
室旁核内脂肪酸酰胺水解酶上调介导慢性心衰大鼠交感神经兴奋亢进*
2018-04-02 07:34王仁俊未晓巍赵永斌齐云峰周晓馥
中国病理生理杂志 2018年3期
王仁俊,周 琴,未晓巍,李 华,赵永斌,齐云峰,栾 剑,周晓馥
(吉林师范大学生命科学学院生物技术系,吉林 四平 136000)
慢性心力衰竭(chronic heart failure,CHF)的临床病理特征之一为交感神经兴奋性增强[1]。CHF状态下中枢神经系统中相关作用机制改变可导致交感神经兴奋性增强,研究报道显示CHF状态下交感活动亢进是下丘脑室旁核(paraventricular nucleus,PVN)神经递质、调质、微小核苷酸或离子通道等内源性物质整合作用失衡的结果[2-3],但PVN内的大麻素类物质是否参与了CHF交感神经激活作用,迄今为止,未见相关报道。
N-花生四烯酰乙醇胺(N-arachidonoylethanolamide,AEA)是一种内源性大麻素样物质,首次发现AEA合成是在神经元中。研究显示多种类型的神经递质是以储存的形式释放,但AEA的释放方式却是“按需求”合成,其分布位置主要集中在内皮细胞、血管平滑肌、心肌及中枢神经系统等[4],已有研究显示AEA的酶降解过程主要为脂肪酸酰胺水解酶(fatty-acid amide hydrolase,FAAH)途径,FAAH在脑中含量高,且能有效水解AEA[5]。在大脑半球中注入FAAH抑制剂PF3845可降低FAAH活性,增加AEA在脑组织中的含量[6-7],即在大脑皮层中AEA含量及FAAH表达量呈负相关。另有文献报道,AEA在抗心肌缺血和抗心律失常等诸多方面发挥重要的保护作用。另外,在肺动脉中的AEA具有扩血管并降低血压的作用[8];在右心房注射AEA可较大程度地增强肺迷走传入神经的敏感性[9],以上研究结果提示AEA在外周对心血管起保护作用。
在中枢神经系统中,增加终纹核中内源性的AEA水平,可有效抑制心动过速[10]; 此外,在海马区中AEA能够增强N-甲基-D-天冬氨酸受体调控的电流[11],N-甲基-D-天冬氨酸受体激活能促进Ca2+内流,引起心肌收缩;将AEA注射到中脑导管周围灰质中,可诱导产生一定量的一氧化氮[12],一氧化氮具有扩张血管的作用,进而降低动脉血压;孤束核中微量注射AEA能够延长压力反射时间,从而抑制交感神经活性[13];在麻醉大鼠PVN内注射AEA可明显降低动脉血压[14]。这些均提示AEA在中枢神经系统中参与了交感兴奋调节及血压反射调控。但在PVN内FAAH和AEA是否参与CHF状态下的交感神经兴奋激活过程,目前尚未见相关报道。本文拟用实验研究验证PVN内上调的FAAH通过抑制AEA的生成介导CHF状态下交感神经兴奋亢进作用这一假说。
材 料 和 方 法
1 实验动物
雄性Wistar大鼠,体重180~220 g,购于长春市亿斯实验动物技术有限责任公司,合格证号:SCXK(吉)2011-0004。基础饲料由长春市亿斯实验动物技术有限责任公司提供,生产许可证号:SCXK(吉)2010-0001。动物生活环境恒温(23±2) ℃,常规24 h昼夜循环。
2 药品、试剂与实验设备
AEA和PF3845等药品和试剂购自长春鼎国生物技术有限公司。重组2型腺相关病毒(recombinant adeno-assciated virus type 2,rAVV2)购自山东维真生物科技有限公司。脑立体定位仪(NARISHIGE,SR-5M-HT);多通道生理信号采集系统(成都泰盟有限公司);激光共聚焦显微镜系统(ZEISS,LSM 710)等。
