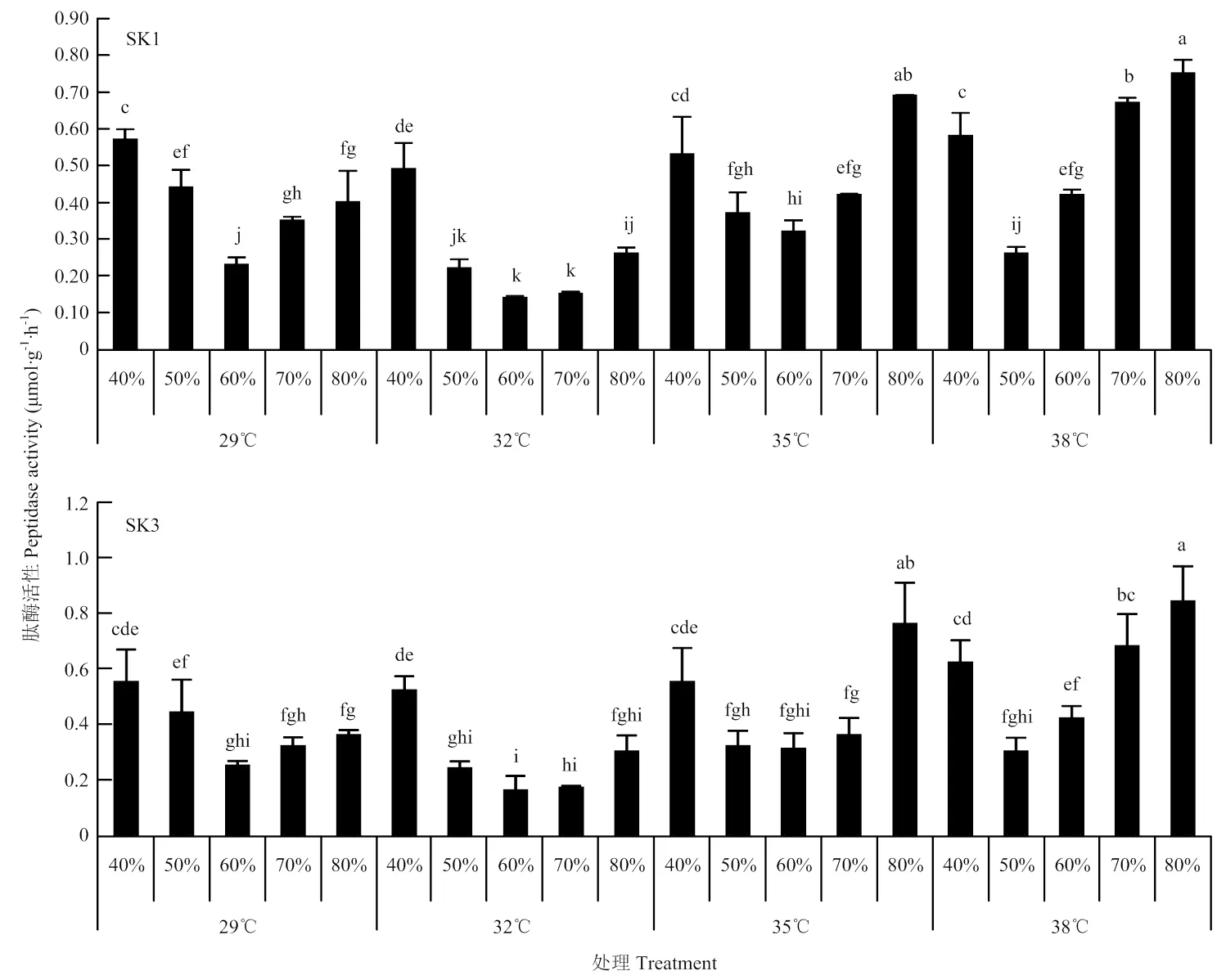
图6 温度和水分对Bt棉肽酶活性影响(2017)

图7 Bt棉杀虫蛋白含量与氮代谢相关酶活性相关性(2017)
3.2 Bt棉杀虫蛋白表达水平受蛋白质分解和合成能力调节
虽然环境影响Bt棉抗虫性表达的机理目前还未明确,但主要有以下几种推断:第一种推断认为逆境使得Bt的启动子甲基化失活,从而导致Bt表达关闭(switch off)[20-21];第二种推断认为不良的环境条件下棉株体内的杀虫蛋白与产生的单宁等物质结合而失活[22-23];第三种推断认为不良环境条件下蛋白质合成下降,导致杀虫蛋白表达量下降[24-25]。其中GOT活性及可溶性蛋白是植物体内氮代谢合成的关键酶和物质,而蛋白酶和肽酶则是氮代谢分解关键酶。张桂玲等[26]研究表明短期盐胁迫后,Bt棉内游离氨基酸含量与Bt杀虫蛋白含量呈极显著负相关,可溶性蛋白含量与Bt杀虫蛋白呈极显著正相关。王永慧等[27]发现,湿度胁迫会导致盛铃期叶片Bt杀虫蛋白含量的下降,这可能是因为胁迫导致Bt 杀虫蛋白的合成能力下降、分解能力上升。陈德华等[28]研究也发现Bt棉氮代谢活性显著影响杀虫蛋白表达的影响。在本文中,笔者发现在32℃和土壤含水量为最大持水量的60%条件下,供试品种泗抗1号和泗抗3号GOT活性和可溶性蛋白含量均显著高于其他处理,而蛋白酶和肽酶活性则显著低于其他处理。相关性分析发现,在不同温度和土壤水分条件下,GOT活性和可溶性蛋白含量与Bt棉棉铃中杀虫蛋白含量呈极显著正相关关系;蛋白酶和肽酶活性则与杀虫蛋白含量呈极显著负相关关系(图7)。由此可见,温度和土壤水分环境主要是通过调节Bt棉蛋白质合成和分解来影响棉铃中杀虫蛋白表达。但不同品种间相比,泗抗3号杀虫蛋白表达量最高时的温度和土壤水分含量均高于泗抗1号,这可能是由于杂交种具有较高的抗逆性(如高温等)[29],其在逆境下仍能保持较高的氮代谢合成能力和较低的氮代谢分解能力,这与前人研究基本一致[30-31]。因此,在Bt棉生长过程中,我们应通过相应的栽培措施,促进棉铃中蛋白质合成相关酶活性或物质含量,降低与蛋白质分解相关酶活性或物质含量,来提高Bt棉杀虫蛋白表达和其自身抗虫性。
