
油用牡丹凤丹光合特性
2018-04-09 05:47张志浩杨玉珍
江苏农业科学 2018年5期
张志浩, 杨玉珍, 李 娟, 潘 婷
(1.郑州大学生命科学学院,河南郑州 450000; 2.郑州师范学院生命科学学院,河南郑州 450044)
牡丹(PaeoniasuffruticosaAndr.)原产于中国,为芍药科芍药属多年生木本落叶小灌木,在我国已有1 600多年的栽培历史[1],素有“国色天香”“花中之王”的美称。通常将牡丹分为观赏牡丹和药用牡丹两大类,前者以洛阳和菏泽地区种植的品种为代表,后者以凤丹(P.ostii)和紫斑(P.rockii)最为著名[2]。近年来,随着研究不断深入,凤丹和紫斑除作传统药用和观赏栽培外,由于其籽营养价值高,还可作为优质木本粮油资源进行开发,受到人们的广泛关注[3]。河南作为农业和人口大省,依托其地理优势和技术优势,在河南省推广栽培油用牡丹不仅能解决农村劳动力,增加农民收入,同时能够促进粮油生产,保障国家粮油安全,具有重要的战略意义。为进一步提高其观赏和药用价值、开发其油用价值,许多学者对凤丹的育苗与栽培技术[4]、组织培养[5-6]、分子育种[7]、药用成分[8]及籽油成分[9]等进行了研究,但是对其光合特性的研究鲜有报道。毕玉伟等虽以盆栽凤丹为试材,初步探究了不同季节的光合特性[10],但缺乏田间条件的对照;张衷华等虽分析了2种主要油用牡丹品种的光合特性及其微环境因子,但仅局限于2个品种的原始集中产区安徽铜陵和甘肃兰州[11]。凤丹为安徽四大名贵中药植物之一,属江南品种,原生长地与河南地区气候差异明显,因此有必要对其在河南地区田间条件下的光合特性进行深入研究。本研究以油用牡丹主要优良栽培品种凤丹为研究对象,在田间条件下对其光合特性进行探讨,比较不同季节的光合日进程,探究其对不同季节气候因子的适应性,为油用牡丹在河南地区的良种选育、大面积推广栽培以及生产管理提供理论指导和科学依据。
1 材料与方法
1.1 材料
供试材料为5年生凤丹白品种(P.ostii‘Phoenix White’),大田种植,水肥条件良好,常规管理。周边无高大遮挡物,处全光照环境。随机选取长势基本一致,无病虫害的健康植株,顶端往下第3或第4张向阳功能叶,尽量保持叶片正常生长角度不变。种植于郑州师范学院黄河滩实习实训基地,该地位于郑州市北郊(34°51′N、113°38′E,海拔110 m),属北温带大陆性季风气候,冷暖适中、四季分明,春季干旱少雨,夏季炎热多雨,秋季晴朗日照长,冬季寒冷少雪,年平均气温在14.0~14.3 ℃之间,平均降水量640.9 mm,无霜期 220 d,全年日照时间约2 400 h。
1.2 方法
于2016年5月上旬(春季)和6月下旬(夏季)选择连续晴朗天气下对凤丹进行光合日变化测定。