3 方法
3.1CHF模型的构建取Wistar大鼠,麻醉固定后,分离气管,连接呼吸机,于3~4肋骨间打开胸腔,使其心脏暴露后,小心剪开心包膜,CHF组大鼠用0号带线缝合针结扎其左冠状动脉,假手术组仅穿线,不结扎,手术缝合。术后注射青霉素抗感染,每天1次,连用3天,常规饲养8周后行急性实验。
3.2监测大鼠超声心动图和血流动力学指标用30%乌拉坦及α-氯醛糖腹腔注射麻醉实验动物,采用心脏超声心动图仪检测左心室射血分数(ejection fraction,EF)、短轴缩短率(fractional shortening,FS)、左心室舒张末期容积(left ventricular end-diastolic volume,LVEDV)和左心室舒张末期内径(left ventri-cular end-diastolic diameter,LVEDD)等。先后分离股动脉和颈总动脉并插管,通过压力换能器连接多道生理记录仪,采集平均动脉压(mean arterial pressure,MAP)、左心室收缩压(left ventricular systolic pressure,LVSP)和左心室舒张末期压(left ventricular end-diastolic pressure,LVEDP)等血流动力学指标,俯卧固定麻醉大鼠,分离肾交感神经,记录肾交感神经放电活动(renal sympathetic nerve activity,RSNA)和标准Ⅱ导联心电图,并同步测量心率(heart rate,HR)。
3.3PVN微量注射AEA或PF3845参照Paxinos & Watson大鼠脑图谱,将麻醉大鼠固定于脑立体定位仪上,于下丘脑PVN(前囟后1.7 mm,L和R均为 0.5 mm,H为7.2 mm)微量注射PF3845(0.5、1和2 nmol/200 nL,溶剂为含1%二甲亚砜的生理盐水)或AEA(2、4和8 nmol/200 nL,溶剂为含0.01% Tocrisolve 100的人工脑脊液,Tocrisolve 100为豆油和水按1∶4比例形成的水溶性乳状液,其乳化剂为Pluronic F-68),注射速度为5 μL/h,注射时间1 h,PVN双侧注射,每侧注射100 nL,实时监测MAP、HR和RSNA等指标。
3.4静脉注射AEA将麻醉大鼠仰卧固定,于其股静脉注射不同浓度的AEA(4、8和16 nmol/0.1 mL),实时监测MAP、HR和RSNA等指标。
3.5携带FAAHshRNA的rAAV2载体的构建针对GeneBank中大鼠FAAH基因序列(NM 024132.3),按照RNA干扰的设计原则设计出可靶向干扰大鼠FAAH基因转录的特异性片段,具体序列为5’-TTGCTGACCCTGCCCCTACTCCAACTGGT-3’,同时设计其能编码shRNA的DNA链(表1),其中包含一个茎环结构及BamH I 和EcoR I的双酶切位点,使用BamH I 和EcoR I酶双酶切 pSIREN载体,使其线性化,再将目的基因片段与线性化的pSIREN载体进行连接,在连接反应体系中其退火的双链DNA(表2),连接成功后将其转化进入感受态细胞中,选择转化成功的感受态阳性细胞转染rAAV2,筛选出第1代毒种并进行毒种扩增,即携带FAAHshRNA的rAAV2载体的扩增。

表1 构建FAAH shRNA寡核苷酸框架Table 1.Building a framework for FAAH shRNA oligonucleotides
