
CaSO4和GA3对NaHCO3胁迫下二月兰种子及幼苗的影响1)
2018-03-27 07:59许凌欣严俊鑫邓雅楠胡鹏敏牟元鋆
东北林业大学学报 2018年3期
许凌欣 严俊鑫 邓雅楠 胡鹏敏 牟元鋆
(东北林业大学,哈尔滨,150040)
二月兰(OrychophragmusviolaceusL.)又名诸葛菜,为十字花科诸葛菜属1年生或2年生草本植物[1]。它返青早,披绿时期长,对生境条件要求不高,生命力旺盛[2],适宜在林下、林边、路边或草地成片栽植,且能与其他植物混种[3],自播能力强,毋需过多养护,是理想的园林地被植物。同时,二月兰为高亚油酸低芥酸种质,是优质的油料作物[4-6]。二月兰的茎叶水解提取物还具有药用价值[7-8]。因此,二月兰具有广泛的应用前景和经济价值。
土壤盐碱化是一个世界性生态问题。目前,对盐碱土壤进行改良和防治的措施有很多,其中最为经济有效的措施就是种植耐盐植物[9-10]。二月兰有一定的抗盐碱性,有望成为应用于盐碱地的优良植物。研究表明:CaSO4和GA3可以提高经济作物的抗盐能力,但CaSO4和GA3是否能提高观赏植物二月兰的耐碱能力还未证实。本试验研究CaSO4和GA3对NaHCO3胁迫下二月兰种子萌发及幼苗生长的影响,以期提高二月兰抗盐碱能力,为二月兰在盐碱地的栽培和应用提供参考。
1 材料与方法
1.1 供试材料
二月兰(Orychophragmusviolaceus)种子购买于世纪天缘生物科技有限公司千绿种业。净种后种子千粒质量为2.87 g,含水量为5.51%。试验于东北林业大学园林学院进行。
1.2 种子预处理
选取均匀、饱满、无病虫害的二月兰种子,用75%酒精浸泡消毒3 min,然后用蒸馏水冲洗4次,再用蒸馏水浸泡2 h后用无菌滤纸吸干待用。
1.3 试验设计
用NaHCO3溶液模拟碱胁迫,参照前期预试验结果,本试验设置了T1(25 mmol·L-1)、T2(50 mmol·L-1)、T3(75 mmol·L-1)的NaHCO3溶液;CaSO4设置3、6、9、12、15 g·L-1共5个质量浓度梯度;GA3设置0.1、0.2、0.3、0.4、0.5 g·L-1共5个质量浓度梯度;以蒸馏水为对照。采用培养皿滤纸发芽法,将三层滤纸置于Φ=10 cm培养皿中,分别加入上述溶液后,再将处理好的种子均匀置于其中,每皿50粒,每种处理设置3个重复,置于光照培养箱中培养,培养条件为光照强度1 000 lx,湿度50%,温度25 ℃,每天14 h光照和10 h黑暗。每24 h用称质量法补充蒸馏水以维持一定的溶液浓度和一定的湿度。以胚根突破种皮作为发芽标准,从种子置皿之日起每天14:00观察和记录种子的萌发及生长情况,连续3 d没有新种子萌发时结束试验,本试验培养时间为12 d。