运用LI-6400XT便携式光合测量系统(LI-COR,Inc,USA)测定:净光合速率(photosynthetic rate,Pn)、气孔导度(conductance to H2O,Gs)、胞间CO2浓度(intercellular CO2concentration,Ci)、蒸腾速率(transpiration rate,Tr)并同时测定大气温度(temperature of air,Tair)、叶片温度(temperature of leaf,Tleaf)、大气CO2浓度(concentration of CO2in air,Ca)、光合有效辐射(in-chamber quantum sensor,PARi)、空气相对湿度(relative humidity in air,RH)、水汽压亏缺(vapor pressure deficit,VPD)等生态因子。测定时间为08:00—18:00,隔1 h测定1次,每次随机选择 3~5个典型株,每株测定1次,取平均值。数据导出后用Microsoft Excel 2003处理,Sigmaplot 12.5作图,SPSS 20.0进行相关性分析。
2 结果与分析
2.1 凤丹净光合速率日变化
由图1可以看出,凤丹在春季和夏季光合日变化均呈现出明显的双峰型曲线,即均存在“午休”现象。春季早上气温相对较低,08:00—12:00气温回升,光照渐强,净光合速率随之增大,2个波峰分别在13:00和15:00,且差异不明显;午后光照度达到最大,为避免水分过度蒸发,保护叶片,部分气孔关闭,出现波谷;随后气孔逐渐张开,15:00出现次高峰;15:00 后气温下降,光照减弱,净光合速率在观测时间段内持续下降。夏季气温和光照度均高于春季,导致第一高峰提前,且与次高峰差值很大。09:00为全天中最大值,随后一直下降,13:00时虽有回升,但恢复能力不足,可能是由干旱胁迫和高温造成的光合器官受到损伤所致。

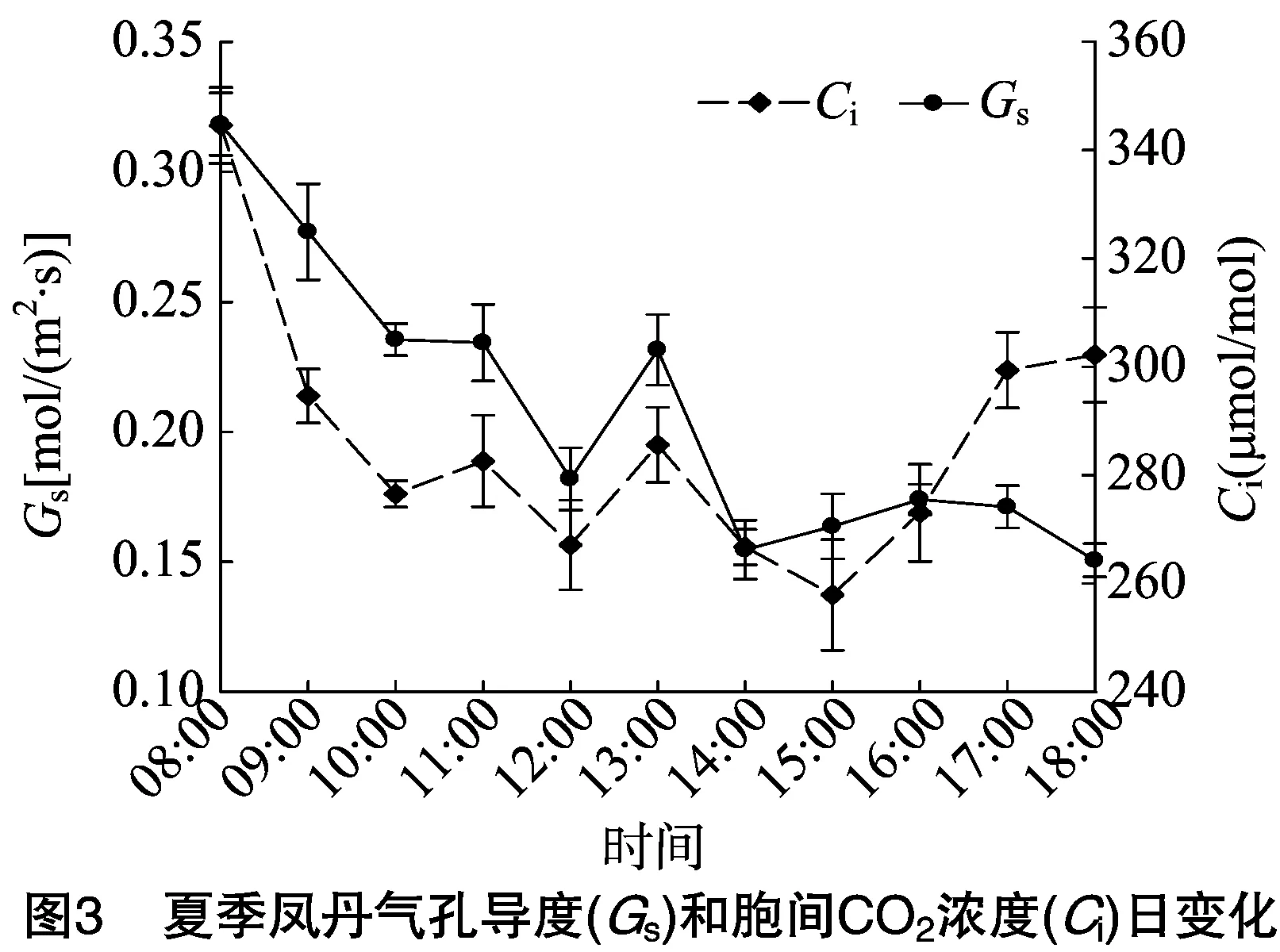
2.2 凤丹气孔导度和细胞间CO2浓度日变化
由图2可以看出,春季凤丹Gs和Ci的日进程在 08:00—18:00有不同的相互关系。12:00前,Gs和Ci变化趋势相同,即Ci伴随Gs的增大而升高,伴随Gs的下降而减小;而12:00—13:00,两者变化趋势相反;13:00—14:00和15:00以后阶段Gs的降低伴随Ci的升高,根据Graham等的研究报道推测,该阶段Gs减小可能是由叶肉细胞光合活性下降造成的非气孔限制因素所致[12-13];15:00以后,由于光强减小,气温下降,13:00—14:00阶段可能是因为叶肉细胞光合机构受损这些可能是导致本研究中Ci增加伴随Gs降低的主要原因。

夏季凤丹气孔导度(Gs)与胞间CO2浓度(Ci)较与春季相比,变化情况差异很大(图3)。09:00—10:00、11:00—12:00、13:00—15:00阶段随着气温升高,光强增大,为防止水分过度丧失,气孔关闭,Ci持续下降,此时Gs下降主要原因推测为气孔限制因素,即气孔关闭导致CO2供应不足;10:00—11:00和15:00以后Ci升高,说明导致此时Gs下降的主要原因为非气孔限制因素。
2.3 凤丹蒸腾速率和水分利用效率日变化
