
胶红酵母的成分及其对刺参免疫力和抗氧化能力的影响
2018-04-08 11:19:36张春晓,董琦,张天任,肖珊,王际辉
大连工业大学学报 2018年2期
张 春 晓, 董 琦, 张 天 任, 肖 珊, 王 际 辉
( 1.大连工业大学 食品学院, 辽宁 大连 116034;2.大连工业大学 生物工程学院, 辽宁 大连 116034 )
0 引 言
刺参是我国沿海海域主要的养殖品种之一。随着海参养殖的快速发展,不规范问题的影响逐渐增大,海参的病害问题日益突出,严重制约了该产业的快速发展[1]。目前,预防刺参疾病最有效的方法是使用抗生素,但是这会带来一系列严重的问题,例如耐药性、食品安全和环境污染等[2]。而益生菌可以通过改善刺参体内肠道菌群和刺激免疫系统而达到预防疾病的目的。
红酵母因其含有丰富的营养物质,已经作为一种新型的功能饵料添加剂添加在饲料中,在水产养殖领域得到了初步的应用[3]。研究表明,饵料中添加海洋红酵母不仅能提高幼参肠道消化酶活力,而且能刺激幼参的非特异性免疫反应[4]。徐琴等[5]研究表明,粘红酵母可以促进中国对虾的生长和提高非特异性免疫。目前应用较为广泛的是海洋红酵母,胶红酵母属于海洋红酵母属,胶红酵母是一种单细胞真核生物,属于抗逆性较强的腐生菌,可以促进有益菌生长而抑制有害菌生长,具有较高的营养价值,能够增强机体的免疫力和提高抗氧化能力[6]。但是对其研究比较少,尤其是在刺参养殖方面。
本实验对胶红酵母的营养成分进行分析,研究了其对刺参免疫力及抗氧化能力的影响,以期为胶红酵母在刺参养殖中的应用提供理论基础。
1 实 验
1.1 胶红酵母的营养成分分析
胶红酵母为实验室保存菌株,于YPD培养基中180 r/min、28 ℃发酵培养至稳定前期。发酵液中菌体的浓度约为4×108CFU/mL。
1.1.1粗营养成分
水分含量采用冷冻干燥法测定;粗蛋白含量采用凯氏定氮法(GB 5009. 5—2010)测定;粗脂肪含量采用索氏抽提法(GB/T 5009.6—2003)测定;灰分含量采用550 ℃灼烧重量法(GB 5009.4—2010)测定;总糖含量采用DNS法测定。
1.1.2氨基酸
游离氨基酸含量根据GB/T 5009.124—2003进行测定。
1.1.3脂肪酸
样品用氯仿-甲醇提取脂肪酸,脂肪酸提取物在碱性条件下进行皂化和甲酯化。采用气相色谱测定脂肪酸甲酯的含量。结果用归一化法计算。
1.1.4葡聚糖
采用碱-酶法提取葡聚糖[7],用高效液相色谱法测定葡萄糖质量分数,经计算得出胶红酵母中葡聚糖的含量。
1.1.5β-胡萝卜素
β-胡萝卜素质量分数的测定参照GB/T 5009.83— 2003第一法,利用高效液相色谱测定。
1.1.6VE
新鲜菌泥经过皂化、提取、洗涤和浓缩后得到提取物。根据GB/T 5009.82—2003第一法,利用高效液相色谱法测定VE质量分数。
1.2 饲养实验
1.2.1饵料制备
用灭菌海水调整菌液浓度为108CFU/mL。将菌液与饵料以107CFU/g混合均匀。饵料每天配制以保证胶红酵母的活性。
1.2.2实验动物
实验用幼刺参规格一致,均为(4.08±0.42) g,所有刺参经过7 d的基础喂养以适应环境。分到6个23.5 cm×23.5 cm×29 cm水缸中,每个水缸大约有200 g刺参。
1.2.3实验设计