1.4 测定指标及方法
1.4.1 种子萌发指标
发芽率(GR)=(萌发种子数/供试种子数)×100%。
发芽势(GE)=(规定时间内发芽种子数/供试种子数)×100%。
发芽指数(GI)=∑(Gt/Dt);Gt为当天发芽数,Dt为相应的发芽时间。
活力指数(VI)=S×发芽指数(GI);S为10株幼苗平均单株干质量,单位为mg[11]。
1.4.2 幼苗生长指标
发芽结束期后,测定胚根胚轴长度和干质量。
胚根胚轴长度测定:用刻度尺测量幼苗的胚轴和胚根长度,每种处理分别随机取5株幼苗(幼苗少于5株的组,取所有的幼苗),每组3次重复。
干质量测定:测定株高后,每种处理分别随机取5株整株幼苗(幼苗少于5株的组,取所有的幼苗),用滤纸吸干其表面水分,置于烘箱中烘干至恒质量,称量其干质量,每组重复3次[12]。
1.5 数据处理
数据分析统计采用SPSS 22.0数据统计软件,制图采用WPS Office软件。
2 结果与分析
2.1 CaSO4和GA3对碱胁迫下二月兰种子萌发的影响
由表1可见,在T1胁迫下,3、6、9、12 g·L-1的CaSO4处理可以促进种子的萌发,其整体的发芽指标均高于对照。且随着CaSO4质量浓度的升高,促进效果呈先上升后下降的趋势。其中以9 g·L-1的CaSO4处理效果最佳,二月兰种子的发芽率、发芽势、发芽指数、活力指数分别较对照提高了13.34%、32.01%、26.19%、49.80%,差异显著(P<0.05)。当CaSO4的质量浓度达到15 g·L-1时会抑制二月兰种子的萌发。0.1、0.2、0.3、0.4 g·L-1的GA3处理会促进二月兰种子的萌发。随着GA3质量浓度的增加,其促进效果先上升后下降,且有两个峰值。总体以0.3 g·L-1的GA3处理效果最佳,二月兰种子的发芽率、发芽势、发芽指数、活力指数分别较对照提高了41.34%、36.00%、25.51%、54.58%,差异显著(P<0.05)。当GA3质量浓度达到0.5 g·L-1时不能促进二月兰种子的萌发。在T1胁迫下,经GA3处理的二月兰种子发芽率、发芽势、发芽指数、活力指数均高于经CaSO4处理的二月兰种子,GA3对二月兰种子萌发的促进作用效果明显优于CaSO4对其的促进作用。