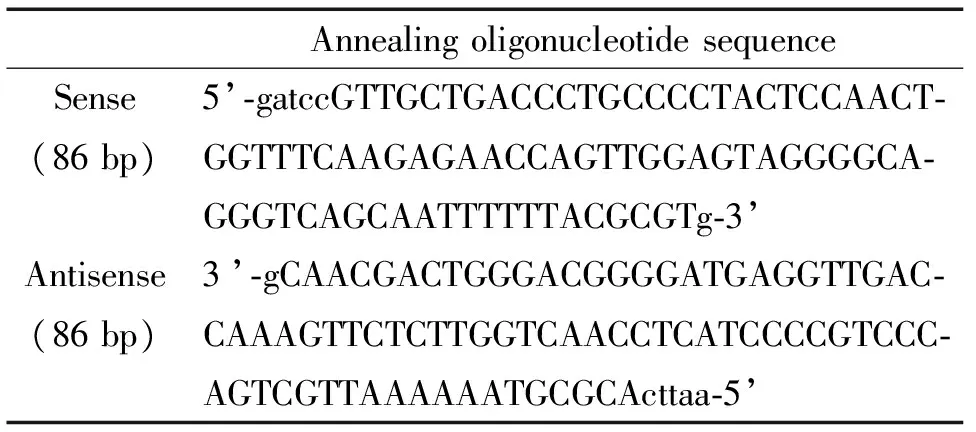
表2 构建退火双链DNA寡核苷酸框架Table .Building a framework of the annealing double-stranded DNA oligonucleotides
3.6PVN内转染FAAHshRNA病毒敲减PVN内FAAH基因将大鼠置于含异氟烷的诱导盒内气体麻醉,待麻醉成功后将其俯卧固定于立体定位仪,带上呼吸面罩,以维持手术过程中麻醉,头部剃毛消毒后,于体式显微镜下,参照大鼠脑立体定位图谱定位,消毒棉球擦拭去除头骨表面结蹄组织及骨膜,用电钻钻出一个小孔,将微量注射器固定在操纵器上,以1 000 μm/min的速度进入脑内,总深度为7.2 mm。到达PVN后稳定10 min,以0.1 μL/min的速度匀速注射事先制备的携带目的基因干扰序列的rAAV2-hSyn-FAAH-shRNA病毒(FAAH-shRNA组)和rAAV2-hSyn-scramble-WPRE-pA病毒(Sc-shRNA组),PVN双侧注射总剂量为200 nL,单侧注射100 nL,滴度为1×1017vg/L,注射完毕后静置20 min,再以1 000 μm/min的速度退出。结束后,以骨蜡封闭小孔,碘伏消毒后缝合,术后肌肉注射青霉素1周抗感染。
3.7Western bolt实验检测PVN内FAAH的蛋白表达水平取实验大鼠PVN组织分别加入蛋白提取缓冲液(10 mmol/L Tris、1 mmol/L EDTA、1% SDS、0.1% Triton X-100和1 mmol/L PMSF),超声粉碎,37 ℃孵育5 min;4 ℃、12 000 r/min离心8 min;上清液即为全细胞蛋白,BCA法测定蛋白浓度;分别在各组取等量的蛋白样品经SDS-PAGE分离后,滤纸-膜-滤纸样的三明治法电转移至NC膜;用封闭液(含5%脱脂奶粉)室温封闭过夜;I 抗4 ℃孵育过夜后,PBS洗涤3次,每次10 min,然后用辣根过氧化物酶荧光标记的 II 抗分别对膜避光孵育1 h,再用PBS洗涤3次,每次10 min,然后用ECL固定,最后显影定影,扫膜。
3.8高效液相色谱检测PVN内AEA的含量取冷冻的PVN组织精密称量后,放入加有水解酶抑制剂PMSF的玻璃瓶中,加入乙腈溶剂20 mL,匀浆打碎后,进行为时1 h的超声。4 ℃静置过夜,沉淀蛋白。将沉淀物全部放入离心机(10 000 r/min)离心10 min,收集上层有机相。利用固相萃取柱过滤收集的有机相,首先加入2 mL流动相,然后使用乙酸乙酯∶丙酮(比例1∶1)共5 mL进行洗脱,收集滤液,再用氮气吹干洗脱液。顺着烧杯边缘加入1 mL乙腈,再涡旋仪混合,复溶,放入-80 ℃冰箱保存备用。
取上述处理得到的样品,精密吸取0.5 mL乙腈样品溶液于离心管,加入0.5 mL AEA衍生化试剂DBD-COCL,恒温50 ℃反应 2 h。加入100 μL去离子水终止反应。进样高效液相色谱检测分析。
3.9组织免疫荧光观察心脏灌流,取脑,4%多聚甲醛固定,蔗糖梯度脱水,冰冻切片,切片厚约25 μm,取大鼠室旁核大脑切片和腰4背根神经节(高表达FAAH)组织切片[作为阳性对照(positive control,PC)],置于PBS中浸泡10 min,以去除OCT; 用含10%正常山羊血清的PBS室温封闭切片1 h; 加入兔抗大鼠FAAH I抗,阴性对照(negative control,NC)采用的I抗是兔IgG,湿盒4 ℃冰箱孵育过夜; PBS洗3次,每次15 min;加入红色荧光标记的山羊抗兔的IgG II 抗,湿盒4 ℃冰箱孵育过夜;PBS洗3次,每次10 min;中性树脂封片后,用共聚焦扫描显微镜观察结果并拍照。
