
美国红枫和元宝枫幼叶春季转色期生理特性研究
2018-04-16 05:58:40刘平宋岩陆秀君曹宝慧刘思佳王玉涛
生态环境学报 2018年3期
刘平,宋岩,陆秀君,曹宝慧,刘思佳,王玉涛
沈阳农业大学林学院,辽宁 沈阳 110866
彩叶树种叶色的变化是遗传因素和外界环境条件共同作用的结果(Wang et al.,2015)。大量研究表明,叶片呈色可能与光保护(Feild et al.,2001)、抗氧化(Kytridis et al.,2006)、渗透调节(Chalker-Scott,1999,2002)、与昆虫等协同进化(Archetti,2000)、光周期转变(Howe et al.,1995)等相关,也受低温(Close et al.,2002;Pietrini et al.,1998)、强光(姜卫兵等,2005)等因素影响。叶片呈色在幼叶、老叶、受胁迫的叶或主脉被切断的叶上都经常发生,很难用单一因素进行解释。相关研究表明,叶绿素、类胡萝卜素和花青素含量直接导致树叶的呈色效果。何亦昆等(1995)发现这3种色素在细胞中所占的位置和含量决定了叶片颜色。普通叶片中叶绿素含量比类胡萝卜素和花青素高,所以叶片呈绿色(潘瑞炽等,1995)。当叶片中花青素占绝对优势时(60%~80%),叶片呈现红色;当叶片中叶绿素和花青素比例减少到一定程度时(降到40%以下),叶片呈现出类胡萝卜素的黄色(楚爱香等,2013)。李玲(2013)通过对 5种红色叶植物紫叶小檗(Berberis thunbergii‘Atropurpurea’)、王族海棠(Malus ‘Royalty’)、紫叶桃(Prunus persica ‘Atropurpurea’)、紫叶李(Prunus ceraifera cv. Pissardii)、红枫(Acer palmatum cv. Atropuceum)的叶片色素进行分析,得出当花青素含量与叶绿素含量之比小于2时,花青素的红色主导作用减弱,绿色互补作用明显,叶片呈现紫红色;而当花色素苷含量/叶绿素含量值大于2时,叶绿素的互补效应弱,以花青素的红色为主导色,叶片呈现红色。有学者认为可溶性糖是花青素合成的能源物质和触发因子(Schaberg et al.,2003),糖在花青素的合成过程中可作为一种信号分子,激活促进花青素合成中一些酶的活性(Xavier et al.,2000)。糖类还可以调节有关植物代谢功能基因的表达,包括植物组织中花青素的表达(Dube et al.,1993)。
美国红枫(Acer×freemanii ‘Autumn Blaze’)又名红花槭,属槭树科槭树属,原产于北美洲,主要分布在加拿大和美国,是美国最受欢迎的绿化树种之一,是欧美经典的彩色行道树种,具有树姿优美、树冠整洁、叶型别致、叶色鲜艳等特点。近年被引入国内,成为我国重要的园林观赏树种。元宝枫(Acer truncatum)又名华北五角枫,属槭树科槭树属,树型优美、叶形奇特,秋季叶色艳丽,因季相变化丰富而广为人知,是华北地区园林中广泛应用的彩叶树种之一。
目前针对彩叶树种秋季叶片呈色期的生理研究较多,而春季幼叶变色的生理特征尚不明晰。为了揭示幼叶变色的原因,探索幼叶对周围环境所做的适应性变化,本文以美国红枫和元宝枫为研究对象,对其幼叶呈色期的色素含量、可溶性糖含量、叶片含水率、PSII原初光能转化效率(Fv/Fm)进行分析比较,以期掌握彩叶植物幼红的生理机制,为园林树种叶色改良或延长观赏期提供理论依据。
