
美国《Emerging Infectious Diseases》2017年第12期有关人兽共患病论文摘译
2018-04-09 01:08:19何梓凯,林丹
中国人兽共患病学报 2018年2期
P1958不发酵山梨醇的产志贺毒素大肠埃希菌的进化//Kyle Schutz, Lauren A. Cowley, Sharif Shaaban,等
2014年7月在英格兰暴发了一起由产志贺毒素大肠埃希菌(以下简称STEC)引起的疫情,31人患病,其中13人(42%)患了溶血性尿毒综合征。研究人员对病菌进行分离和测序,并把测序的结果与已知的大肠埃希菌O55:H7以及O157:H7的结果进行了比较。根据STEC O55:H7暴发菌株的主要基因组进行系统发育分析,表明其进化史的最简约模式为,能发酵山梨醇的致病性祖代O55:H7菌株,被编码志贺毒素2a(stx2a)的噬菌体溶原化,引起srlA基因无义突变而失去发酵山梨醇的能力。STEC O157:H7和STEC O55:H7平行而聚敛的进化史可能表明了二者进化过程中存在共同的驱动因素。作为有临床意义的病原菌,STEC O157:H7的出现与获得编码stx2a噬菌体有关,因此带有stx2a基因的STEC O55:H7的出现,是公共健康领域当前所关注的一个问题(图1)。
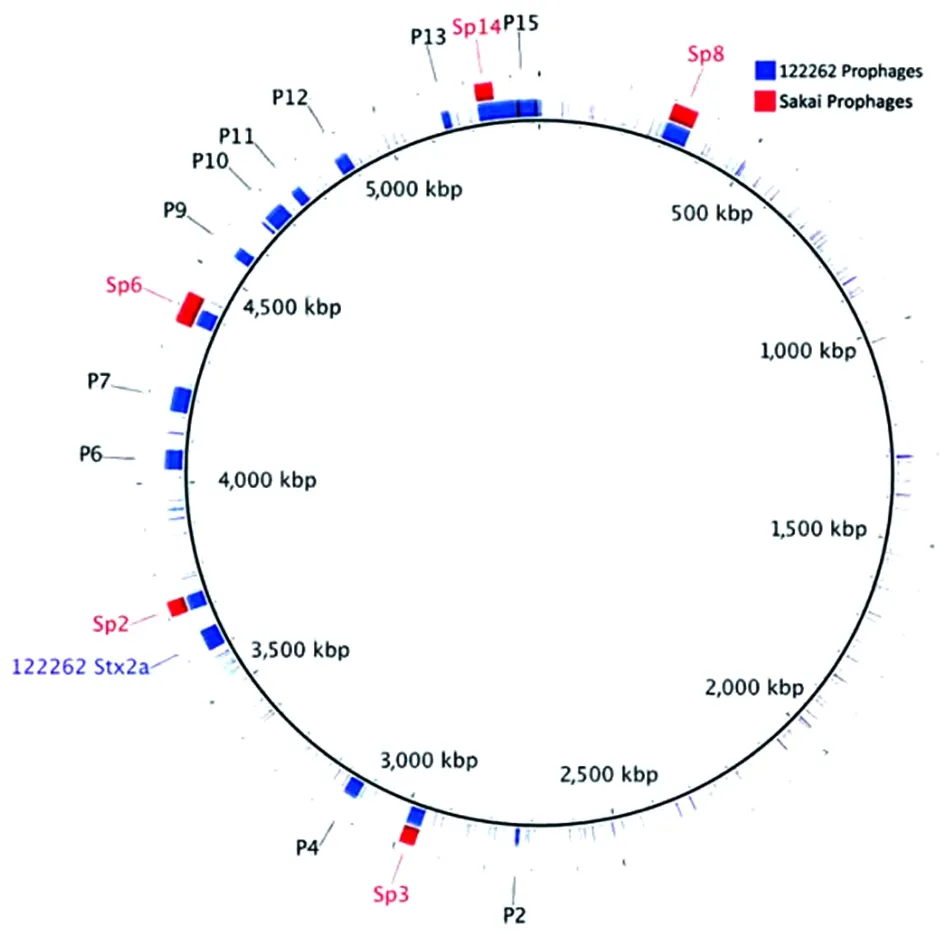
BLAST环形图像生成器(BRIG)绘制的环状图。采用BLAST+(21)程序,比较了STEC O55:H7 122262的前噬菌体和同源的STEC O157:H7 Sakai前噬菌体。STEC O55:H7 122262 染色体被设置为参比基因组,122262前噬菌体(P1-P15)组成第一环。同源的STEC O157:H7 Sakai噬菌体(Sp2,Sp3,Sp6,Sp8和Sp14)在通过BLAST分析识别后,根据在表2中的已知位置添加到图像中。推测的前噬菌体序列数据是从PHAge搜集工具(22)中检索得到,并绘制进BRIG图中。
P1966发生多次重配的病毒是2016年荷兰甲型H5N8高致病性禽流感流行的原因//Nancy Beerens, Rene Heutink, Saskia A. Bergervoet,等
2016年,在荷兰暴发了由高致病性甲型H5N8禽流感病毒引起的禽间疫情,导致了大量野鸟死亡,部分商业养殖场和鸟类饲养基地也受到了波及。为了了解野鸟和家禽所携带的病毒之间的关系,研究人员对提取到的病毒进行全基因组测序。系统进化分析的结果表明,该型病毒与2016年5月在俄罗斯发现的clade2.3.4.4分支的H5亚型病毒有关,但又包含了新型PB2和NP基因片段,以及2种不同的PA片段的变异。分子时钟分析表明,基因重配最可能发生在俄罗斯或蒙古的野鸟体内。此外,荷兰野鸟中还发现2株遗传背景不同的H5N5重配病毒。此项研究为clade 2.3.4.4的H5亚型病毒的快速和持续重配提供了依据,并可能导致致病性、传染性、传播能力和人兽共患能力等病毒特征的快速变化(图2)。