实验设一个空白对照组和一个实验组。空白对照组用基础饵料喂养,实验组在基础饵料的基础上添加胶红酵母的浓度为107CFU/mL。每个处理组设3个平行,整个养殖实验周期为60 d。
在整个养殖实验进行过程中,每天17:00投喂饵料,饵料的投喂量为刺参体重的1.5%。实验期间每周换一次全水,中间换一次半水,水温保持在17~20 ℃,pH 7.7~7.9,盐度3.0%~3.3%。用增氧泵间隔充氧,使溶氧量不低于6 mg/L。
1.2.4样品采集
取样前48 h停止投喂饵料,并清理残饵,使肠道内容物排空。解剖时取刺参体腔液与等量抗凝剂混合,得到抗凝体腔液。所有的样品液氮速冻后置于-80 ℃冰箱中储存。实验前,样品解冻后于3 000g、4 ℃离心10 min,用移液枪取上清液,分装在1.5 mL离心管中,-20 ℃储存,用于后续实验。
1.2.5免疫酶活性的测定
酸性磷酸酶(ACP)、碱性磷酸酶(AKP)活性测定、溶菌酶(LZM)、总超氧化物歧化酶(T-SOD)和一氧化氮合酶(NOS)的活性测定均采用南京建成生物工程研究所的检测试剂盒进行测定。
1.2.6抗氧化能力的测定
过氧化氢酶(CAT)、谷胱甘肽过氧化物酶(GSH-PX)、总抗氧化能力(T-AOC)活性和丙二醛的含量均采用南京建成生物工程研究所的检测试剂盒进行测定。
1.3 数据分析
实验数据用(平均值±标准差)表示,并采用统计软件SPSS 20.0进行单因素方差分析,不同处理之间采用 Ducan’s多重比较,以P<0.05表示显著差异。
2 结果与讨论
2.1 胶红酵母的营养成分分析
2.1.1粗营养成分
胶红酵母菌粉中粗蛋白、粗脂肪、灰分和总糖质量分数分别为57.37%、2.79%、5.08%和19.44%。蛋白质是胶红酵母菌体细胞的主要组成成分,在饵料中添加酵母可以满足水产养殖动物对生物饵料的蛋白质需求[8]。总糖可以为生物体提供能量,胶红酵母中的总糖主要来源于酵母细胞壁中的葡聚糖,而葡聚糖可以刺激生物体的免疫反应[9]。
2.1.2氨基酸组成
如表1所示,胶红酵母菌体中共检测出18种游离氨基酸。Rhishipal等[10]对海洋酵母的研究表明,鱼类和甲壳类动物生长需要精氨酸、缬氨酸、苏氨酸、蛋氨酸、亮氨酸、异亮氨酸、苯丙氨酸、赖氨酸、组氨酸、色氨酸10种氨基酸,本研究结果表明胶红酵母中这10种氨基酸,可以促进刺参的生长。此外,胶红酵母菌体中还含有丰富的鲜味氨基酸(谷氨酸、天冬氨酸、苯丙氨酸、丙氨酸、甘氨酸、酪氨酸),占总氨基酸的质量分数为2.43%。

表1 胶红酵母菌体的氨基酸组成Tab.1 Composition of amino acids in Rhodotorulamucilaginosa
研究表明,氨基酸对鱼类的诱食及嗅觉都有极强的刺激作用,尤其是L-氨基酸,已被公认为是引诱鱼、甲壳类、昆虫和其他水产动物最有效的化合物之一[11],在一定程度上具有诱食作用,可以更好地促进海参进食。以上结果表明,此胶红酵母菌体中氨基酸种类丰富,适合于作为饵料添加剂应用于刺参养殖。
2.1.3脂肪酸组成
如表2所示,胶红酵母菌体中不饱和脂肪酸占总脂肪酸的82.02%,其中C18:1的含量最高,占总脂肪酸的76.40%。不饱和脂肪酸是鱼类生长所必需的能量和营养物质,在机体内具有广泛的生理功能和生物学效应,可以调节免疫系统[12]。但是酵母菌体中缺少EPA和DHA,可以在饵料中添加富含这两种多不饱和脂肪酸的藻类作为补充。此胶红酵母菌体不饱和脂肪酸的含量丰富,可以改善饵料的结构,提高机体的功能。