4 结论
Bt棉铃壳中杀虫蛋白含量与温度和土壤水分含量呈二元二次函数关系,且最有利于泗抗1号(SK1)和泗抗3号(SK3)棉铃中Bt蛋白表达的温度和土壤水分含量分别为31.2℃、最大持水量的57.8%和33.2℃、最大持水量的60.8%。进一步分析发现温度和土壤水分主要是通过调控蛋白质分解与合成能力影响杀虫蛋白表达。
[1] JAMES C. Preview: Global status of commercialized biotech/GM crops., 2004(32): 1-12.
[2] JAMES C. 2014年全球生物技术/转基因作物商业化发展态势. 中国生物工程杂志, 2015, 35(1): 1-14.
JAMES C. The development trend of global biotechnology/GM crops commercialization in 2014., 2015, 35(1): 1-14. (in Chinese)
[3] JAGGER R. The company view on Bt cotton., 1997(1): 26.
[4] KAISER J. Pest overwhelm Bt cotton crop.1996, 273(5274): 423.
[5] 刘耀武, 刘洪春, 付桂月, 李宏华, 孙福燕. 近年抗虫棉抗虫性下降原因分析及对策. 中国植保导刊, 2008, 28(1): 30-31.
LIU Y W, LIU H C, FU G Y, LI H H, SHUN F YReason analyzed and counter measures of the decline of insect-resistance for anti-insect cotton in recent years., 2008, 28(1): 30-31. (in Chinese)
[6] LUBNA B. 不同类型Bt 棉花的抗虫性及棉铃虫对Bt 蛋白的抗性动态监测[D]. 北京: 中国农业科学院, 2014.