1 材料与方法
1.1 材料
试验所用美国红枫取自沈阳农业大学后山苗圃地,为6年生扦插苗,苗高2.1 m,地径为4.5~5 cm。元宝枫取自沈阳农业大学校园内,树高约为3.5 m,胸径约为15 cm。采样从4月21日开始,在晴朗的上午采集 5~10株树冠中部向阳处的枝条上的幼叶,然后分别装入黑色袋中带回实验室进行处理,采样间隔时间为4 d,共进行5次采样。
1.2 方法
质体色素和可溶性糖的提取与测定参照李合生(2000)的方法。花青素含量测定参照钱见平(2013)的方法。
叶绿素荧光采用德国进口便携式OP-30P叶绿素荧光仪进行测定:采样之前,在采样处选3~5片叶进行活体叶片测定,测定部位为叶正面,测定之前先对叶片进行暗处理15 min,然后用叶绿素荧光仪进行逐片测定。
叶片含水率采用烘干称重法测量:先用分析天平测量待测样本鲜重,然后用烘箱在 80 ℃下烘干叶片至恒重并测量干重,采用公式:叶片含水率=(鲜重-干重)/鲜重×100%进行计算。
光照强度采用便携式数字照度计 DCDS-3测量。从第1次采样前5 天开始,测量采样点处的光强。温度数据来源于国家气象数据网。
以上指标均每株3次重复。
1.3 统计分析
试验数据用 Excel处理并作图,用 SPSS 22.0进行数据分析。
2 结果与分析
2.1 春季转色期温度、光照强度与叶色变化情况
2.2 春季转色期叶片叶绿素和类胡萝卜素含量的变化
如表1所示,试验初期叶片呈现红绿色,后逐渐变为全绿色。由图1可知,试验期内两树种的质体色素变化趋势相似。其中,叶绿素a整体上都呈逐步上升趋势,并且含量远高于叶绿素b和类胡萝卜素含量。美国红枫叶绿素a变化范围为0.65~1.24 mg·g-1,试验后期比前期增加了 1倍左右,而元宝枫为0.39~1.24 mg·g-1,增加了2倍,上升速率明显快于美国红枫。美国红枫叶绿素b和类胡萝卜素变幅为 0.2~0.4 mg·g-1,小于元宝枫 0.1~0.4 mg·g-1的变幅。叶绿素b在中前期普遍低于类胡萝卜素,到了后期逐渐超过类胡萝卜素。叶绿素 a(F=0.01,P=0.932)与叶绿素 b(F=0.09,P=0.768)在树种间无显著差异,类胡萝卜素有差异但不显著(F=4.06,P=0.047)。这3种色素在转色期存在显著差异。由图2可知,两树种总叶绿素都逐渐增加,美国红枫从 0.88 mg·g-1逐渐增加到 1.63 mg·g-1,元宝枫从 0.5 mg·g-1增加到 1.67 mg·g-1,变色初期美国红枫总叶绿素含量普遍高于元宝枫,后期二者总叶绿素含量无显著差异。

表1 春季转色期环境因子与叶色变化情况Table 1 The changes of environmental factors and leaf color during leaf color transformation in spring
2.3 春季转色期叶片花青素含量的变化
试验期内叶片花青素含量的变化如图3所示。美国红枫叶片花青素含量从6.6色素单位逐步降低到4.4色素单位,降低33.3%。而元宝枫的变化趋势与美国红枫相反,在 2.4~4.1色素单位之间波动变化。整个试验期美国红枫都高于元宝枫。花青素含量树种间差异显著(F=68.69,P<0.001),两个树种各采样期之间也存在显著差异(F=19.35、12.98,P<0.001),说明花青素含量受树种和时间的影响很大。