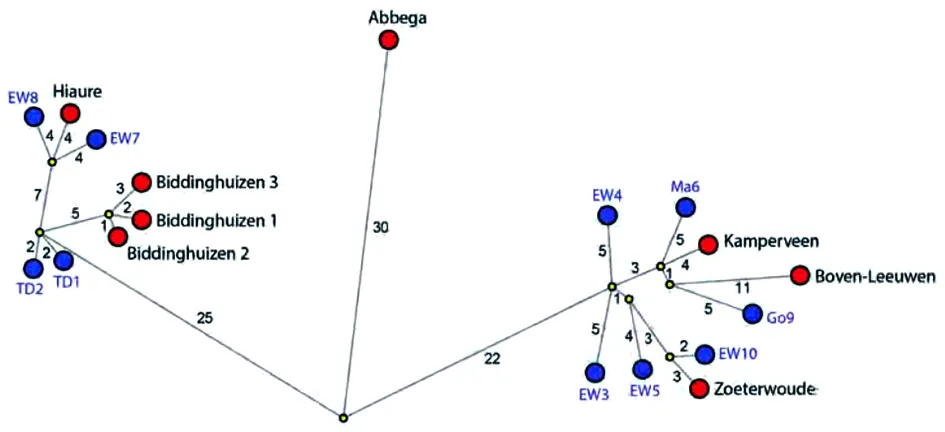
图2中介邻接网络图显示2016年在荷兰商业家禽养殖场(红点)分离得到的高致病性甲型H5N8禽流感病毒与最相似的野鸟病毒(蓝点)之间的遗传关系。预测的中值向量用黄色表示。图中线的长度表示遗传距离,数字表明了核苷酸变化数目。这里分析了编号1-10的野鸟病毒分离株。(见在线技术附件1表2,地址:https://wwwnc.cdc.gov/EID/article/23/12/17-1062-Techapp1.xlsx)GISAID EpiFlu数据库(地址:https://www.gisaid.org)登录号见在线技术附件1表1中。EW,欧亚赤颈鸭;Go,灰鹅;Ma,绿头鸭;TD,凤头鸭
P1974一种新鉴定的正粘病毒——韦尔弗利特湾病毒对欧绒鸭雏的实验感染//Valerie Shearn-Bochsler, Hon Sang Ip, Anne Ballmann,等
韦尔弗利特湾病毒(以下简称WFBV)是一种Quaranjavirus属的新型正粘病毒。2006年在韦尔弗利特湾(地处美国马萨诸塞州巴恩斯特布尔郡)的一次欧绒鸭集体死亡事件中,首次从鸭子尸体上分离得到该病毒,并且在后来该地区反复多次发生的欧绒鸭死亡事件中分离得到。在欧绒鸭的尸体中,美国的研究人员发现鸭子的肝脏、胰脏、脾脏和肠道都有坏死的现象。因此他们对6周龄的健康欧绒鸭进行WFBV病毒的接种,以期重现疾病发生的自然过程。大约25%的受接种幼鸭产生了疾病的临床症状,并需要被执行安乐死;在试验中,另外有18.75%的幼鸭患病,体重下降。对照组的幼鸭未被病毒感染,不发生临床症状。实验组中出现临床症状的幼鸭与那些自然感染并死亡的幼鸭出现一致的病理损害。研究表明,从37.5%受感染的鸭子身上分离得到WFBV病毒,存活至接种后5日的幼鸭产生了针对WFBV的血清抗体滴度(图3)。
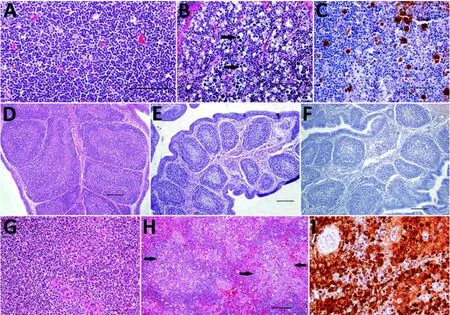
图3对照组和实验组实验感染韦尔弗利特湾病毒的欧绒鸭)淋巴器官的组织病理学和免疫组织化学法(IHC)法检查(接种2天后)。A图一只对照组小鸭的胸腺皮质组织的苏木精和伊红(H&E)染色;B图,一只实验组鸭子的胸腺组织H&E染色,箭头处显示淋巴细胞出现多病灶性急性凋亡;C图,一只实验组鸭子的胸腺组织的IHC染色,显示胸腺细胞上对WFBV病毒的阳性免疫标记;D图,一只对照组鸭子法氏囊的H&E染色;E图,一只实验组鸭子法氏囊的H&E染色,显示淋巴细胞有显著的弥散性消耗;F图,一只实验组鸭子法氏囊的IHC法染色,显示其对WFBV病毒呈阴性免疫标记;G图,一只对照组鸭子的脾脏组织的H&E染色;H图,一只实验组鸭子脾脏组织的H&E染色,箭头处显示为多病灶合并急性坏死;I图,脾脏组织IHC染色,放大200倍,显示其对WFBV病毒有阳性免疫标记。在图A,B和C中,比例尺为50μm,在图D,E,F,G和H中,比例尺为100μm。
