
大黄鱼人工感染杀香鱼假单胞菌的病理形态学及病原分布的初步研究
2018-04-11 09:00谢建军王庚申许文军
浙江海洋大学学报(自然科学版) 2018年6期
陈 卓,施 慧,谢建军,汪 玮,王庚申,何 杰,许文军
(浙江海洋大学海洋与渔业研究所,浙江省海洋水产研究所,浙江省海水增养殖重点实验室,浙江舟山 316021)
杀香鱼假单胞菌Pseudomonas plecoglossicida是引起大黄鱼Pseudosciaena crocea内脏白点病的病原,大黄鱼一旦感染发病,在自然状态下死亡率较高,是危害大黄鱼养殖业的重要病原菌。本实验室在养殖生产中发现杀香鱼假单胞菌还会感染黑鲷Acanthopagrus schlegelii、小黄鱼Larimichthys polyactis等其它海水养殖鱼类。研究表明杀香鱼假单胞菌对多种水产常用抗生素类药物有抵抗性,通过依赖药物以控制该菌效果较差,也可能导致水产品中的药物残留进而引起食品质量安全问题[1-4]。但是,近年随着养殖环境的日益恶化,大黄鱼内脏白点病在各大黄鱼养殖区域广泛爆发,给大黄鱼养殖业带来了巨大的经济损失。
目前,已有不少学者对杀香鱼假单胞菌的致病机理及相关的毒力因子展开研究,张丹枫等[5]对大黄鱼进行了人工感染实验与超微电镜的观察,从病理学角度展示了病原菌对鱼内脏组织的入侵与危害;张杰等[6]采用双交换同源重组法构建了杀香鱼假单胞菌Ⅲ型分泌系统转录调控蛋白ExsA突变株,从分子生物学角度阐述了该菌的致病机制,但杀香鱼假单胞菌的致病机理至今尚不清楚。本文拟通过人工感染试验分析杀香鱼假单胞菌在大黄鱼各器官组织中的分布情况及其对组织产生的病理学变化来阐述病理损伤与病原分布的相互关系,为更深入的探讨杀香鱼假单胞菌致病机理以及疾病的防治提供理论基础和依据。
1 材料与方法
1.1 材料
杀香鱼假单胞菌ZS161211为本实验室分离和保存;健康大黄鱼体重约40±3 g,购自舟山台门星鑫水产养殖场,对试验鱼进行细菌分离及寄生虫观察确认无病后实验室暂养1周备用;兔抗杀香鱼假单胞菌多克隆抗体由本实验室制备保存。
硫代硫酸盐柠檬酸盐胆盐蔗糖琼脂培养基(TCBS)、胰蛋白胨大豆肉汤培养基(TSB),购自杭州微生物试剂有限公司;电泳仪、紫外凝胶成像仪(Bio-Rad);离心机(Eppendorf);恒温培养箱(上海精宏实业有限公司);恒温摇床(太仓市科教器材厂);显微镜(Nikon)。
1.2 试验方法
1.2.1 人工回感试验
将保存的菌株ZS161211在TSA培养基上划线,18℃下培养24~48 h,用1.5%生理盐水从平板上洗下菌苔,分光光度计测定菌液浓度,将菌液稀释至终浓度为5×104 cfu/mL。试验鱼每尾腹腔注射稀释的菌液100 μL,使感染菌相对鱼体重的终浓度为100 cfu/g,对照组鱼腹腔注射1.5%生理盐水100 μL。试验期间水温控制在20℃,不投饵,连续充气。注射后24 h开始取样,取样时对试验鱼的肝脾肾内脏组织分别进行细菌分离,同时取适量组织块使用Bouin氏液进行固定。
1.2.2 人工感染试验鱼病理组织切片制备
无菌取发病鱼的肝、脾、肾等内脏组织,加1.5%无菌生理盐水剪碎,涂布TCBS培养基,27℃培养24~48 h观察细菌生长情况;取试验鱼的肝、脾、头肾、中肾及鳃丝分别约4 mm×4 mm×5 mm,经Bouin氏液固定,在24 h后换数次75%乙醇冲洗,最后保存于75%的乙醇中。固定的样品经后续不同浓度乙醇系列脱水及丙酮脱水过程后,移入二甲苯溶液中透明,经浸蜡包埋后切片、贴片,烤干后展片以苏木精-伊红染色法(H&E染色)染色,树胶封片,显微镜下观察拍照。
1.2.3 免疫组化检测杀香鱼假单胞菌在组织内分布