由表1可见,在T2胁迫下,3、6、9、12 g·L-1的CaSO4处理均可促进二月兰种子萌发。其中以9 g·L-1的CaSO4处理效果最佳,二月兰种子的发芽率、发芽势、发芽指数、活力指数分别较对照提高了17.65%、56.25%、53.94%、148.86%,差异显著(P<0.05)。当CaSO4的质量浓度达到15 g·L-1时会抑制二月兰种子的萌发。0.1、0.2、0.3、0.4 g·L-1的赤霉素处理均可促进二月兰种子萌发,其整体发芽指标均高于对照。其中以0.3 g·L-1的赤霉素处理效果最佳,二月兰种子的发芽率、发芽势、发芽指数、活力指数分别较对照提高了50.01%、39.59%、58.79%、176.14%,差异显著(P<0.05)。当赤霉素质量浓度达到0.5 g·L-1时,对二月兰种子萌发影响不大。在T2胁迫下,GA3处理的二月兰种子发芽率、发芽势、发芽指数、活力指数均高于CaSO4处理的二月兰种子,GA3对二月兰种子萌发的促进作用效果更好。
由表1可见,在T3胁迫下,CaSO4处理可显著促进二月兰种子萌发,其整体的发芽指标均高于对照。且以12 g·L-1的硫酸钙处理效果最佳,二月兰种子的发芽率、发芽势、发芽指数、活力指数分别较对照提高了272.25%、477.83%、353.49%、779.25%,差异显著(P<0.05)。0.1、0.2、0.3 g·L-1的赤霉素处理均可促进二月兰种子萌发,其整体发芽指标均高于对照。其中以0.3 g·L-1的赤霉素处理效果最佳,二月兰种子的发芽率、发芽势、发芽指数、活力指数分别较对照提高了138.01%、200.00%、167.44%、486.79%,差异显著(P<0.05)。当赤霉素质量浓度达到0.4 g·L-1时二月兰种子几乎不能萌发。在T3胁迫下,GA3处理的二月兰种子发芽率、发芽势、发芽指数、活力指数均低于CaSO4处理的二月兰种子,CaSO4对二月兰种子萌发的促进作用效果更好。
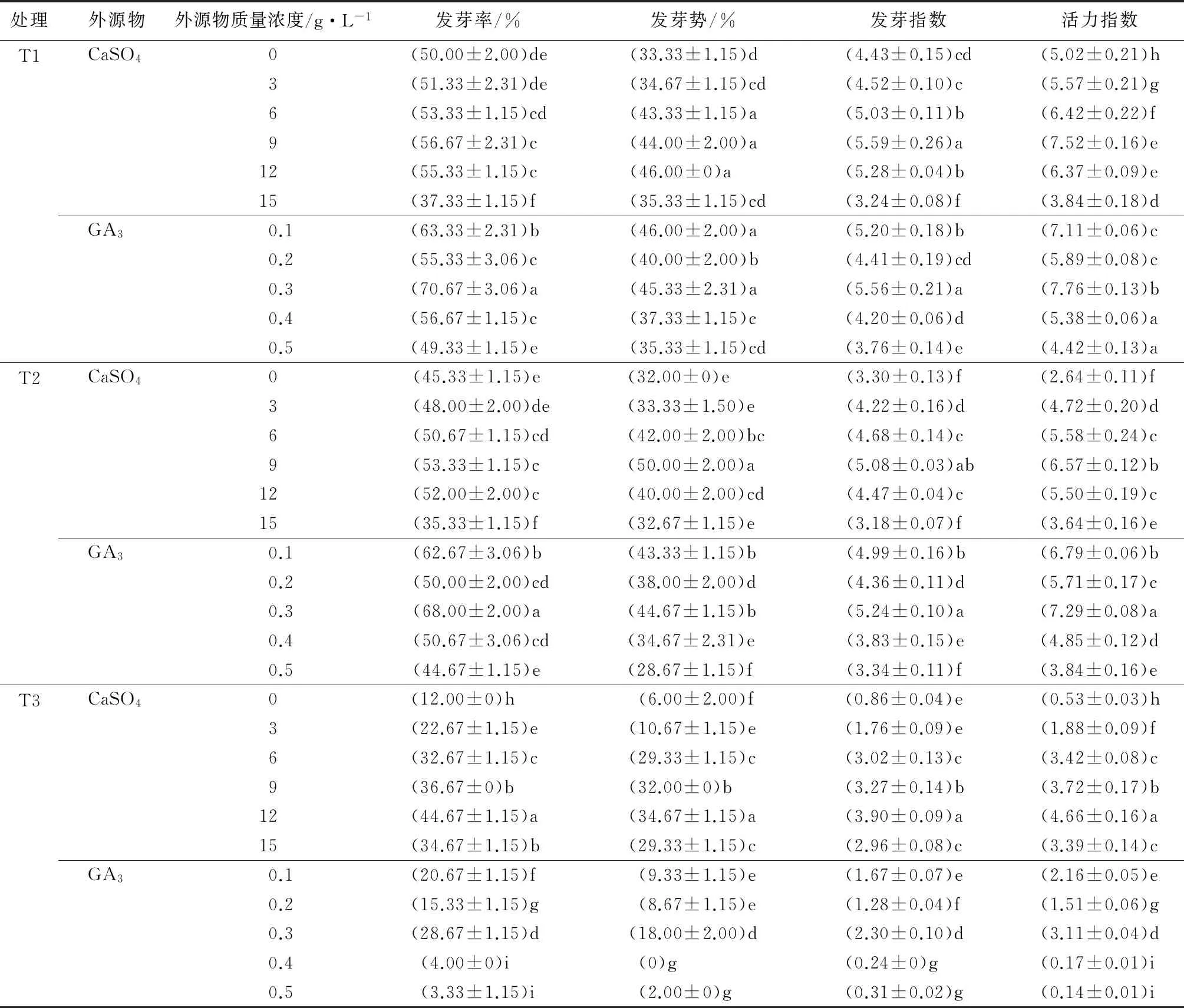
表1 NaHCO3胁迫下不同质量浓度CaSO4、GA3对二月兰种子萌发的影响
注:表中数据为平均值±标准差;同列不同字母表示各指标在同一处理同一外源物不同质量浓度条件下差异性显著(P<0.05)。