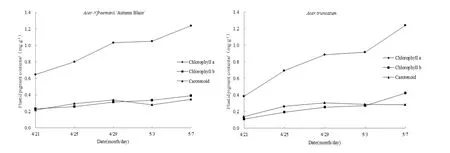
图1 春季转色期两树种质体色素含量变化Fig. 1 The changes of plastid pigment contents of two species during leaf color transformation in spring

图2 春季转色期两树种总叶绿素含量变化Fig. 2 The changes of total chlorophyll contents of two species during leaf color transformation in spring
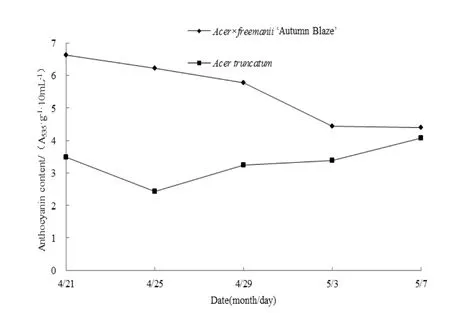
图3 春季转色期两树种花青素含量变化Fig. 3 The changes of anthocyanin contents of two species during leaf color transformation in spring
2.4 春季转色期叶片可溶性糖含量的变化
许多研究认为,可溶性糖的积累为彩叶植物叶片转色奠定了物质基础,且与花青素间存在一定的相关性。由图4可知,美国红枫和元宝枫可溶性糖都呈波动性变化,前者由初期52.0 mg·g-1降到26.3 mg·g-1,之后小幅增加到 30.4 mg·g-1;后者从初期到末期一直在34.5~44.3 mg·g-1范围内波动变化。可溶性糖在树种间差异显著(F=36.54,P<0.01),在不同采样期之间也存在显著差异(F=5.57,P=0.003;F=8.01,P<0.01),因此,可溶性糖含量也受到树种和时间的影响。

图4 春季转色期两树种可溶性糖含量变化Fig. 4 The changes of soluble sugar content of two species during leaf color transformation in spring
2.5 春季转色期叶片含水率的变化
如图5所示,试验期内两树种叶片含水率变化趋势相似,呈逐渐下降的趋势。美国红枫从68.39%逐渐降低到56.55%;元宝枫从74.64%逐渐降低到63.50%。转色后期下降速度明显加快,降幅分别为15.99%和11.95%。这是因为试验初期幼叶生命活动旺盛,抗逆性较弱,随着叶片的成熟,生命活动逐渐减弱,水分含量减少。整个试验期元宝枫叶片含水率高于美国红枫。
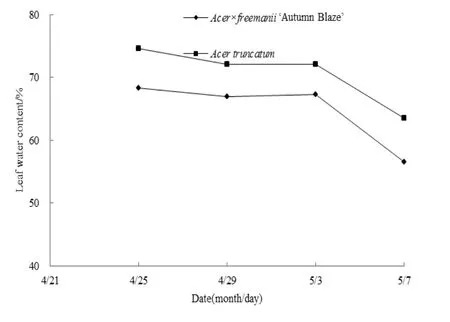
图5 春季转色期两树种叶片含水率变化Fig. 5 The changes of leaf water content of two species during leaf color transformation in spring
2.6 春季转色期叶片Fv/Fm的变化