蒸腾速率的变化在一定程度上反映了植物调节水分平衡的能力以及对逆境的适应能力。由图4可知,春季凤丹蒸腾速率的日进程呈双峰型,分别在11:00的5.39 mmol/(m2·s)和15:00的5.46 mmol/(m2·s);波谷在12:00,与此时气孔部分闭合有关。与春季不同,夏季凤丹蒸腾速率日变化轨迹有3个峰值,首个峰值在09:00,为全天中的最大值 6.41 mmol/(m2·s);第2个峰值在13:00,为 6.34 mmol/(m2·s);最后一个峰值在16:00,为 3.98 mmol/(m2·s);波谷分别在12:00与14:00。

水分利用效率WUE(water use efficiency)是衡量蒸腾作用程度的另一个重要指标,其不但可用于描述植物产量和耗水量之间关系,更反映了植物在某一生长时期内能量转换效率及植物耗水性和对水分利用的情况。利用某一时刻净光合速率与蒸腾速率之比,可估算叶片瞬时的水分利用效率,即WUE=Pn/Tr[14]。植物在不同季节、一天中的不同时刻对水分的利用情况是不同的。
图5为凤丹在春季和夏季WUE的日变化情况。从整体上看,凤丹叶片在夏季较春季对水分的利用效率更高,且最高水分利用效率均出现在较早时刻(09:00前);09:00后,2个季节的日变化趋势相反。在春季,11:00时,水分利用情况在一天中最差,而夏季则出现在13:00。

2.4 各生理生态指标相关性分析
由表1和表2可知,大田条件下凤丹在春季Pn与Gs、Tr、Tleaf呈极显著正相关,与VPD、Tair呈显著正相关,与Ci呈极显著负相关;Gs与Tr、PARi呈极显著正相关;Ci与Ca呈显著正相关;Tr与Tleaf、VPD呈极显著正相关,与Tair显著正相关,与RH、WUE呈显著负相关。而在夏季Pn与Gs、PARi呈极显著正相关,与Tr显著正相关;Gs与Ci、Tr显著正相关;Ci与Ca呈显著正相关,与Tair、Tleaf、VPD呈显著负相关;Tr与PARi呈显著正相关,与WUE呈显著负相关。
气孔是CO2和水蒸气进出叶片的通道,气孔的状态直接影响着植物的光合过程和体内外的水分平衡。结合表1和表2可以看出,Gs与Pn、Tr三者之间都呈显著正相关关系,并且不存在季节差异。Ci在春季与Pn呈显著负相关,夏季则没有该现象,但在夏季Tair和Tleaf显著影响Ci。
除气孔因素外,外界气候因子也显著影响着植物的生理活动。Tleaf在春季受Tair、RH、VPD影响显著,通过改变光合作用有关酶的活性来间接影响Pn。在夏季,光合有效辐射PARi成为制约Pn的首要因子。VPD为水汽压亏缺或饱和水汽压差,表示实际空气距水汽饱和状态的程度,反映空气的干燥程度,也是驱动林木蒸腾的主要动力[15]。VPD通过影响气孔的闭合间接影响植物的光合作用和蒸腾作用以及叶片水分的利用效率。春季空气相对湿度RH较小,水汽压亏缺大,VPD值与Pn、Tr呈显著正相关,即在相对干燥的空气中促进光合作用和蒸腾作用,而夏季RH较大,空气相对湿润, 这种效果并不显著,相反提高了水分的利用效率WUE。

表1 春季凤丹光合生理指标与环境因子相关性
注:**、*表示在0.01、0.05水平(双侧)上显著相关(下同)。

表2 夏季凤丹光合生理指标与环境因子相关性
3 结论与讨论
3.1 凤丹的光合“午休”现象