LUBNA B. Resistance of different transgenic Bt cotton and monitoring the resistant dynamics of(Hubner) to Bt toxin[D]. Beijing: Chinese Academy of Agricultural Sciences, 2014. (in Chinese)
[7] 周冬生, 吴振廷, 王学林, 倪春耕, 郑厚今, 夏静. 施肥量和环境温度对转Bt基因棉抗虫性的影响. 安徽农业大学学报, 2000, 27(4): 352-357.
ZHOU D S, WU Z T, WANG X L, NI C G, ZHEN H J, XIA J. Influence of fertilization and environmental temperature on the resistance of Bt transgenic cotton to cotton bollworm., 2000, 27(4): 352-357.
[8] ZHANG X, WANG G X, LÜ C H, CHEN Y, CHEN D H. The effect of low temperature on the insecticidal properties of Bt cotton.,, 2012, 10(3): 397-403.
[9] 夏兰芹, 郭三堆. 高温对转基因抗虫棉中Bt杀虫基因表达的影响. 中国农业科学, 2004, 37(11): 1733-1737.
XIA L Q, GUO S D. The expression of Bt toxin gene under different thermal treatments., 2004, 37(11): 1733-1737. (in Chinese)
[10] WANG J, CHEN Y, YAO M H, LI Y, WEN Y J, CHEN Y, ZHANG X, CHEN D H. The effects of high temperature level on square Bt protein concentration of Bt cotton., 2015, 14(10): 1971-1979.
[11] 王留明, 王家宝, 沈法富, 张学坤, 刘任重. 渍涝与干旱对不同转Bt基因抗虫棉的影响. 棉花学报, 2001, 13(2): 87-90.
WANG L M, WANG J B, SHEN F F, ZHANG X K, LIU R Z. Influences of water logging and drought on different transgenic Bt cotton cultivars., 2001, 13(2): 87-90. ( in Chinese)
[12] BENEDICT J H, SACHS E S, ALTMAN D W, DEATON W R, KOHEL R J, RING D R, BERBERICH S A. Field performance of cottons expressing transgenic Cry1A insecticidal proteins for resistance toand(Lepidoptera: Noctuidae)., 1996, 89(1): 230-238.
[13] 陈松, 吴敬音, 何小兰, 黄骏麒, 周宝良, 张荣铣. 转基因抗虫棉组织中Bt毒蛋白表达量的ELISA测定. 江苏农业学报, 1997, 13(3): 154-156.
CHEN S, WU J Y, HE X L, HUANG J Q, ZHOU B L, ZHANG R X. Quantification using ELISA ofinsecticidal protein expressed in the tissue of transgenic insect-resistant cotton., 1997, 13(3): 154-156. (in Chinese)
[14] 邹琦. 植物生理学实验指导, 北京: 中国农业出版社, 2000; 127-130.
ZOU Q.. Beijing: China Agriculture Press, 2000: 127-130. (in Chinese)
[15] 张宪政. 作物生理研究法. 北京: 农业出版社, 1990: 200-201.
ZHANG X Z.. Beijing: Agricultural Publishing House, 1990: 200-201. (in Chinese)
[16] Hallikeri S S, Halemani H L, Patil B C, Nandagavi R A. Influence of nitrogen management on expression of cry protein in Bt-cotton ()., 2011, 56(1): 62-67.
[17] Martins C M, Beyene G, Hofs J L, Krueger K, Vander V C, Schlueter U, Kunert K J. Effect of water-deficit stress on cotton plants expressing thetoxin., 2008, 152(2): 255-262.
[18] CHEN Y, WEN Y J, CHEN Y, JOHN T C, ZHANG X, WANG Y H, WILLIAM A P, CHEN D H. Effects of extreme air temperature and humidity on the insecticidal expression level of Bt cotton., 2012, 11(11): 1836-1844.