Fv/Fm是 PSⅡ最大光化学量子产量,反映 PSⅡ反应中心内禀光能转换效率,也叫最大PSⅡ的光能转换效率,在非胁迫条件下,该参数的变化极小,不受物种和生长条件的影响;正常叶片的Fv/Fm在0.75~0.85之间,胁迫条件下该参数明显下降。因此,该指标可以反映树种受胁迫的程度。如图6所示,两树种Fv/Fm的变化规律相似,均随叶片成熟而逐渐升高。美国红枫最小值为0.53,最大值为0.72;元宝枫最小值为0.49,最大值为0.75。幼叶转色期也是叶片逐渐成熟的过程,从Fv/Fm变化来看,两树种幼叶呈现红色可能是一种抵御胁迫的重要途径。Fv/Fm在树种间差异不显著(F=0.114,P=0.736),而在不同采样期之间差异显著(F=34.58,P<0.01;F=38.63,P<0.01),表明美国红枫和元宝枫红色幼叶阶段可能受到了环境因子胁迫,随着叶片逐渐成熟,胁迫降低。

图6 春季转色期两树种叶片Fv/Fm变化Fig. 6 The changes of Fv/Fm of two species during leaf color transformation in spring
2.7 花青素与总叶绿素比值的变化
试验期两树种花青素与总叶绿素的比值(Ant/Chl)变化如图7所示。美国红枫从7.6逐渐降至2.7,而元宝枫从7.0迅速降低到2.7,之后变动不明显。树种间差异显著(F=24.10,P<0.01),不同采样期之间也存在显著差异(F=10.87,P<0.001;F=19.41,P<0.01),说明该参数受树种和时间的影响。这与所观察的叶色变化较一致,美国红枫随时间推进逐渐由红绿转为绿色,而且绿色越来越深,直至完全变绿,转色期在半个月左右;而元宝枫只在试验初期出现微红,后迅速变黄绿,直至完全变绿。从转色期色素含量变化可见,美国红枫和元宝枫幼叶呈现红色不是单一色素决定,而主要受Ant/Chl比值影响。当Ant/Chl大于2.5时,叶绿素的主导作用减弱,叶片呈现花青素的颜色——红色;当比值小于 2.5时,花青素的主导作用减弱,叶片呈现叶绿素的颜色——绿色。因此,认为色素比例影响着叶片颜色的变化。

图7 春季转色期两树种叶片花青素/总叶绿素的比值变化Fig. 7 The changes of Ant/Chl of two species during leaf color transformation in spring
2.8 生理指标的相关性分析
美国红枫和元宝枫幼叶各生理指标间的相关系数见表2。美国红枫叶绿素a、叶绿素b、类胡萝卜素和花青素之间都存在显著相关性。其中,叶绿素 a、叶绿素b、类胡萝卜素3个指标之间呈显著正相关,相关系数都很高。而花青素与这3个指标均呈负相关关系,其中,花青素与叶绿素a为极显著负相关,与叶绿素b为显著负相关,而与类胡萝卜素关系不显著。Fv/Fm值与各色素之间呈显著相关性,其中,Fv/Fm值与叶绿素a、叶绿素b、类胡萝卜素之间为显著正相关,而与花青素之间为显著负相关关系。其余生理指标间相关性不显著。元宝枫各生理指标间相关关系与美国红枫相似。叶绿素a、叶绿素b、类胡萝卜素和花青素之间也存在显著相关性。其中,叶绿素a、叶绿素b、类胡萝卜素3个指标之间均呈显著正相关(P<0.01),相关系数也很高。不同的是,其花青素与3个指标之间均呈极显著正相关关系。可溶性糖和叶片含水率与其他指标之间不存在显著相关性。
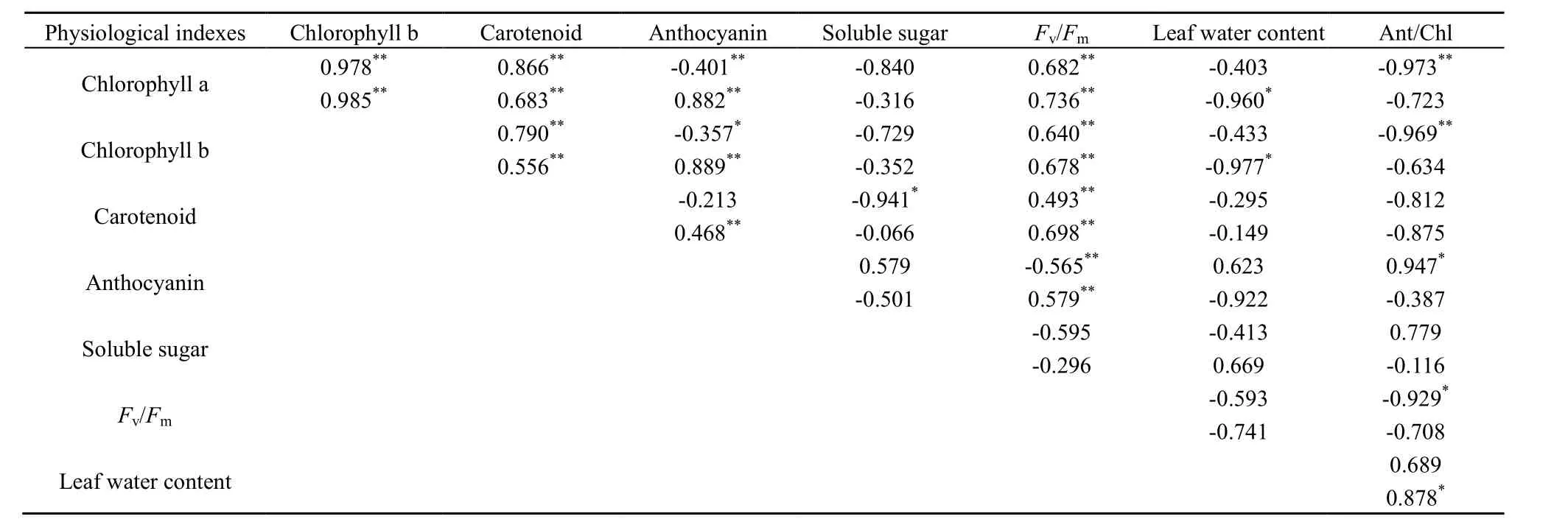
表2 春季转色期美国红枫和元宝枫各生理指标的相关性分析Table 2 The correlation analysis of different physiological indexes of Acer×freemanii ‘Autumn Blaze’ and Acer truncatum during leaf color transformation in spring