3.10ELISA检测血浆中去甲肾上腺素(norepinephrine,NE)水平腹主动脉采血法取各实验组大鼠动脉血,在腹主动脉分叉处向心端1~3 mm处穿刺并采血。4 ℃ 离心20 min,放置于-80 ℃冰箱待后续检测,具体步骤按照NE水平ELISA检测试剂盒说明书操作,通过酶标仪读取并分析数据。
3.11心脏组织病理学观察抽取1%伊文氏蓝2 mL推入腹腔静脉,1 min后迅速剪下心脏,冲洗后沿心脏长轴通过心脏切片器横切5~6片1 mm薄片,随后置于含3% 2,3,5-氯化三苯基四氮唑的培养皿中37 ℃孵育20 min。梗死面积(infarct size,IS;%)=梗死区面积/心脏切片的总面积×100%。该比值≥30%为冠脉结扎术诱导CHF模型成功标志。
4 统计学处理
所有实验数据通过Prism 5.0软件进行数据处理和统计分析。数据均采用均数±标准误(mean±SEM)表示,采用单因素方差分析和Newman-Keuls检验进行多重比较或采用Student’st检验。以P<0.05表示差异有统计学意义。
结 果
1 CHF模型成功建立
左冠脉结扎术后 8周,利用超声心动图等技术对假手术组及CHF组大鼠各指标进行检测,结果显示,与假手术组相比,CHF组大鼠的LVEDP显著升高,左心室内压最大上升速率(dp/dt)和FS明显下降(P<0.05);CHF组大鼠的肺体比和心脏重量均明显增加(P<0.05),提示CHF大鼠有肺淤血等CHF迹象;CHF组大鼠心肌梗死面积明显增加(P<0.05),其心肌梗死面积在32%~41%之间,而假手术组大鼠并未出现心肌梗死的现象,见表3。以上结果均显示CHF大鼠心脏结构和功能均发生显著变化,CHF模型构建成功。
2 PVN注射位点鉴定
药物注射完毕后,向PVN中注入2%的滂恩天蓝50 nL,腹腔注入过量乌拉坦处死大鼠,迅速取脑。于冰冻切片机切片,切片厚度约为40 μm,体式显微镜下观察其注射位点,参照Paxinos & Watson大鼠脑图谱,结果显示核团注射成功,见图1。
表3CHF组与假手术组大鼠解剖学、血流动力学和交感驱动生理学指标检测
Table 3.The indexes of anatomy,hemodynamics and sympathetic drive in the CHF rats and the sham rats (Mean±SEM.n=6)
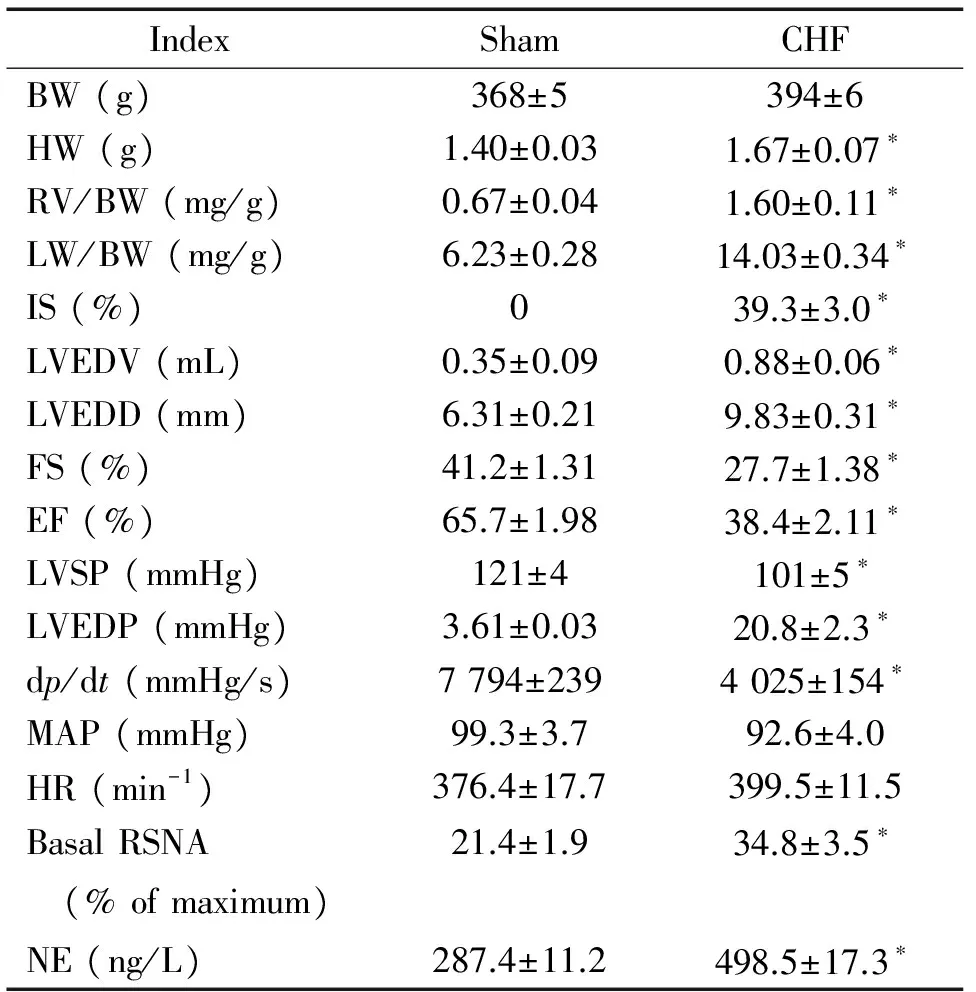
IndexShamCHFBW(g)368±5394±6HW(g)1.40±0.031.67±0.07∗RV/BW(mg/g)0.67±0.041.60±0.11∗LW/BW(mg/g)6.23±0.2814.03±0.34∗IS(%)039.3±3.0∗LVEDV(mL)0.35±0.090.88±0.06∗LVEDD(mm)6.31±0.219.83±0.31∗FS(%)41.2±1.3127.7±1.38∗EF(%)65.7±1.9838.4±2.11∗LVSP(mmHg)121±4101±5∗LVEDP(mmHg)3.61±0.0320.8±2.3∗dp/dt(mmHg/s)7794±2394025±154∗MAP(mmHg)99.3±3.792.6±4.0HR(min-1)376.4±17.7399.5±11.5BasalRSNA (%ofmaximum)21.4±1.934.8±3.5∗NE(ng/L)287.4±11.2498.5±17.3∗