自然条件下,植物光合作用日进程有2种典型的方式:一种为先上升后下降,中午时分达到峰值,即单峰型;另一种为日进程有2个峰值,分别出现在上午和下午的晚些时候,即双峰型,2个峰值间的低谷就是所谓的光合“午休”现象。当午休现象严重时,第2个峰值表现得不明显或消失。本研究中春季凤丹净光合速率日进程呈典型的双峰型,“午休”出现在14:00;而夏季,高温和强光加剧“午休”,次高峰不明显,光合恢复后劲不足,“午休”现象严重。光合作用的“午休”现象,是植物在长期进化过程中形成的对付环境胁迫的一种方法,是对生存有益的调节过程,但却以牺牲光合生产力为代价。Xu等分析造成该现象的种种因子时推测,强光照、低土壤水势和低空气湿度可能是主要的环境因子,气孔导度降低为主要的生理因子,而光系统Ⅱ光化学效率的降低和脱落酸合成的增加是主要的生化因子[16]。因此,在生产实践中可以采取在夏季适当遮阴、及时灌溉等措施来缓解这种现象,尽可能减少光合生产力的损失。
3.2 不同季节凤丹的气孔限制与非气孔限制
植物对多种环境胁迫响应的共同特征是叶片光合速率的降低[17],气候因子具有显著的季节差异。在不同季节,导致凤丹净光合速率下降的原因一是叶肉光合细胞活性降低导致的非气孔限制,二是部分气孔关闭造成的气孔限制,判断依据是前者使Ci升高,后者则使Ci降低。若2种因素同时存在,Ci的变化方向依赖于占优势的那个因素[17-18]。在本试验中,春季上午时段气温较低,光强较弱,净光合速率持续升高,直到午后的14:00才出现下降,非气孔因素是主因,与此时强光和高温有关。在夏季,高温和强光照出现时段提前,加之季节性干旱,气孔因素和非气孔因素则都是导致净光合速率下降的主要因素。
3.3 不同季节凤丹的蒸腾作用和水分利用效率
绿色植物在进行光合作用的过程中,须要通过与周围环境发生气体交换,而这个过程会引起植物水分的动态变化。春季凤丹高蒸腾速率集中在11:00—16:00,而在夏季上午的蒸腾速率较高。相关研究认为,影响蒸腾作用快慢取决于叶片内外的蒸气压差的大小,影响因素包括光照、空气相对湿度、温度、风等[19-20],内部因素有气孔和气孔下腔[21-22]以及叶片内部面积[22]。这与本研究中蒸腾速率与光合有效辐射、大气温度、叶温、水汽压亏缺以及气孔导度呈极显著或显著的正相关关系,与空气相对湿度、水分利用效率呈显著负相关关系的研究结果相一致。
在生产实践中,应尽量维持作物的水分平衡,即使植物吸水量足以达到弥补蒸腾损失水量的状态[23-24]。本研究中2个季节09:00后的水分利用情况大致相反,明显和不同季节空气相对湿度以及水分的亏缺程度有关,这与樊巍等的研究结果[25-26]一致。夏季较春季对水分的利用效率高,这显然与夏季干旱有关。蒸腾速率与水分利用效率呈显著负相关,夏季 13:00 水分利用效率到达一天中的最低值,而此时也正值蒸腾速率的峰值,是否错开该时间段,在11:00—12:00或 14:00—15:00进行适当灌溉以减少水资源浪费来提高凤丹产量还须进一步研究。
参考文献:
[1]李嘉珏,张西方,赵孝庆. 中国牡丹[M]. 北京:中国大百科全书出版社,2011.
[2]付世萃,张大伟,王海龙,等. 吉林省油用牡丹发展前景概述[J]. 湖南林业科技,2014,41(4):78-80.
[3]周琳,王雁. 我国油用牡丹开发利用现状及产业化发展对策[J]. 世界林业研究,2014,27(1):68-71.