3 结论与讨论
3.1 结论
(1)美国红枫和元宝枫在春季幼叶由红转绿的过程中,叶绿素a、叶绿素b的含量都呈现逐步上升,叶绿素a+b增幅分别为85.3%和235.5%,类胡萝卜素没有显著变化。美国红枫花青素含量稳步下降,降幅为33.3%,而元宝枫呈现波动状态,且美国红枫在转色期花青素含量都高于元宝枫,树种间差异显著(P<0.01)。转色期美国红枫和元宝枫叶片可溶性糖含量呈波动性变化,树种间差异显著(P<0.01)。叶片含水率都呈缓慢下降趋势。Fv/Fm在树种间差异不显著,都随叶片成熟而逐渐增加,表明两树种叶片在逐渐成熟之前可能都受到不同程度胁迫。
(2)色素比例决定叶片颜色。当两树种的Ant/Chl比值大于2.5时,叶绿素的主导作用减弱,叶片呈现花青素的颜色——红色;当比值小于2.5时,花青素的主导作用减弱,叶片呈现叶绿素的颜色——绿色。
(3)相关性研究表明,两树种各色素之间都存在显著相关性,美国红枫花青素与质体色素表现为显著负相关,而元宝枫表现为显著正相关。两树种在花青素和可溶性糖之间都没有表现出显著相关性。Fv/Fm与各色素间存在显著相关性(P<0.01)。
3.2 讨论
3.2.1幼叶转色期色素变化
许多阔叶树种的幼叶在春季经常表现为红色(Taulavuori et al.,2011),随着树叶的生长发育,红色幼叶逐渐变为绿色成熟叶,绿色成熟叶在秋冬季随着树叶的衰老又变为红色。槭树属树种存在这样的变化特征。美国红枫和元宝枫幼叶都先呈现红色,随着叶子逐渐成熟完全变绿。在这个过程中,两个树种的叶绿素含量、类胡萝卜素含量、总叶绿素含量都逐渐增加,而花青素变化规律在两个树种间差异较大,美国红枫呈显著下降趋势,而元宝枫呈显著增加趋势,但是元宝枫花青素含量一直低于美国红枫。可见叶片呈色不是由单一色素决定的。相关研究表明,叶绿素、类胡萝卜素和花青素3种色素的比例大小影响叶片的颜色。据 Singh et al.(1988)研究,变叶木(Codiaeum variegatum)的斑斓叶色是光合色素(叶绿素 a、b及类胡萝卜素)和非光合色素(花青素)的比例变化的结果。不同品种红檵木(Loropetalum chinense var. rubrum)5月、6月是否出现返青现象,取决于花青素和叶绿素相对含量高低(唐前瑞等,2006)。陈继卫等(2010)认为日本红枫(Acer palmatum‘Atropurpureum’)转色期色素含量和比值变化快的叶片转色也快。朱书香等(2009)的研究表明,紫叶李(Prunus cerasifera var. arropurea)、紫叶矮樱(Prunus×cistenena ‘Pissardii’)、黑杆樱李(Prunus wrasifers‘Nigra’) 和 美 人 梅 ( Prunus×bliriana‘Meirenmei’)4种李属彩叶植物中各色素含量比例不尽相同且随着季节变化而变化,因此各树种呈现不同颜色。本研究结果也表明美国红枫和元宝枫在转色期如何呈色取决于花青素与叶绿素的比值。美国红枫幼叶由红变绿的转色期,花青素与叶绿素比值从7.6逐渐降低到2.7,试验后期叶色已完全变绿。而元宝枫花青素与叶绿素比值从 7.0迅速降低到2.5,之后变化趋于稳定,维持在2.5左右,同时观察到花青素与叶绿素比值在2.5左右时,叶色变为绿色。由此可知,当美国红枫和元宝枫花青素与总叶绿素比值大于2.5时,叶绿素的主导作用减弱,叶片呈现花青素的颜色——红色;当比值小于 2.5时,花青素的主导作用减弱,叶片呈现叶绿素的颜色——绿色。