[19] ZHOU G S, WANG Y H, ZHAI F Y, LU S Y, NIMIR A E, PAN H, LÜ D M. Combined stress of low temperature and flooding affects physiological activities and insecticidal protein content in transgenic Bt cotton., 2015, 66(7): 740-746.
[20] STAM M, MOL J N M, KOOTER J M. The silence of genes in transgenic plants., 1997, 79(1): 3-12.
[21] DONG H Z, LI W J. Variability of endotoxin expression in Bt transgenic cotton., 2007, 193(1): 21-29.
[22] FINNEGAN E J, LIEWELLYN D J, FITT G P. What’s happening to expression of the insect protection in field-grown ingard cotton. 11th Australian Cotton Conference. Brisbane, 2002: 291-297.
[23] HOLT H. Season-long monitoring of transgenic cotton plants-development of an assay for the quantification of Bacillus thuringiensis insecticidal Crystal protein. Australian Cotton Growers Research Association. Narrabri NSW, Australia, 1998.
[24] CHEN D H, YE G Y, Yang C Q, CHEN Y, WU Y K. Effect of introducinggene on nitrogen metabolism in cotton., 2005, 92(1): 1-9.
[25] CHEN D H, YE G Y, YANG C Q, CHEN Y, WU Y K. The effect of high temperature on the insecticidal properties of Bt cotton., 2005, 53(3): 333-342.
[26] 张桂玲, 温四民. 盐胁迫对转Bt 基因棉苗期Bt 蛋白表达量和氮代谢的影响. 西北农业学报, 2011, 20(6): 106-109.
ZHANG G L, WEN S M. Effects of salt stress on Bt protein content and nitrogen metabolism of transgenic Bt cotton., 2011, 20(6): 106-109. (in Chinese)
[27] 王永慧, 陈建平, 张祥, 陈源, 陈德华. 温湿度胁迫对Bt 棉叶片氨基酸组分及Bt 蛋白含量的影响. 华北农学报, 2012, 27(6): 102-106.
WANG Y H, CHEN J P, ZHANG X, CHEN Y, CHEN D H. Effect of combination stress of temperature and humidity on amino acids and Bt protein content in leave of Bt cotton., 2012, 27(6): 102-106. (in Chinese)
[28] 陈德华, 聂安全, 杨长琴, 陈源, 吴云康. Bt棉毒蛋白表达特征与氮代谢关系及其化学调节的研究. 中国棉花, 2003, 30(7): 10-12.
CHEN D H, NIE A Q, YANG C Q, CHEN Y, WU Y K. The research of the relationship between Bt protein expression characteristics and nitrogen metabolism and its chemical control of Bt cotton., 2003, 30(7): 10-12. (in Chinese)
[29] DONG H Z, LI W J, TANG W, LI Z H, ZHANG D M. Heterosis in yield, endotoxin expression and some physiological parameters in Bt transgenic cotton., 2007, 126(2): 169-175.
[30] ZHANG X, LÜ C H, CHEN Y, WANG G X, CHEN Y, CHEN D H. Relationship between leaf C/N ratio and insecticidal protein expression in Bt cotton as affected by high temperature and N rate., 2014, 13(1): 82-88.
[31] Ian J R. Effect of genotype, edaphic, environmental conditions, and agronomic practices on Cry1Ac protein expression in transgenic cotton., 2006, 10(4): 252-262.