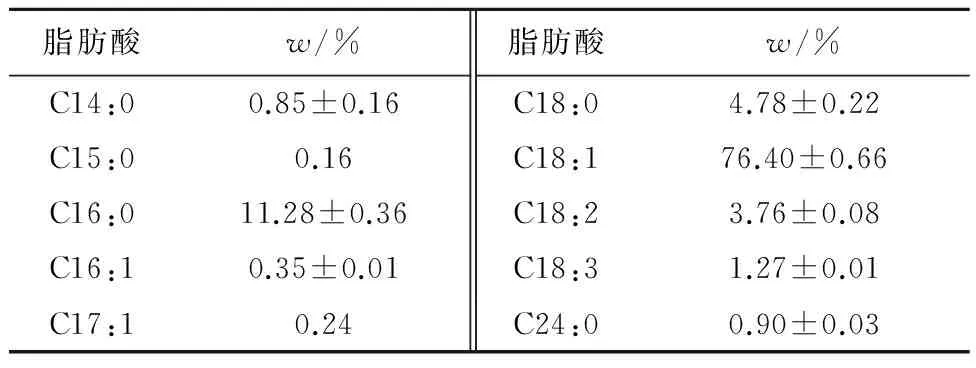
表2 胶红酵母菌体的脂肪酸组成Tab.2 Composition of fatty acids in Rhodotorulamucilaginosa
2.1.4葡聚糖、β-胡萝卜素和VE质量分数
葡聚糖是一种免疫多糖,可以增强巨噬细胞和嗜中性粒细胞的迁移和吞噬,减少肠道炎症反应和增强机体抵抗病原体的能力。葡聚糖是酵母细胞壁的主要成分,胶红酵母中葡聚糖的质量分数为3.41%,约占酵母细胞壁干质量的26%~30%[13]。β-胡萝卜素是VA的前体物质,具有促进机体生长、增强机体免疫力和提高抗氧化能力的作用,胶红酵母中β-胡萝卜素的质量分数为17.05 μg/g。有研究表明,饵料中添加β-胡萝卜素可以促进刺参生长和提高抗氧化能力[14]。目前已经广泛应用于食品工业、保健品及饵料添加剂等方面。VE是一种非酶抗氧化物质,可以保护细胞膜和亚细胞膜不被氧化,胶红酵母中VE的质量分数为535.54 μg/g。研究表明,饵料中添加60.5~98.8 mg/kg的VE能促进花鲈的生长和提高其免疫力[15]。近年来研究还发现,在饵料中添加VE可以提高机体的抗应激能力[16]。
2.2 免疫活性
饵料中添加胶红酵母对刺参免疫活性的影响如表3所示。结果表明,在饵料中添加浓度为107CFU/g 的胶红酵母,刺参体腔液中的ACP、AKP、LZM和T-SOD的活性与空白对照组相比显著提高,且较空白组提高了33.08%、50.00%、107.14%、17.11%。刺参体腔液中的NOS活性与空白对照组相比有所提高,但没有显著性差异。
表3饵料中添加胶红酵母对刺参免疫活性的影响
Tab.3Effects of dietary addition ofRhodotorulamucilaginosaon immunity ofApostichopusU/mL
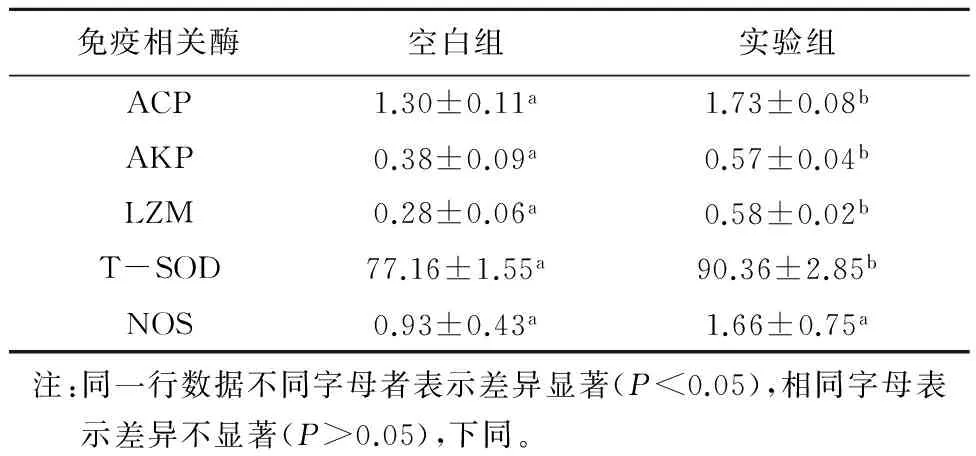
免疫相关酶空白组实验组ACP1.30±0.11a1.73±0.08bAKP0.38±0.09a0.57±0.04bLZM0.28±0.06a0.58±0.02bT-SOD77.16±1.55a90.36±2.85bNOS0.93±0.43a1.66±0.75a注:同一行数据不同字母者表示差异显著(P<0.05),相同字母表示差异不显著(P>0.05),下同。
在刺参的免疫体系中,体液免疫是刺参抵抗外来微生物入侵的主要防御体系。本实验的研究表明,胶红酵母中含有丰富的免疫活性物质,包括葡聚糖、β-胡萝卜素和VE。ACP是机体内巨噬细胞溶菌体的标志酶,AKP是动物体内一种重要的调控酶[17],它们是机体内两种重要的水解酶,参与磷酸基团的转移以及蛋白质的合成代谢。宫魁等[18]研究表明,破壁酵母可以提高ACP活性和AKP活性。本实验的研究结果表明,饵料中添加胶红酵母可以显著提高ACP活性和AKP活性,与其研究结果一致。LZM存在于海参的各个组织中,它的活性是免疫学的重要指标,主要通过作用于被吞噬物质的细胞壁起到杀菌的作用[19]。