3.2.2幼叶转色期各生理指标间的关系
叶片转色期,除了色素含量发生变化,其他一些生理指标也发生变化,如糖含量、酶活性、荧光特性、pH值、水分含量等。本研究结果显示,春季转色期美国红枫和元宝枫叶片可溶性糖呈波动性变化,树种间差异显著。两树种花青素与可溶性糖的相关性很小,没有达到显著水平。陈继卫等(2010)的研究结果也显示这两个指标相关性不显著,但李利霞(2015)的研究结论与此相反。可溶性糖的变化与光照强度和昼夜温差有很大关系,它的积累为彩叶植物叶片转色奠定了物质基础,且与花青素间存在一定的相关性(荣立苹等,2011)。陈建芳(2014)研究表明,叶面喷施不同浓度的蔗糖溶液均可促进元宝枫叶片中花青素相对含量的增加,其中以 4%的蔗糖溶液效果最佳。杨少华(2011)用 60 mmol·L-1蔗糖处理拟南芥(Arabidopsis thaliana)幼苗,促进花青素大量合成。因此,在实践管理中,采用有利于糖合成的栽培措施可以实现人工调控叶色,使叶片快速呈色,如采用叶面喷施蔗糖等措施。
美国红枫和元宝枫幼叶Fv/Fm较小,随着叶片逐渐成熟而增大到接近 0.78。Fv/Fm能够反映植物受胁迫的程度。Hughes et al.(2007)认为幼叶易受到高光胁迫而产生花青素使树叶变红,花青素在植物光敏感期扮演光保护的角色。而幼叶易受高光胁迫有很多原因,比如未成熟的叶绿体结构(Pettigrew et al.,1998;Choinski et al.,2003)、有限的气孔导度和 CO2交换量(John et al.,2009)等。Guo et al.(2000)认为春季低温胁迫不仅会损伤植物光合机构,而且会破坏光合电子传递和光合磷酸化以及暗反应的有关酶系,低温胁迫下植物更容易产生光抑制。美国红枫和元宝枫春季新萌生的幼叶普遍泛红,可能是因为该时期气温较低、叶片发育不完全,对低温还不太适应;并且此时光照强,幼叶的光合系统不完善,可能发生光抑制,因此需要合成大量的花青素来减轻光破坏。之后随着气温的升高和叶片的成熟,叶片更加适应外界环境,叶片光能转换效率提高,光合作用能力逐渐提高,因此受胁迫程度逐渐减小。本试验测量了两树种的 Fv/Fm,该参数的变化进一步说明春季幼叶受到一定环境因子胁迫,随着叶片的发育,胁迫程度逐渐减小。
本研究中发现叶片转色期 Ant/Chl与叶片的相对含水量呈正相关。Wang et al.(2014)的研究表明,幼叶未成熟表皮快速的蒸腾作用导致水分失衡,与树叶变红存在因果关系。陈卫元等(2007)发现红叶石楠(Photinia frasery)受到干旱胁迫时,叶片含水率减少,气孔导度也降低,而花青素含量大幅增加。目前有关幼叶呈现红色与叶片含水率的关系研究还较少,有待进一步研究。
参考文献:
ARCHETTI M. 2000. The origin of autumn colours by coevolution [J].Journal of Theoretical Biology, 205(4): 625-630.
