
库源调节对油茶叶片碳水化合物含量及其酶活性的影响
2018-03-27 03:07彭映赫陈永忠许彦明陈隆升彭邵锋
西南林业大学学报 2018年1期
彭映赫 陈永忠 许彦明 王 瑞 陈隆升 马 力 彭邵锋
(1. 湖南省林业科学院,湖南 长沙 410004;2. 国家油茶工程技术研究中心,湖南 长沙 410004)
库源关系改变可通过影响卡尔文循环、PSII反应中心光化学效率、叶片气孔闭合等过程调节作物的光合作用及其产物的运输和分配[1-2]。研究表明,减源增库可提高作物光合能力,增加碳代谢能力[3]。生产中,摘叶、疏花疏果等措施都会影响库源关系,改变光合同化产物在库源间的分配,从而影响作物产量和品质[2-5]。源活性可用光合末端产物碳水化合物衡量,库源关系中源叶碳水化合物含量变化受其代谢相关酶活性调控,因此,它们的动态变化及相互关系是研究库源关系的重要内容之一,对阐明库源关系中光合末端产物对源叶光合作用的调控具有重要价值。
经济林木生长周期长,类型丰富,但库源关系的研究相对滞后。近年来,随着林木栽培生理的发展和生产中精细化栽培需求的增长,经济林木库源关系成为研究热点。油茶 (Camelliaoleifera) 为山茶科山茶属常绿灌木或小乔木,是重要的木本食用油料树种,全国栽培总面积达3.67 × 106hm2,年产茶油4.5 × 108kg。目前,烟草 (Nicotianatabacum)[6]、水稻 (Oryzasativa)[7-8]、玉米 (Zeamays)[9]、西红柿 (Lycopersiconesculentum)[10]、棉花 (Gossypiumspp.)[11-12]等农作物的库源关系方面已有大量报道,但针对油茶的研究较少,尤其碳代谢相关酶活性变化方面几乎处于空白。已有研究表明,在环剥枝条上,通过调控叶果比来研究库源关系是可行的[13]。因此,本试验以5年生油茶为试材,通过调控叶果比来研究库源关系改变对源叶片可溶性糖和淀粉含量及其相关酶活性的变化。