2.3 抗氧化活性
如表4所示,饵料中添加胶红酵母能显著提高CAT活性、GSH-PX活性和降低MDA浓度,且CAT活性和GSH-PX活性与空白组相比提高了17.46%和142.86%,MDA浓度较空白组降低了70.00%。但对于T-AOC的提高不显著。生物体内抗氧化防御系统包括抗氧化酶类和抗氧化剂类,其中抗氧化酶类包括CAT和GSH-PX等,它们可以消除体内过量的NOS,将其控制在安全范围内,避免生物体遭到氧化损伤[20]。
表4饵料中添加胶红酵母对刺参抗氧化活性的影响
Tab.4Effects of dietary addition ofRhodotorulamucilaginosaon antioxidant activity ofApostichopusjaponicus
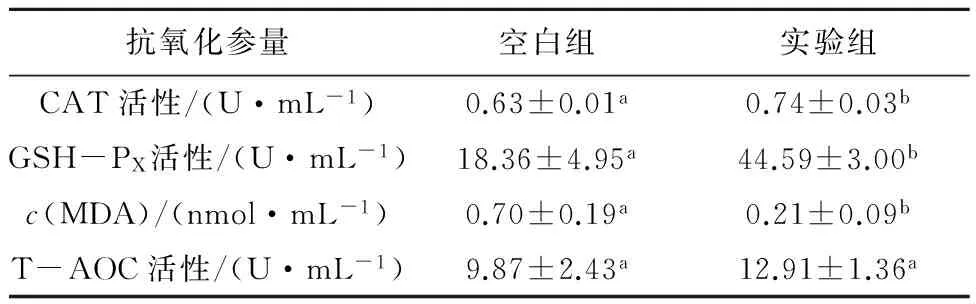
抗氧化参量空白组实验组CAT活性/(U·mL-1)0.63±0.01a0.74±0.03bGSH-PX活性/(U·mL-1)18.36±4.95a44.59±3.00bc(MDA)/(nmol·mL-1)0.70±0.19a0.21±0.09bT-AOC活性/(U·mL-1)9.87±2.43a12.91±1.36a
饵料中添加胶红酵母可以提高CAT活性和GSH-PX活性,可能与胶红酵母中含有大量的β-胡萝卜素和 VE两种具有抗氧化能力的物质有关。MDA是生物体在逆境条件下发生脂质过氧化作用的产物之一,其浓度代表膜脂过氧化作用的强弱[21]。研究表明,MDA浓度越低,说明机体内过氧化产物越少。本实验的结果表明,实验组刺参的MDA含量明显低于空白对照组,说明实验组的海参能够更好地适应环境的变化,提高机体的应激能力。T-AOC是用于衡量机体抗氧化体系的综合性指标,其大小可代表和反应机体对外来刺激的抵抗能力以及机体自由基的代谢状态,主要作用是分解和清除代谢过程中产生的活性氧自由基[22]。饵料中添加胶红酵母对于T-AOC 的提高不显著,说明实验组整个抗氧化系统的抗氧化能力有所提高,但是提高不显著。
3 结 论
胶红酵母菌体中营养物质丰富,营养价值较高,胶红酵母菌体中粗蛋白质量分数为57.37%,不饱和脂肪酸占总脂肪酸的82.03%。胶红酵母含有丰富的活性物质,包括葡聚糖、β-胡萝卜素和VE,质量分数分别为3.41%、17.05 mg/kg、535.54 mg/kg,是较好的饵料添加剂。添加107CFU/g 胶红酵母可以显著提高刺参酸性磷酸酶、溶菌酶、碱性磷酸酶、总超氧化物歧化酶、过氧化氢酶、谷胱甘肽过氧化物酶的活性,降低丙二醛的浓度。表明在饵料中添加胶红酵母可以在一定程度上提高刺参的免疫力和抗氧化能力。
参考文献:
[1] 王印庚,荣小军,张春云,等.养殖海参主要疾病及防治技术[J].海洋科学,2005,29(3):1-7.
[2] YANG A F, ZHOU Z C, HE C B, et al. Analysis of expressed sequence tags from body wall, intestine and respiratory tree of sea cucumber (Apostichopusjaponicus) [J]. Aquaculture, 2009, 296(3): 193-199.