CHALKER-SCOTT L. 1999. Environmental significance of anthocyanins in plant stress responses [J]. Photochemistry and Photobiology, 70(1):1-9.
CHALKER-SCOTT L. 2002. Do anthocyanins function as osmoregulators in leaf tissues? [J]. Advances in Botanical Research, 37: 103-127.
CHOINSKI J S, RALPH J R P, EAMUS D. 2003. Changes in photosynthesis during leaf expansion in Corymbia gummifera [J].Australian Journal of Botany, 51(1): 111-118.
CLOSE D C, BEADLE C L, HOLZ G K, et al. 2002. Effect of shadecloth tree shelters on cold-induced photoinhibition, foliar anthocyanin and growth of Eucalyptus globulus and E. nitens seedlings during establishment [J]. Australian Journal of Botany, 50(1): 15-20.
DUBE A, BHARTI S, LALORAYA M M. 1993. Inhibition of anthocyanin synthesis and phenylalanine ammonia-lyase activity by Co2+in leaf disks of Terminalia catappa [J]. Physiologia Plantarum, 88(2):237-242.
FEILD T S, LEE D W, HOLBROOK N M. 2001. Why leaves turn red in autumn. The role of anthocyanins in senescing leaves of red-osier dogwood [J]. Plant Physiology, 127(2): 566-574.
GUO Y P, ZHANG L C, HONG S S, et al. 2000. Responses of gas exchange and chlorophyll fluorescence to different low temperatures in satsuma mandarin [J]. Acta Photophysiologica Sinica, 26(2): 88-94.
HOWE G T, HACKETT W P, FURNIER G R, et al. 1995. Photoperiodic responses of a northern and southern ecotype of black cottonwood [J].Physiologia Plantarum, 93(4): 695-708.
HUGHES N M, MORLEY C B, SMITH W K. 2007. Coordination of anthocyanin decline and photosynthetic maturation in juvenile leaves of three deciduous tree species [J]. New Phytologist, 175(4): 675-685.
JOHN L, SINDER, JOHN S, et al. 2009. Juvenile Rhus glabra leaves have higher temperatures and lower gas exchange rates than mature leaves when compared in the field during periods of high irradiance [J].Journal of Plant Physiology, 166(7): 686-696.
KYTRIDIS V P, MANETAS Y. 2006. Mesophyll versus epidermal anthocyanins as potential in vivo antioxidants: evidence linking the putative antioxidant role to the proximity of oxy-radical source [J].Journal of Experimental Botany, 57(10): 2203-2210.
PETTIGREW W T, VAUGHN K C. 1998. Physiological, structural, and immunological characterization of leaf and chloroplast development in cotton [J]. Protoplasma, 202(1-2): 23-37.
PIETRINI F, MASSACCI A. 1998. Leaf anthocyanin content changes in Zea mays L. grown at low temperature: significance for the relationship between quantum yield of PS II and the apparent quantum yield of CO2assimilation [J]. Photosynthesis Research, 58(3): 213-219.
SCHABERG P G, VAN DEN BERG A K, MURAKAMI P F, et al. 2003.Factors influencing red expression in autumn foliage of sugar maple trees [J]. Tree Physiology, 23(5): 325-333.