*P<0.05vssham group.
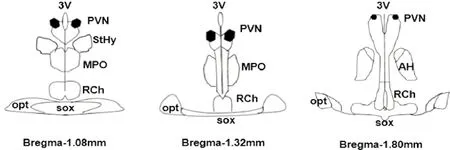
Figure 1.Schematic drawings of rat hypothalamus in coronal section.PVN:paraventricular nucleus; AH:anterior hypothalamic area; 3V:the third cerebral ventricle; RCh:retrochiasmatic area; MPO:medial preoptic nucleus; opt:optic tract; sox:supraoptic decussation; StHy:striohypothalamic nucleus.
图1大鼠下丘脑PVN微量注射靶点所在脑区冠状切片示意图
3 CHF大鼠PVN内AEA含量、FAAH蛋白表达及免疫荧光检测
冠脉结扎8周后,对CHF大鼠PVN内AEA含量及FAAH蛋白表达进行检测,结果显示,与假手术组相比,CHF组PVN内AEA含量减少,FAAH蛋白表达量及FAAH阳性神经元数量显著增加(P<0.05),见图2。这些结果提示PVN内AEA含量的减少以及FAAH蛋白表达的上调可能与心衰的发生发展有关。
4 PVN注射PF3845对交感驱动和心功能指标的影响

Figure 2.The AEA content (A),FAAH protein expression (B) and average number of FAAH-positive neurons (C) in PVN in the rats with CHF.Scale bar=20 μm.Mean±SEM.n=6.*P<0.05vssham group.
图2大鼠PVN内AEA含量、FAAH蛋白表达及FAAH阳性神经元数量
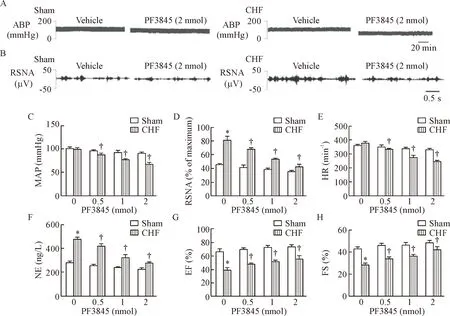
Figure 3.The effect of microinjection of PF3845 to PVN on the sympathetic drive and cardiac function indexes.A:original tracings of artery blood pressure (ABP); B:original tracings of renal sympathetic nerve activity (RSNA); C:mean arterial pressure (MAP); D:renal sympathetic nerve discharge (RSND); E:heart rate (HR); F:plasma norepinephrine (NE); G:ejection fraction (EF); H:fractional shortening (FS).Mean±SEM.n=6.*P<0.05vssham+vehicle (0 nmol PF3845) group;†P<0.05vsCHF+vehicle (0 nmol PF3845) group.