结果表明,在3种胁迫下,适宜的CaSO4和GA3处理均可显著促进二月兰种子的萌发,且不同质量浓度的CaSO4和GA3处理对二月兰种子的萌发影响不同。随着CaSO4质量浓度的增加,二月兰种子的发芽率、发芽势、发芽指数和活力指数都呈先上升后下降的趋势。而随着GA3质量浓度的增加,二月兰种子的发芽率、发芽势、发芽指数和活力指数的变化为先上升再下降,再上升后下降的双峰趋势,并在0.3 g·L-1时达到最佳效果。在T1、T2胁迫下,GA3处理可显著促进二月兰种子的萌发,且促进效果优于CaSO4处理。而在T3胁迫下,GA3处理几乎不能促进二月兰种子的萌发,而经CaSO4处理的促进效果明显。
2.2 CaSO4和GA3对碱胁迫下二月兰幼苗生长的影响
由表2可见,在T1胁迫下,6、9、12、15 g·L-1的CaSO4处理可以促进二月兰幼苗的生长。其中以9 g·L-1的CaSO4处理效果最佳,二月兰幼苗的胚根长度、胚轴长度、干质量分别较对照提高了43.20%、162.83%、19.47%,差异显著(P<0.05)。0.1、0.2、0.3、0.4 g·L-1的GA3会促进二月兰幼苗的生长。其中0.2 g·L-1的GA3对二月兰胚轴生长处理效果最佳,较对照提高220.33%,差异显著(P<0.05);0.3 g·L-1的GA3对二月兰胚根生长和干质量增加的处理效果最佳,分别较对照提高56.07%、23.89%,差异显著(P<0.05)。在T1胁迫下,GA3处理的二月兰幼苗比CaSO4处理的二月兰幼苗胚根、胚轴的增长量更大,干质量积累更多,在此条件下GA3对二月兰幼苗生长的促进作用效果更好。
由表2可见,在T2胁迫下,CaSO4处理可以促进二月兰幼苗的生长,其整体的形态指标均高于对照。其中以9 g·L-1的CaSO4处理效果最佳,二月兰幼苗的胚根长度、胚轴长度、干质量分别较对照提高了88.56%、139.58%、61.25%,差异显著(P<0.05)。0.1、0.2、0.3 g·L-1的GA3处理能促进二月兰幼苗的生长。其中0.2 g·L-1的GA3对二月兰胚轴生长处理效果最佳,较对照提高138.33%,差异显著(P<0.05);0.3 g·L-1的GA3对二月兰胚根生长和干质量增加的处理效果最佳,分别较对照提高103.00%、73.75%,差异显著(P<0.05)。在T2胁迫下,GA3处理比CaSO4处理对二月兰幼苗生长的促进作用效果更好。
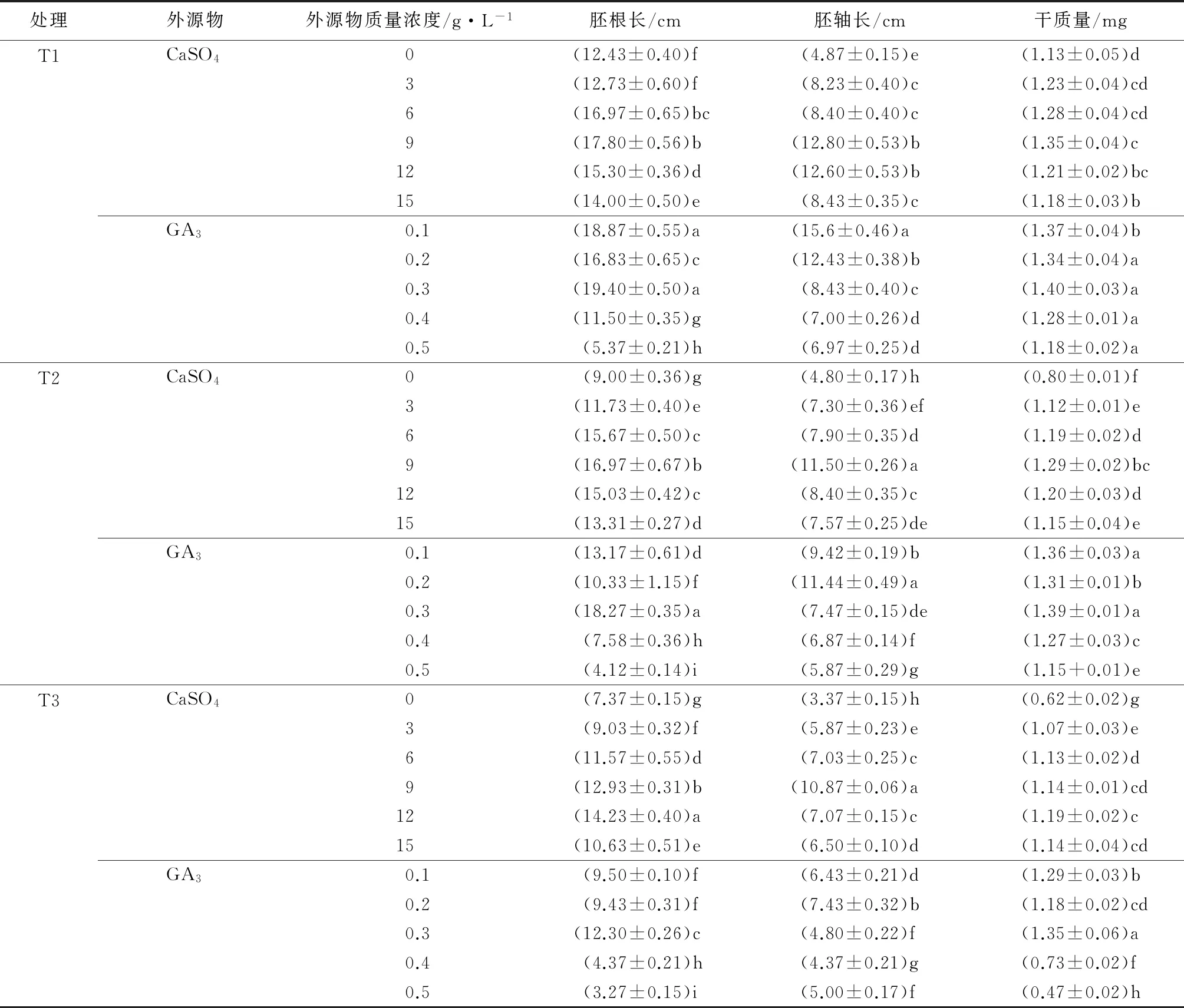