1 材料与方法
1.1 材料与处理
试验于2015年8月在湖南省林业科学院试验林场进行。试验材料为5年生油茶,株行距2 m × 3 m。试验选用立地条件相似、长势基本一致的油茶树6株,在其果实膨大期间,每株选取树冠外围南面长势基本相同的当年生枝条,在非人为去果情况下,选择自然生长的不同叶果比营养枝,分别为T1(5叶,即无果),T2(3叶1果枝),T3(4叶1果枝),T4(5叶1果枝),T5(5叶2果枝)。同一株树相同处理均为12枝,同时对试验枝基部进行环剥,通过环剥实现对库源关系进行控制,阻断试验枝与树体其他部分之间碳水化合物的流动。
于2015年8月21日对所有处理的叶片全部取样,每2株树相同处理叶片混为一个样品,共计3份样品,样品放入冰盒迅速带回实验室,用去离子水清洗擦干,其中每个处理的叶片选择一部分80 ℃烘干,用于碳水化合物含量的测定,剩余的放于-70 ℃冰箱中保存用于酶活性的测定。
1.2 测定指标与方法
总可溶性糖采用乙醇提取,蒽酮比色法[14]测定;蔗糖采用间苯二酚法[14]测定;果糖采用间苯二酚法[14]测定;淀粉采用高氯酸提取,用蒽酮比色法[14]测定;蔗糖合成酶、蔗糖磷酸合成酶活性采用间苯二酚法[15]测定;酸性转化酶、碱性转化酶活性采用3, 5-二硝基水杨酸比色法[15]测定;淀粉磷酸化酶活性采用无机磷比色法[15]测定。室内实验测定均为3次重复。
试验数据采用Excel 2007及SPSS 19.0进行统计和相关分析。
2 结果与分析
2.1 油茶叶片可溶性糖和淀粉含量变化