图3PVN内微量灌注PF3845对交感驱动和心功能指标的影响
与溶剂对照组相比,CHF组大鼠PVN内微量注射FAAH抑制剂PF3845导致MAP、HR和RSNA和血浆NE明显降低,同时EF和FS增加(P<0.05),在注射25~40 min时变化最为明显,见图3。结果提示抑制FAAH可能导致AEA含量增加,从而通过抑制交感驱动改善心衰大鼠心脏功能。
5 PVN注射AEA对交感驱动和心动能指标的影响
与溶剂对照组相比,微量注射AEA到CHF组大鼠PVN中可导致MAP、HR、RSNA和血浆NE水平明显降低,而EF和FS明显升高(P<0.05),且注射20~25 min时各项指标开始出现变化,见图4。这些结果提示外源AEA可能通过抑制交感驱动而改善心衰大鼠心脏功能。
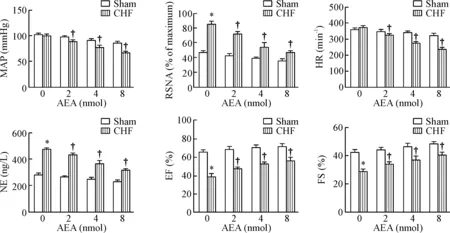
Figure 4.The effect of microinjection of AEA to PVN on the sympathetic drive and cardiac function indexes.Mean±SEM.n=6.*P<0.05vssham+vehicle (0 nmol AEA) group;†P<0.05vsCHF+vehicle (0 nmol AEA) group.
图4PVN内微量灌注AEA对交感驱动和心功能指标的影响
6 静脉注射AEA
与溶剂对照组相比,假手术组和CHF组大鼠静脉注射2倍于PVN内微量注射剂量的AEA对MAP、HR和RSNA均无明显影响(P>0.05,数据未在文中显示)。这一结果提示AEA产生的交感抑制作用主要是通过中枢下丘脑室旁核来完成的。
7 FAAH基因敲减对交感驱动和心功能指标的影响
与sham+Sc-shRNA组相比,CHF+Sc-shRNA组室旁核内FAAH蛋白表达量显著上调(P<0.05);然而,与CHF+Sc-shRNA组相比,CHF+FAAH-shRNA组室旁核内FAAH蛋白表达量显著下调(P<0.05),提示rAAV2-hSyn-FAAH-shRNA病毒可有效沉默PVN内FAAH基因,见图5。FAAH基因敲减(即微量注射病毒)4周后,检测CHF组及假手术组大鼠心功能及其交感驱动指标,结果显示敲减PVN内FAAH基因表达可显著改善CHF大鼠心功能(P<0.05),见表4,且CHF组大鼠交感驱动指标显著降低(P<0.05),见图6。
讨 论
本研究运用超声心动图、神经电生理、Western blot及高效液相色谱分析等技术,首次发现PVN内FAAH表达量显著上调,AEA生成量显著减少;在CHF大鼠模型PVN内微量注射AEA、PF3845及rAAV2-FAAHshRNA病毒后,CHF组交感驱动指标显著降低,心功能获得明显改善,研究结果提示抑制PVN内FAAH蛋白的表达可能通过增加AEA含量,从而抑制交感神经兴奋性。

Figure 5.The effects ofFAAHgene knockdown on the protein expression of FAAH in PVN.Mean±SEM.n=6.*P<0.05vssham+Sc-shRNA group;†P<0.05vsCHF+Sc-shRNA group.
图5PVN内敲减FAAH基因对FAAH蛋白表达的影响
表4敲减PVN内FAAH基因对大鼠心脏解剖、超声心动图和血流动力学指标的影响
Table 4.The effect of knockdown ofFAAHgene in PVN on the indexes of anatomy,hemodynamics and sympathetic drive in the CHF rats and the sham rats (Mean±SEM.n=6)

IndexSham+Sc⁃shRNASham+FAAH⁃shRNACHF+Sc⁃shRNACHF+FAAH⁃shRNABW(g)367±4359±4395±6397±6HW(g)1.38±0.041.36±0.051.69±0.03∗1.57±0.02†RV/BW(mg/g)0.71±0.110.63±0.281.57±0.21∗0.91±0.32†LW/BW(mg/g)6.21±0.316.17±0.2914.01±0.32∗9.12±0.53†IS(%)0034±3∗31±2LVEDV(mL)0.34±0.080.32±0.050.89±0.03∗0.64±0.04†LVEDD(mm)6.26±0.255.89±0.369.79±0.33∗7.41±0.43†EF(%)67.10±4.2473.10±5.8241.20±3.98∗60.30±3.51†FS(%)41.30±2.5742.20±0.8928.50±1.55∗34.20±2.31†LVSP(mmHg)118±3113±2102±4∗109±5†LVEDP(mmHg)3.5±2.12.8±1.121.7±3.8∗12.3±3.6†dp/dt(mmHg/s)7893±2147902±3194093±298∗4958±271†
*P<0.05vssham+Sc-shRNA group;†P<0.05vsCHF+Sc-shRNA group.