表2 NaHCO3胁迫下不同质量浓度CaSO4、GA3对二月兰幼苗生长的影响
注:表中数据为平均值±标准差;同列不同字母表示各指标在同一处理同一外源物不同质量浓度条件下差异性显著(P<0.05)。
由表2可见,在T3胁迫下,CaSO4处理可以增长二月兰幼苗的胚根长度、胚轴长度和干质量。12 g·L-1的硫酸钙处理对二月兰胚根长度和干质量处理效果最佳,分别较对照提高了93.08%、91.94%,差异显著(P<0.05)。0.1、0.2、0.3 g·L-1的GA3处理能促进二月兰幼苗的生长。其中0.2 g·L-1的GA3对二月兰胚轴生长处理效果最佳,较对照提高120.47%,差异显著(P<0.05);0.3 g·L-1的赤霉素对二月兰胚根生长和干质量增加的处理效果最佳,分别较对照提高66.89%、117.74%,差异显著(P<0.05)。在T3胁迫下,经GA3处理的比CaSO4处理的对二月兰幼苗生长的促进作用效果更好。
结果表明,在3种碱胁迫下,适宜的CaSO4和GA3处理均可显著促进二月兰幼苗的生长。在3种胁迫下,二月兰幼苗胚根、胚轴长度及干质量随CaSO4质量浓度的增加呈先上升后下降的趋势。二月兰幼苗胚根、胚轴长度及干质量随GA3质量浓度的增加呈先上升再下降,且有两个峰值。GA3质量浓度为0.3 g·L-1时对胚根和干质量的促进效果最佳,而0.2 g·L-1时对胚轴的促进效果最佳。随着NaHCO3浓度的上升,CaSO4处理的最佳质量浓度也上升。而GA3处理的最佳质量浓度不随NaHCO3浓度的改变而变化。在T1、T2、T3胁迫下,GA3处理对二月兰幼苗生长的促进效果与CaSO4处理的效果差别不大,但GA3处理对二月兰幼苗生长的促进作用更明显。
3 结论与讨论
植物是否能在所处环境中生存受到许多因素的影响,植物种子能否萌发是其中重要的影响因素。碱性环境会抑制种子萌发和幼苗的生长,严重影响植物的生存。本试验研究了CaSO4和GA3对碱胁迫下二月兰种子萌发和幼苗生长的影响。毛桂莲等[13]对油葵的研究结果表明,加入CaSO4使溶液的pH值发生了改变,减轻了盐碱胁迫对油葵萌发和生长的抑制作用。而且Ca2+作为第二信使对受盐碱胁迫的植物有重要的作用,它可以增加膜结合Ca2+量,从而提高逆境下膜的稳定性。同时CaSO4作为硫肥对植物生长也有促进作用[14]。阿依夏木等[14]对甘草种子的研究结果表明,加入CaSO4可显著促进甘草的萌发和生长。本试验中,在NaHCO3胁迫条件下,加入CaSO4可促进二月兰种子的萌发和二月兰幼苗的生长。