将已制作好的石蜡切片脱蜡水化,滴加3%H2O2于载玻片上进行内源性酶灭活,然后将切片浸入枸橼酸盐缓冲液中,直接煮至92~98℃进行抗原暴露,冷却后滴加5%BSA封闭,滴加兔抗杀香鱼假单胞菌抗湿盒孵育,滴加生物素化山羊抗兔二抗湿盒孵育后,加入辣根过氧化物酶进行SP反应,滴加新鲜配制的试剂DAB显色,清洗后苏木素复染,最后滴加中性树脂胶封片。用PBS代替一抗作为阴性对照,显微镜下观察拍照,显微镜下中呈棕褐色反应产物代表抗原的定位,未有抗原定位的则被苏木精染色成蓝色。
2 结果
2.1 临床症状与剖解变化
试验组鱼注射24 h后出现游动缓慢,应激反应迟钝等临床症状;注射72 h后试验鱼出现死亡,死亡鱼体表无溃疡等症状,主要是鳃充血。剖解病死鱼,部分鱼有腹水,肝充血严重,脾脏布满白色小结节和肾脏白色结节较脾脏大(图1)。无菌取病死鱼的内脏组织分别在TCBS和TSA培养基上接种划线,27℃培养24 h,TCBS上长出特征性的杀香鱼假单胞菌针尖大小的绿色小菌落,TSA上为乳白色、边缘光滑、表面湿润的菌落。经药敏试验和生理生化学试验确认与感染菌株ZS161211一致。
临床解剖观察(图2)显示试验组感染鱼的肝、脾、肾等内脏组织与器官都表现出不同程度的充血、出血、坏死等。肝细胞发生广泛性变性,或坏死、溶解,有大量的炎性细胞浸润;肾小管上皮细胞肿胀变性,肾间质巨噬细胞和中性粒细胞浸润、严重空泡变性、坏死崩解;脾脏少量含铁血黄素的沉积,淋巴细胞坏死,中性粒细胞和大量巨噬细胞浸润。
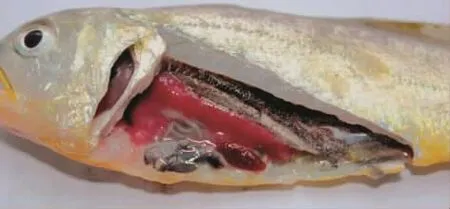
图1 病死鱼临床解剖图Fig.1 Clinical anatomic map of P.crocea

图2 病鱼脾脏充满白色结节Fig.2 P.crocea spleen full of the white nodulars
2.2 组织病理学变化
健康大黄鱼肝组织结构清晰,细胞形状规则;感染鱼肝脏组织结构不明晰且伴随充血,肝血窦与中央静脉瘀血更明显,肝血窦、中央静脉以及一些肝细胞内发现病原菌;肝细胞肿胀明显、细胞核固缩,病变严重的区域失去原有形态结构形成坏死灶,坏死灶中心区内充满解体后的细胞碎片(图3)。
健康脾脏结构清晰,细胞完整;感染鱼脾脏组织坏死严重,实质结构遭到破坏,在红髓和白髓均可见散在的坏死灶,坏死灶内大量中性粒细胞和巨噬细胞浸润,脾髓出血较为严重,有少量含铁血黄素的沉积,坏死灶内实质细胞界限模糊不清,崩解后呈现空泡化。
健康肾脏组织致密;感染鱼肾脏充血,结构紊乱;肾小管形状不规则,上皮细胞肥大、颗粒变性且界限模糊不清晰、管腔狭窄,部分上皮细胞增生堵塞官腔;肾小球萎缩或崩解;严重的肾间质组织广泛性坏死,大量巨噬细胞和中性粒细胞浸润,间质细胞发生玻璃样变性,严重的坏死,甚至溶解、消失。
健康鳃丝组织紧密,有规律性;感染鱼鳃丝排列不整齐,有的呈现波纹状扭曲;鳃小片肿大,窦状隙扩张充血、渗出;鳃小片上皮细胞肥大、变圆,严重的上皮细胞崩解坏死脱落。
2.3 菌体在鱼体内的分布
阳性信号主要分布于肝窦、巨噬细胞、肾小管上皮细胞、肾间质细胞和巨噬细胞内。肝组织强阳性信号主要集中于肝窦和静脉,吞噬细胞破裂后部分周边区域、肝窦周围少量肝细胞和窦壁细胞有明显坏死性病变;脾组织中的阳性信号位于脾脏间质细胞和巨噬细胞的白髓区。这些迅速增殖的感染性病原体在器官和组织中引起明显的病理变化。肾组织中的阳性信号主要位于肾小管上皮细胞和肾小球淋巴细胞。由于在肾间质中大量的细菌复制,组织发生坏死和崩解,并且也可在崩解组织中检测到。鳃组织中的阳性信号集中在小的上皮细胞和基底毛细血管中(图4)。

图3 人工回感鱼组织病理变化图,HEFig.3 Histopathological lesions on P.crocea infected by P.plecoglossicida

图4 人工回感鱼组织免疫组化图Fig.4 Immunohistochemical staining results of P.crocea infected by P.plecoglossicida
3 讨论
本研究在水温20℃条件下,对养殖大黄鱼进行了杀香鱼假单胞菌人工感染实验,运用组织病理与免疫组化技术对发病鱼内脏组织进行分析,结果显示杀香鱼假单胞菌对大黄鱼组织机体,尤其是内脏组织造成了严重损伤。