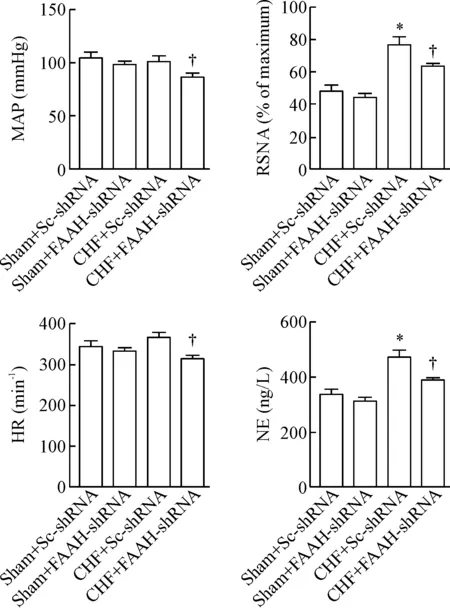
Figure 6.The effects ofFAAHgene knockout on sympathetic drive.Mean±SEM.n=6.*P<0.05vssham+Sc-shRNA group;†P<0.05vsCHF+Sc-shRNA group.
图6PVN内FAAH基因敲减对大鼠交感驱动指标的影响
在本次研究中,对假手术和CHF组大鼠双侧室旁核进行微量注射不同浓度的AEA和PF3845,实时监测假手术和CHF组大鼠的MAP、HR和RSNA的变化,结果显示在注射AEA和PF3845后,CHF组MAP、HR和RSNA和血浆NE水平均减少且心功能明显改善,可能机制为CHF状态下FAAH高表达,使得AEA生成量减少,交感驱动指标增强,心功能降低;相反,当抑制FAAH活性时,AEA含量增加,CHF原本增强的交感驱动受到的抑制作用较大,而假手术组大鼠PVN内内源性FAAH活性相对较低,故抑制PVN内FAAH活性或增加AEA浓度后,对假手术组大鼠交感驱动指标影响不如CHF组明显。选择冠脉结扎4周的大鼠,对CHF组与假手术组分别进行双侧PVN内rAAV2-FAAHshRNA病毒转染,结果显示病毒转染2周后,CHF组与假手术组的交感驱动指标均无明显变化,3周后交感驱动指标开始出现明显变化且随着时间的推移其变化越显著,因此本研究中我们选用了病毒转染4周的动物模型进行交感驱动指标的检测。
与本研究结果相矛盾的是,Niederhoffer等[15]曾报道在兔脑池中注入AEA,会增强交感神经兴奋性。但脑池中核团众多,具体是哪个核团发挥作用,尚不明确。且兔与鼠虽然同属小型哺乳动物,但其基因仍有很大差异。然而,有学者指出在孤束核中AEA能够通过延长压力反射时,从而抑制交感神经放电;另有研究报道显示,在麻醉大鼠PVN内注射AEA可导致动脉血压降低[14],该报道结果提示心血管自主调节神经核团内AEA发挥交感传出抑制作用,这与本研究结果相一致。
当然本研究中也存在着许多的不足,为此我们将其归纳为以下几点:(1)本研究结果虽然提示了CHF状态下FAAH可通过水解AEA激活交感传出神经兴奋性,但其具体通过哪条信号通路发挥的作用,目前尚不明确,这也是本课题组接下来的研究重点;(2)本研究虽然证实在PVN内AEA的浓度变化对交感神经兴奋性产生相应的影响,但也已有报道指出AEA不存储于突触泡中,而是“按需求”合成,是否有其它因子参与了AEA对交感神经兴奋性的调节,目前仍不明确。
综上所述,本研究结果提示,CHF状态下,PVN内FAAH蛋白表达量增加,可能进一步水解AEA,使AEA含量降低,进而导致了交感神经兴奋亢进。本研究所证实的CHF状态下PVN内FAAH上调介导CHF大鼠交感神经兴奋亢进这一假说,虽不能直接应用于临床,但为临床上CHF基因治疗提供了实验室理论依据,为治疗CHF疾病提供了新的靶点。
[参考文献]
[1]Fukuta H,Goto T,Wakami K,et al.Effects of catheter-based renal denervation on heart failure with reduced ejection fraction:a systematic review and meta-analysis[J].Heart Fail Rev,2017,22(6):657-664.