发芽率、发芽势、发芽指数、活力指数较对照上升,胚根长度、胚轴长度、干质量较对照增多。可能是因为CaSO4中和了NaHCO3的碱性,降低了pH值,引入的Ca2+提高膜的稳定性,从而减轻了伤害。这与毛桂莲等[13]对油葵种子和阿依夏木等[14]对甘草种子的研究结果一致。本试验中,在相同胁迫下,随着CaSO4质量浓度的升高,种子的活力呈先上升后下降的趋势。但当加入过多的CaSO4时不仅不能促进二月兰萌发和幼苗的生长,而且还会抑制其萌发和生长。可能是因为当加入过多的CaSO4时,悬浊液中未溶解的CaSO4附着在种子表面,阻碍了种子对水分的吸收和种子的呼吸作用,从而抑制了种子的萌发和幼苗生长。但是其影响种子萌发和幼苗生长的作用,以及对碱胁迫的缓解机制还需要进一步研究。
高春智等[15]的研究表明,GA3浸种可以促进樟子松种子萌发。徐芬芬等[16]、张韵等[17]的研究表明,GA3浸种可以缓解盐碱胁迫对小白菜[16]、黄瓜[17]等种子的萌发抑制。本试验结果表明,在NaHCO3胁迫条件下,加入GA3能促进二月兰种子的萌发和幼苗的生长。与高春智等[15]徐芬芬等[16]、张韵等[17]研究结果一致。可能是因为GA3可以调节种子体内激素平衡。还有研究表明,GA3能修复细胞膜完整性,减少营养物质外渗。本试验结果显示,相同质量浓度的GA3处理对二月兰胚根和胚轴的影响效果不同。这可能是因为胚根和胚轴对GA3的敏感程度不同。高春智等[15]、徐芬芬等[16]、张韵等[17]的研究结果表明,最佳的GA3处理质量浓度不同。本试验结果表明,0.3 g·L-1的GA3对二月兰种子萌发和生长的促进效果最明显。这与高春智等[15]、徐芬芬等[16]、张韵等[17]的结果不同。可能是因为GA3处理的方法不同导致结果不同。且不同植物对GA3的敏感程度不同也是产生差异的原因。虽然CaSO4和GA3能够促进盐碱胁迫下的植物萌发和生长。但是其影响种子活力的生理、生化机制和分子机理和对碱胁迫的缓解机制还需要进一步研究。
综上所述,适宜质量浓度的CaSO4和GA3对NaHCO3胁迫下二月兰种子萌发及幼苗生长有促进作用。可以为二月兰在盐碱地的应用提供参考。本研究还具有一定的局限性,单一的碱胁迫并不能代表自然状态下的盐碱胁迫,有必要进行盐碱混合交叉胁迫处理下二月兰种子和幼苗的生理理化性质的研究。
[1] 李悦.诸葛菜配子与合子染色体加倍及其相关研究[D].北京:北京林业大学,2008.