[3] 汪洋,孔维宝,韩锐,等.红酵母简介[J].生物学通报,2015(4):15-17.
[4] 杨志平,徐哲,周倩,等.饵料中添加海洋红酵母C11对幼参消化酶及免疫反应的影响[J].渔业科学进展,2015,36(6):107-112.
[5] 徐琴,李健,刘淇,等.噬菌蛭弧菌和粘红酵母对中国对虾生长及非特异免疫因子的影响[J].渔业科学进展,2007,28(5):42-47.
[6] 叶伟庆,吴园园,高上吉,等.4株海洋红酵母分离鉴定及其代谢产物分析[J].热带作物学报,2013,34(10):2046-2050.
[7] 徐希柱,辛培超.不溶性酵母多糖分析方法的研究[J].中国酿造,2008(13):80-83.
[8] SHIAU S Y. Nutrient requirements of penaeid shrimps [J]. Aquaculture, 1998, 164(1/2/3/4): 77-93.
[9] 韩梅,翟玉贵,王小波,等.锁掷酵母与海洋红酵母的营养成分分析[J].食品与发酵工业,2013,39(1):11-15.
[10] RHISHIPAL R, PHILIP R. Selection of marine yeasts for the generation of single cell protein from prawn-shell[J]. Bioresource Technology, 1998, 65(3): 255-256.
[11] 杨鸢劼,邴旭文,徐增洪.不饱和脂肪酸对黄鳝部分非特异性免疫和代谢指标的影响[J].中国水产科学,2008,15(4):600-605.
[12] 关洪斌,李宁,薛佳萤,等.氨基酸强化饵料对大鲮鲆诱食活性的研究[J].氨基酸和生物资源,2009,1(3):25-29.
[13] DALMO R A, BOGWALD J. β-glucans as conductors of immune symphonies[J]. Fish and Shellfish Immunology, 2008, 25(4): 384-396.
[14] 王吉桥,樊莹莹,徐振祥,等.饲料中β-胡萝卜素和虾青素添加量对仿刺参幼参生长及抗氧化能力的影响[J].大连海洋大学学报,2012,27(3):215-220.
[15] 周立斌,张伟,王安利,等.饲料维生素E添加量对花鲈生长、组织中维生素E积累量和免疫指标的影响[J].水产学报,2009(1):95-102.
[16] 施兆鸿,张艳亮,高权新,等.饲料维生素E水平影响云纹石斑鱼幼鱼对氨氮胁迫的响应[J].动物营养学报,2015,27(5):1596-1604.
[17] 牟海津,江晓路.免疫多糖对栉孔扇贝酸性磷酸酶,碱性磷酸酶和超氧化物歧化酶活性的影响[J].中国海洋大学学报(自然科学版),1999(3):463-464.
[18] 宫魁,王宝杰,刘梅,等.全营养破壁酵母对仿刺参非特异性免疫及肠道菌群的影响[J].中国水产科学,2012,19(4):641-646.
[19] CANICATTI C, ROCH P. Studies onHolothuriapolii(Echinodermata) antibacterial proteins.Ⅰ. Evidence for and activity of coelomocyte lysozyme[J]. Cellular and Molecular Life Sciences, 1989, 45(8): 756-759.
[20] SILVA A M M, NOVELLI E L B, FASCINELI M L, et al. Impact of an environmentally realistic intake of water contaminants and superoxide formation on tissues of rats[J]. Environmental Pollution, 1999, 105(2): 243-249.
[21] 沙飞,常亚青,丁君.两种降温模式的低温胁迫对刺参抗氧化酶活性及丙二醛含量的影响[J].大连海洋大学学报,2015,30(1):25-29.
[22] 张春玲,胡俊峰,王丕文,等.苯并(a)芘对鲫鱼肝脏总抗氧化能力的影响[J].环境与健康杂志,2004,21(5):325-326.
猜你喜欢
领导文萃(2022年15期)2022-07-06 07:13:48
杂文选刊(2022年7期)2022-06-30 11:18:35
文萃报·周五版(2022年15期)2022-04-21 12:15:19
红蜻蜓·低年级(2021年12期)2022-01-19 05:18:32
红蜻蜓·低年级(2021年12期)2021-12-19 15:06:23
大连海洋大学学报(2020年2期)2020-05-06 02:26:10
现代农业(2016年6期)2016-02-28 18:42:51
新少年(2015年5期)2015-07-10 09:38:04
食品工业科技(2014年15期)2014-03-11 18:17:20
现代检验医学杂志(2014年3期)2014-02-02 02:42:18