SINGH S, SINGH S O. 1988. Photosynthetic and non-photosynthetic pigments in croton varieties [J]. Journal of the Andaman Science Association, 4(1): 77-78.
TAULAVUORI K, HUTTUNEN S, PIHLAJANIEMI H, et al. 2011.Reddish spring colouring of deciduous leaves: a sign of ecotype? [J].Trees, 25(2): 231-236.
WANG F, XING S J, JI Y P, et al. 2015. Leaf structural reddening in smoke tree and its significance [J]. Urban Forestry & Urban Greening, 14(1):80-88.
WANG F, YAMAMOTO H, Li X M, et al. 2014. Leaf reddening of sweet gum in water imbalance [J]. Ecological Informatics, 19(19): 47-52.
XAVIER V, FABIENNE L, STEPHANIE K, et al. 2000. Sugar sensing and Ca2+-calmodulin requirement in Vitis vinifera cells producing anthocyanins [J]. Phytochemistry, 53(6): 659-665.
陈继卫, 沈朝栋, 贾玉芳, 等. 2010, 红枫秋冬转色期叶色变化的生理特性[J]. 浙江大学学报:农业与生命科学版, 36(2): 181-186.
陈建芳. 2014. 温湿度和外源糖对元宝枫秋叶变色的影响研究[D]. 北京:北京林业大学: 43-45.
陈卫元, 曹晶, 姜卫兵. 2007. 干旱胁迫对红叶石楠叶片光合生理特性的影响[J]. 中国农学通报, 23(8): 217-220.
楚爱香, 张要战, 王萌萌. 2013. 四种槭树属(Acer)植物秋色叶变化与色素含量和可溶性糖的关系[J]. 江西农业大学学报, 35(1): 108-111,137.
何亦昆, 代庆阳, 苏学辉. 1995. 雁来红叶色转变与超微结构及色素含量的关系[J]. 四川师范学院学报: 自然科学版, 16(3): 195-197.
姜卫兵, 庄猛, 韩浩章, 等. 2005. 彩叶植物呈色机理及光合特性研究进展[J]. 园艺学报, 32(2): 352-258.
李合生. 2000. 植物生理生化实验原理与技术[M]. 北京: 高等教育出版社: 134-137, 194-197.
李利霞. 2015. 鸡爪槭叶色变化机制的研究[D]. 重庆: 重庆师范大学.
李玲. 2013. 五种红色叶植物叶片色素分析[D]. 西安: 西北农林科技大学: 38-39.
潘瑞炽, 董愚得. 1995. 植物生理学[M]. 第 3版. 北京: 高等教育出版社: 77-79.
钱见平. 2013. 元宝枫转色机理的初步研究[D]. 泰安: 山东农业大学:11-12.
荣立苹, 李倩中, 李淑顺, 等. 2011. 三角枫及其变异株转色期叶色变化生理[J]. 江苏农业学报, 27(5): 1089-1092.
唐前瑞, 陈德富, 陈友云, 等. 2006. 红檵木叶色变化的生理生化研究[J]. 林业科学, 42(2):111-115.
杨少华. 2011. 蔗糖调节拟南芥花青素的生物合成[J]. 中国生物化学与分子生物学报, 27(4): 364-369.
朱书香, 杨建民, 王中华, 等. 2009. 4种李属彩叶植物色素含量与叶色参数的关系[J]. 西北植物学报, 29(8):1663-1669.
猜你喜欢
故事作文·低年级(2024年3期)2024-04-01 01:35:05
山东农业科学(2022年10期)2022-11-17 06:28:02
中国生物防治学报(2021年1期)2021-04-18 03:26:07
食用菌(2020年1期)2020-02-18 08:17:22
中国农资(2019年50期)2020-01-02 08:52:54
课外生活(小学1-3年级)(2019年8期)2019-08-27 03:58:22
山东林业科技(2019年2期)2019-06-03 10:10:56
吉林农业(2019年9期)2019-01-06 06:06:24
今日农业(2019年14期)2019-01-04 08:57:40
现代园艺(2017年21期)2018-01-03 06:41:57