大黄鱼内脏白点病是近年来在大黄鱼养殖中暴发流行的一种细菌性疾病,因病鱼脾、肝脏、肾等内脏长满白色结节而得名,假单胞菌属的细菌被确认为该病病原,目前有许多关于大黄鱼内脏白点病病原的报道[7-9]。由于大黄鱼内脏白点病临床发病急、体表无明显临床症状、低水温发病等特点给该病的防治造成很大困难。关于该病病原假单胞菌种的鉴定一直未能统一,直到2013年张杰等[6]通过全基因组测序确认其为杀香鱼假单胞菌。杀香鱼假单胞菌由日本学者NISHIMORI,et al[10]从淡水养殖的患病香鱼体内首次分离得到并且命名。杀香鱼假单胞菌不仅存在于淡水环境中,也广泛存在于海水环境中,而且是危害较大的致病菌[4]。
本文采用杀香鱼假单胞菌单位为100 cfu/g,注射至大黄鱼腹腔内,感染24 h即开始发病,肝、脾、中肾、头肾上出现明显的白色结节,与自然发病症状相同,且从感染鱼内脏组织中分离培养到了杀香鱼假单胞菌。组织病理观察结果显示,感染鱼肝、脾、肾等内脏组织都出现明显病变损伤,全身组织器官广泛性表现出不同程度的充血、出血、坏死等,随着菌体在大黄鱼体内大量增殖,最终导致其全身多组织细胞坏死溶解,鱼因多组织器官的功能衰竭而死亡,这些组织病理变化与张丹枫等[5]对自然患病鱼的组织病理观察结果一致。
免疫组化技术利用标记的抗体对细胞或组织内的相应抗原进行定位检测,经过组织化学的显色反应呈现醒目的阳性色彩,因此免疫组化可清楚直观的反映病原感染生物体后的分布状况[11-12]。随着现代鱼类病害学、分子生物学的迅速发展,使病理形态和功能的研究更加深入,免疫组化已成为在水产病害学科研究中的新型手段。连林坤[13]运用免疫组化方法开展了螺原体对中华绒螯蟹Eriocheir sinensis H.Milne-Edwards和克氏原螯虾Procambarus clarkii不同器官和组织的感染病理学研究,证实了不同水生动物上分离到的螺原体对不同寄主存在感染差异。本试验应用免疫组化法对人工感染大黄鱼各组织器官内杀香鱼假单胞菌进行定位检测,在试验组鱼的肝、脾、肾和鳃丝各相关组织器官中观察到了特征性的阳性信号,这些阳性信号主要分布于肝血窦、脾间质或巨噬细胞、肾间质活巨噬细胞以及腮丝的巨噬细胞内,表明杀香鱼假单胞菌对机体多组织、器官都具有侵染性。试验结果表明免疫组化能成功反映杀香鱼假单胞菌感染大黄鱼后在机体内的分布情况。通过组织病理学观察与免疫组化染色的结果相比较,鱼体组织病理损伤程度与免疫组化检测的中阳性信号的有无与强弱表现出较高一致性,其病理损伤与病原菌分布数量呈明显的正相关。
鱼类的巨噬细胞是鱼类的非特异性免疫系统的重要组成部分,当病原微生物表面覆盖有免疫球蛋白和补体成分时,巨噬细胞可通过这些因子的特异性受体识别并杀伤微生物[14]。但一些病原菌可以适应巨噬细胞内的生活环境,并作为其生活史的一部分,使其自身免受其它细胞介导和体液免疫反应的伤害,巨噬细胞内存活能力已成为很多病原菌毒力机制的重要组成部分[15-16]。张杰等[17]通过电镜超薄切片观察发现杀香鱼假单胞菌也可以在巨噬细胞内存活和增殖,本试验的免疫组化结果显示在试验鱼的多个组织器官的巨噬细胞内出现了强阳性信号,为以后进一步进行杀香鱼假单胞菌对大黄鱼的致病机理研究提供了更多参考资料。
猜你喜欢
海洋开发与管理(2022年10期)2022-11-11
辽河(2022年5期)2022-07-07
中国临床医学影像杂志(2022年2期)2022-05-25
当代水产(2022年3期)2022-04-26
农产品市场周刊(2021年17期)2021-09-28
感染、炎症、修复(2021年1期)2021-07-28
中国现代医药杂志(2020年10期)2020-12-14
散文诗世界(2019年6期)2019-09-10
中国临床医学影像杂志(2019年6期)2019-08-27
意林·全彩Color(2019年7期)2019-08-13