[2]马柳一,尹玉洁,位庚,等.慢性心衰大鼠下丘脑室旁核瞬时外向钾通道蛋白Kv4.2和Kv4.3低表达促进肾交感神经兴奋性[J].中国病理生理杂志,2016,32(3):522-526,533.
[3]李晓燕,刘金玲,桂乐,等.下丘脑室旁核小电导钙激活钾通道过表达降低慢性心衰大鼠肾交感神经兴奋性[J].中国病理生理杂志,2014,30(8):1478-1482.
[4]阳志晖,杨锐,陆阳.N-花生四烯酰乙醇胺及其类似物的神经保护作用[J].国际药学研究杂志,2010,37(3):209-212,216.
[5]Sticht MA,Limebeer CL,Rafla BR,et al.Endocannabinoid regulation of nausea is mediated by 2-arachidonoylglycerol (2-AG) in the rat visceral insular cortex[J].Neuropharmacology,2016,102:92-102.
[6]Tchantchou F,Tucker LB,Fu AH,et al.The fatty acid amide hydrolase inhibitor PF-3845 promotes neuronal survival,attenuates inflammation and improves functional recovery in mice with traumatic brain injury [J].Neuro-pharmacology,2014,85:427-439.
[7]Henry RJ,Kerr DM,Flannery LE,et al.Pharmacological inhibition of FAAH modulates TLR-induced neuroinflammation,but not sickness behaviour:an effect partially mediated by central TRPV1[J].Brain Behav Immun,2017,62:318-331.
[8]Baranowska-Kuczko M,MacLean MR,Kozlowska H,et al.Endothelium-dependent mechanisms of the vasodilatory effect of the endocannabinoid,anandamide,in the rat pulmonary artery[J].Pharmacol Res,2012,66(3):251-259.
[9]Lin YS,Lin RL,Bien MY,et al.Sensitization of capsaicin-sensitive lung vagal afferents by anandamide in rats:role of transient receptor potential vanilloid 1 receptors[J].J Appl Physiol,2009,106(4):1142-1152.
[10] Gomes-de-Souza L,Oliveira LA,Benini R,et al.Involvement of endocannabinoid neurotransmission in the bed nucleus of stria terminalis in cardiovascular responses to acute restraint stress in rats[J].Br J Pharmacol,2016,173(19):2833-2844.
[11] Yang K,Lei G,Xie YF,et al.Differential regulation of NMDAR and NMDAR-mediated metaplasticity by anandamide and 2-AG in the hippocampus[J].Hippocampus,2014,24(12):1601-1614.
[12] Batista PA,Fogaça MV,Guimarães FS.The endocannabinoid,endovanilloid and nitrergic systems could interact in the rat dorsolateral periaqueductal gray matter to control anxiety-like behaviors[J].Behav Brain Res,2015,293:182-188.
[13] Seagard JL,Dean C,Patel S,et al.Anandamide content and interaction of endocannabinoid/GABA modulatory effects in the NTS on baroreflex-evoked sympathoinhibition[J].Am J Physiol Heart Circ Physiol,2004,286(3):H992-H1000.
[14] Grzeda E,Schlicker E,Luczaj W,et al.Bi-directional CB1 receptor-mediated cardiovascular effects of cannabinoids in anaesthetized rats:role of the paraventricular nucleus[J].J Physiol Pharmacol,2015,66(3):343-353.
[15] Niederhoffer N,Szabo B.Cannabinoids cause central sympathoexcitation and bradycardia in rabbits[J].J Pharmacol Exp Ther,2000,294(2):707-713.
猜你喜欢
汽车实用技术(2022年7期)2022-04-20
粉末冶金技术(2021年3期)2021-07-28
房地产导刊(2020年11期)2020-12-28
铁道通信信号(2019年4期)2019-10-10
模具制造(2019年7期)2019-09-25
山东体育学院学报(2019年3期)2019-08-30
制造技术与机床(2019年7期)2019-07-22
首都体育学院学报(2018年6期)2018-11-30
中华诗词(2018年3期)2018-08-01