由图1可知,油茶叶片的可溶性总糖和蔗糖含量随叶果比的变化呈先下降后上升的趋势,不同处理组之间差异未达显著水平。去叶减源处理 (T2) 可溶性总糖和蔗糖含量最低,分别比T1低6.63%、11.43%。增源处理 (T3、T4) 叶片可溶性糖及淀粉含量分别较T2增加了0.05%、12.8%和1.43%、10.12%。增库处理 (T5) 使库的需求相对增加,植株体内更多的光合产物流向果实库。同处理间,果糖和淀粉含量差异明显 (P< 0.05)。其中,果糖与可溶性总糖含量变化趋势一致,当留叶数为4 (T3) 时,果糖含量最低,分别较其他各处理组降低了5.83%、3.11%、15.55%和8.6%。T4叶片果糖含量最高,为15.48 mg/kg,增库处理后,果糖含量随之下降。无果处理组 (T1) 叶片淀粉含量达到最大值,为106.72 mg/kg,叶果比减小 (T2,T5),叶片淀粉积累量下降,并显著低于其他各处理 (P< 0.05)。对于不同叶果比处理,叶片可溶性糖和淀粉含量与其留叶数目呈正相关关系。
2.2 油茶叶片碳水化合物代谢相关酶活性的变化
与碳水化合物代谢相关酶的活性变化见图2,不同处理组间相关酶活性均存在显著性差异 (P< 0.05),无果处理组 (T1) 中转化酶 (AI)、蔗糖合成酶 (SS)、淀粉磷酸化酶 (SP) 活性显著降低 (P< 0.05),增源处理后 (T2~T5),酶的活性逐渐增加。其中SS、蔗糖磷酸合成酶 (SPS) 呈单峰曲线类型,T3、T4达到最大值,增库处理 (T5)其酶活性显著下降,分别低于最大值34.68%、17.96%;增源扩库处理后 (T2~T5),AI和SP活性逐渐增加,在T5出现最大值。除 T1外,各组酶活性存在显著性差异 (P< 0.05)。
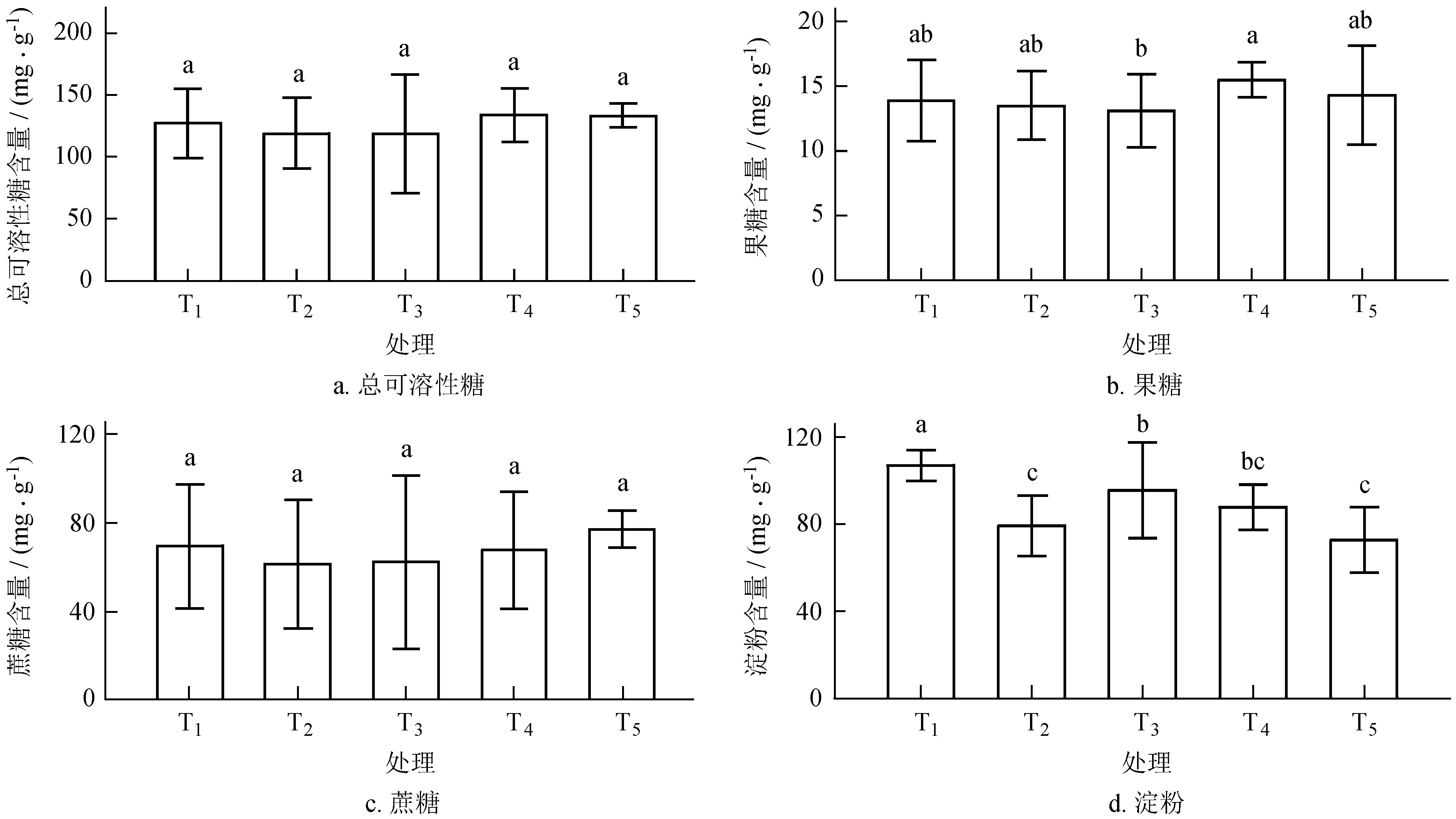
图1不同库源比条件下叶片碳水化合物含量变化
Fig.1 Variation of carbohydrate content in leaves of different sink-source ratio
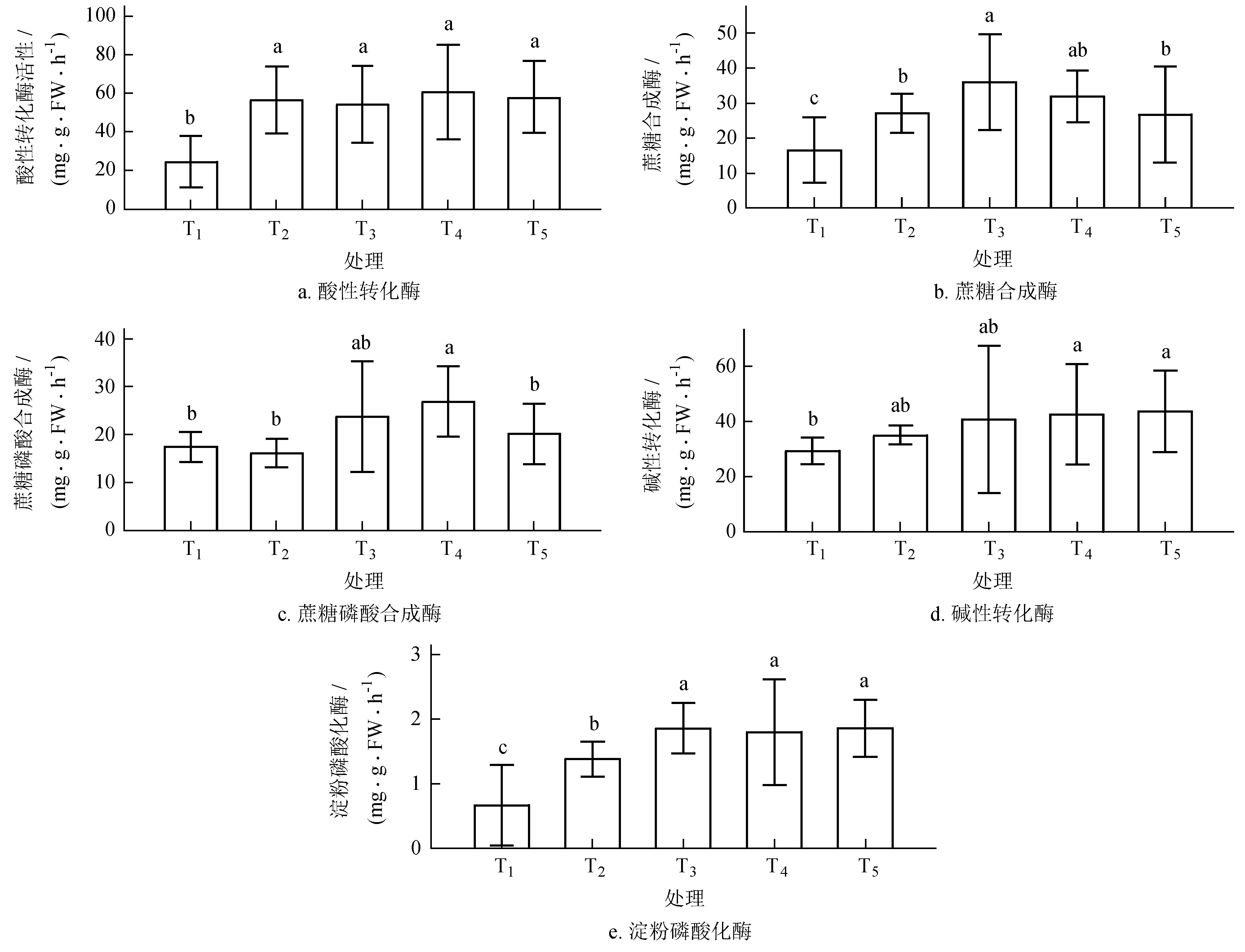