[4]叶艳涛,李艳霞. 油用牡丹“凤丹”播种育苗技术[J]. 林业实用技术,2015(11):36-37.
[5]Jaime A. Teixeira da S,Shen M M,Yu X N.tissue culture and micropropagation of tree peony(PaeoniauffruticosaAndr.)[J]. Crop Sci Biotech,2012,15(3):159-168.
[6]Wang H L,Tanaka M. Effect of IBA concentration,carbon source,substrate,and light source on root induction ability of tree peony(PaeoniasuffruticosaAndr.)plantletsinvitro[J]. Europ J Hort Sci,2012,77(3):122-128.
[7]张璐,李玉娥,管世铭,等. 牡丹PsAP1基因的表达分析及功能鉴定[J]. 华北农学报,2015,30(1):84-89.
[8]Li Y X,Wang X. Study on the extraction technology of paeonol from cortex moutan[J]. Medicinal Plant,2011,2(7):67-68.
[9]钱明月,李梅青,吴悠,等. 凤丹籽油理化特性及脂肪酸GC-MS分析[J]. 天然产物研究与开发,2014(26):380-383.
[10]毕玉伟,秦俊,王奎玲,等. 不同季节凤丹光合特性的初步研究[J]. 天津农业科学,2011,17(1):18-21.
[11]张衷华,唐中华,杨逢建,等. 两种主要有用牡丹光合特性及其微环境影响因子分析[J]. 植物研究,2014,34(6):770-775.
[12]Graham D F,Thomas D S. Stomatal conductance and photosynthesis[J]. Rev Plant Physiol,1982,33:317-345.
[13]许大全. 光合作用气孔限制分析中的一些问题[J]. 植物生理学通讯,1997,33(4):241-244.
[14]刘晓荣,廖飞雄,王建新,等. 白鹤芋的光合特性和水分利用效率[J]. 西北农业学报,2015,24(2):142-145.
[15]李炜,司建华,冯起,等. 胡杨(Populuseuphratica)蒸腾耗水对水汽压差的响应[J]. 中国沙漠,2013,33(5):1377-1384.
[16]Xu D Q,Shen Y K. External and internal factors responsible for midday depression of photosynthesis[M]. 2nd ed. Boca Raton:Crc press,2005:287-297.
[17]陈晓亚,薛红卫. 植物生理与分子生物学[M]. 4版. 北京:高等教育出版社,2012:267-279.
[18]杨玉珍,张云霞,彭方仁. 干旱胁迫对不同种源香椿苗木光合特性的影响[J]. 北京林业大学学报,2011,33(1):44-48.
[19]Chen X M,Song J. Effect of juglone fromC.cathayensisexocarp on summer peony photoinhibition[J]. 河南农业大学学报,2007,41(5):527-530.
[20]候小改. 4个牡丹品种光合特性的比较研究[J]. 河南农业大学学报,2007,41(5):527-530.
[21]张文达,蒙宽宏,杨凯. 山杨与白桦蒸腾速率及气孔阻抗特征的研究[J]. 林业勘查设计,2014(4):46-48.
[22]董泽军. 植物蒸腾作用高速率之原因[J]. 中国农学通报,2010,26(21):131-135.
[23]环海军,杨再强,刘岩,等. 鲁中地区夏玉米水分盈亏及灌溉需水量的时空变化特征[J]. 江苏农业科学,2016,44(3):342-347.
[24]胡程达,刘荣花,张永录. 不同水分条件下冬小麦农田蒸散研究[J]. 江苏农业科学,2016,44(7):484-487.
[25]樊巍. 农林复合系统的林网对冬小麦水分利用效率的研究[J]. 林业科学,2000,36(4):16-20.
[26]于文颖,纪瑞鹏,冯锐,等. 不同生育期玉米叶片光合特性及水分利用效率对水分胁迫的响应[J]. 生态学报,2015,35(9):2902-2909.
猜你喜欢
特产研究(2022年6期)2023-01-17
节水灌溉(2021年5期)2021-05-26
植物资源与环境学报(2020年4期)2020-08-10
科学导报(2020年26期)2020-06-09
中国农业科技导报(2020年12期)2020-03-15
天然产物研究与开发(2018年3期)2018-05-07
天然产物研究与开发(2018年3期)2018-05-07
现代园艺(2018年2期)2018-03-15
河北林业科技(2016年5期)2016-11-08
中国林业产业(2016年5期)2016-04-03