[2] 史滟滪,梁发辉,冀媛媛,等.二月兰在园林中的应用价值[J].天津农林科技,2011(2):18-20.
[3] 桑晓东.优良地被植物二月兰在吉林省的新发现[J].辽宁林业科技,2014(6):47-48.
[4] 张兆清,罗鹏,李旭锋.四川野生十字花科油料植物资源研究[J].西南农业学报,1990,3(3):19-22.
[5] 罗鹏,兰泽蘧,黄建,等.珍贵油料植物诸葛菜的研究[J].自然资源学报,1991,6(3):206-210.
[6] 孙小芹,庞慧,郭建林,等.十字花科58属94种野生植物种子脂肪酸组分分析[J].林产化学与工业,2011,31(6):46-54.
[7] 王欣,李向,王红庆.诸葛菜中黄酮类化合物提取方法和抗氧化性研究[J].安徽农业科学,2007,35(27):8667-8669.
[8] 王欣,夏新奎,徐固华.微波辅助萃取诸葛菜中的总黄酮研究[J].安徽农业科学,2007,35(22):6705-6706.
[9] 赵秋,高贤彪,吴迪,等.越冬绿肥二月兰耐盐能力及在盐碱耕地上的培肥效果[J].中国土壤与肥料,2010(4):65-68.
[10] 谷洪彪,姜纪沂.土壤盐碱化的灾害学定义及其风险评价体系[J].灾害学,2013,28(1):23-27.
[11] 张志良,瞿伟菁.植物生理学实验指导[M].北京:高等教育出版社,2003.
[12] 王玉萍,王映霞,白向利,等.硅对NaCl胁迫下甜瓜种子萌发及幼苗生长的影响[J].草业学报,2015,24(5):108-116.
[13] 毛桂莲,马春燕,谢亚军,等.CaSO4对盐碱胁迫下油葵种子萌发的影响[J].农业科学研究,2009,30(3):21-23,43.
[14] 阿依夏木·沙吾尔,吐尔逊·吐尔洪,贾宏涛,等.硫肥类型与施用量对甘草种子发芽的影响[J].中药材,2016,39(6):1224-1229.
[15] 高春智,何炎红,田有亮,等.不同浓度赤霉素浸种对樟子松种子萌发的影响[J].内蒙古农业大学学报(自然科学版),2012,33(3):67-72.
[16] 徐芬芬,李田.外源赤霉素GA3对混合盐碱胁迫下小白菜种子萌发的影响[J].生物加工过程,2012,10(6):56-59.
[17] 张韵,郁继华,钟新榕,等.外源ABA和GA3对NaCl胁迫下黄瓜种子萌发特性的影响[J].甘肃农业大学学报,2006,41(2):27-30.
猜你喜欢
热带海洋学报(2022年6期)2022-12-05
广西林业科学(2021年6期)2022-01-21
山西农业科学(2021年8期)2021-08-17
现代园艺(2020年19期)2020-10-02
山西农业科学(2020年1期)2020-02-27
中国稻米(2019年6期)2019-12-05
种子(2019年6期)2019-07-13
种子(2019年4期)2019-05-28
现代农村科技(2019年4期)2019-01-06
上海农业学报(2017年4期)2017-04-10