(责任编辑 杨鑫浩)
Relationship between Temperature, Soil Moisture, and Insecticidal Protein Content in Bt Cotton Boll Shell and the Mechanism of Nitrogen Metabolism
ZHANG Xiang, WANG Jian, PENG Sheng, RUI QiuZhi, LI LiNan, CHEN Yuan, CHEN Yuan, CHEN DeHua
(Yangzhou University/Jiangsu Provincial Key Laboratory of Crop Genetics and Physiology, Yangzhou 225009, Jiangsu)
【】 The effects of different temperature and soil moisture on the insecticidal protein contents in(Bt) cotton boll shell were investigated to provide a theoretical reference for the safe and stable utilization of Bt cotton in production.【】The study was undertaken on two Bt cotton cultivars Sikang 1 (SK1, a conventional cultivar) and Sikang 3 (SK3, a hybrid cultivar) during 2016 and 2017 growing seasons at Yangzhou University Farm, Yangzhou, China. The study was arranged with two factors that consisted of four temperature (29, 32, 35, and 38℃) and five soil moisture content (40%, 50%, 60%, 70% and 80% field capacity). The potted cotton plants were exposed to the twenty treatments for 4 days. In 2016, the effects of temperature and soil moisture on the insecticidal protein contents in boll shell were determined, and the soluble protein contents, glutamate oxaloacetate transaminase (GOT) activities, protease and peptidase activities were further studied in 2017. 【】The highest contents of insecticidal protein for SK1 and SK3 were both observed under the treatment of 32℃/60% field capacity, which was 471.1 ng·g-1FW and 351.7 ng·g-1FW, respectively. Under the same soil moisture, the insecticidal protein contents for SK1 and SK3 at 32℃ were significantly higher than those for other three temperature conditions. Under the same temperature, the insecticidal protein content under 60% field capacity for SK1 and SK3 was higher. Polynomial regression analysis showed a quadratic relationship between the insecticidal protein contents and the temperature and soil moisture. The binary quadric fitting equations for SK1 and SK3 were: Y=-3230.2+17.2X1+199.1X2-0.3X12-3.7X22-0.7X1X2(=0.829**), Y=-3322.0+40.7X1+145.2X2- 0.3X12-2.0X22-0.3X1X2(=0.739**), respectively, where Y stands for insecticidal protein contents, and X1is soil moisture, X2is temperature. Based on the equations, the maximum insecticidal protein contents would be obtained under the combination of 31.8℃/57.8% for SK1 and 33.2℃/60.8% for SK3, respectively. The physiological characteristics of nitrogen metabolism showed that the soluble protein contents and the GOT activities were highest in the boll shell for SK1 and SK3 under the combination of 32℃/60% field capacity, whereas the lowest protease and peptidase activities were also detected under this condition. The correlation analysis showed that there was a significant positive correlation between the insecticidal protein contents with soluble protein content and GOT activity (= 0.613**;= 0.735**), while the insecticidal protein was negatively correlated with protease activity and peptidase activity (= -0.724**;= -0.738**).【】 The temperature and soil moisture affected the expression of Bt insecticidal protein by regulating the protein synthesis and degradation. And the relationship between Bt protein contents and temperature and soil moisture could be quantified by a binary quadric fitting equation.
Bt cotton; temperature; soil moisture; insecticidal protein; nitrogen metabolism
2017-08-02;
2017-11-17
国家自然科学基金项目(31171479,31301263,31471435)、国家博士后基金项目(2016M591934)、江苏省博士后基金(1601116C)、江苏省高等学校自然科学研究重大项目(17KJA210003)、江苏省高校优势学科建设工程资助项目、江苏高校品牌专业建设工程、中国农业科学院科技创新工程项目(植棉技术标准化团队)(2016PCTS-1)、江苏省“双创博士”
张祥,E-mail:yzzhangxiang@163.com。
陈德华,Tel:0514-87979357;E-mail:cdh@yzu.edu.cn
猜你喜欢
河南科技学院学报(自然科学版)(2021年6期)2021-11-15 05:43:06
现代畜牧科技(2021年9期)2021-10-13 06:38:40
棉花学报(2021年1期)2021-02-05 09:37:16
今日农业(2019年12期)2019-08-15 00:56:32
中国棉花(2019年3期)2019-03-27 06:56:14
中国中医药信息杂志(2016年7期)2016-12-01 06:07:45
川北医学院学报(2015年5期)2015-12-05 08:22:28
天津医科大学学报(2015年3期)2015-06-05 12:21:49
新疆大学学报(自然科学版)(中英文)(2014年2期)2014-11-06 07:49:12
中国质量与标准导报(2014年4期)2014-03-11 19:54:48