P1999匈牙利神经侵袭性星状病毒暴发导致仔猪脑脊髓炎、虚弱和瘫痪//ákos Boros, Mihály Albert, Péter Pankovics,等
近2年来在匈牙利一座规模大、产量高的养猪场里,新生猪幼崽(25-35日龄)中流行一种神经系统疾病临床表现为后半身麻痹、高死亡率,尸检发现存在脑脊髓炎和神经系坏死。从该养猪场6份猪脑和脊髓标本,通过RT-PCR和原位杂交法,鉴定出了猪星状病毒3型。通过定量RT-PCR法在标本组织测试中发现,脑干和脊髓中病毒载量最高。从另外2个地理位置相距遥远的养猪场中留存的动物脑和脊髓标本中,也发现了类似的猪星状病毒3型。病毒RNA主要集中在神经细胞特别是脑干、小脑(浦肯野细胞)、颈部脊髓的神经细胞中。星状病毒基本上无法在粪便中被检测到,但出现在呼吸道的样本中,这表明星状病毒存在通过呼吸道感染的可能,因此星状病毒可导致常见的造成神经系统障碍的传染病(图4)。

图4
匈牙利某农场2头有病症新断奶猪崽的中枢神经系统组织的病理学检查结果。颈髓(A-E)、小脑(F-J)和脑皮层(M)的病理切片是来自于指征病例动物(GD-1),脑干(K)的病理切片来自于另外一只处于I期发病的动物(GD-11)。图(A,D,F,I,L)用苏木精和伊红染色。脑灰质(图A,D)和分子层(图F,I,L和M)多病灶出现胶质细胞增生(黑色箭头)。神经元变性和坏死明显,表现为嗜酸性粒细胞增多、角变性、神经元结构缺失和出现空泡(图A, D中双箭头)。一些浦肯野细胞神经元由于轻度空泡化而出现细微的角变(图I中双箭头)。原位杂交法检测神经侵袭性猪星状病毒。猪星状病毒杂交探针局限于神经元(图B,E,K的白色箭头),或者浦肯野细胞神经元(图G,J黑色双箭头)并延伸到穿过分子层的树突(图G,J黑色箭头)。胶质增生细胞(图L、M中黑色箭头)也出现猪侵袭性星状病毒3型的杂交探针。图C、H为对照探针在连续切片上,没有杂交信号。原位杂交中,(图A-C,F-H)的比例尺为500μm,图(D,E,I-M)为50 μm。
(福建省疾病预防控制中心何梓凯、林丹 译陈爱平 校)
猜你喜欢
昆明医科大学学报(2022年2期)2022-03-29 00:51:20
世界科学技术-中医药现代化(2021年7期)2021-11-04 08:10:52
考试与评价·高一版(2021年2期)2021-08-14 23:53:10
中国动物传染病学报(2021年3期)2021-07-21 03:19:28
求学·理科版(2020年7期)2020-07-14 08:27:44
参花·青春文学(2020年3期)2020-03-18 16:37:54
森林与人类(2016年2期)2016-03-07 11:54:12
广东海洋大学学报(2015年3期)2015-12-22 10:05:26
中国医学影像学杂志(2015年9期)2015-12-15 11:03:28
沈阳医学院学报(2014年4期)2014-12-27 13:44:32