图2不同库源比条件下叶片碳水化合物代谢相关酶活性变化
Fig.2 Variation of related enzymes activities to carbohydrate metabolism in leaves of different sink-source ratio
2.3 油茶叶片碳水化合物含量与其代谢相关酶活性的相关性分析
可溶性糖及淀粉含量与其代谢相关酶活性之间的相关性分析见表1。结果表明,可溶性总糖含量与蔗糖、果糖含量呈显著正相关 (P< 0.05),r分别为0.634、0.612,而与SS、SPS、AI相关性不明显。蔗糖含量与其代谢酶活性间没有明显的相关性,果糖含量与其均呈正相关关系,但相关性不显著。淀粉含量与AI、淀粉磷酸合成酶成极显著负相关 (P< 0.01),r分别为-0.669、-0.517。各代谢酶之间也存在明显相关性,SS与酸性转化酶、碱性转化酶、淀粉磷酸化酶呈显著正相关,r分别为0.674、0.567和0.734,同时,AI与淀粉磷酸化酶亦呈极显著正相关 (P< 0.01),r为0.815、0.687。

表1 叶片碳水化合物与代谢相关酶活性的相关性Table 1 The correlation between carbohydrate content and its related enzymes activities in leaves
注:*表示差异显著,**表示差异极显著。
3 结论与讨论
库源关系改变是提高光合效率和光合产物总量、调整同化物运输与分配的重要途径[16],并可对经济林木树种的经济产量及品质产生影响[1]。叶片光合产物需输出以满足库生长发育的需要,而果实是油茶重要的库[17]。本试验中,无果处理组(T1)库强低,叶片可溶性糖和淀粉含量显著增加,但与其代谢相关的酶活性显著低于其他各组,在库相同的条件下(T2、T3、T4),留叶数增加,其可溶性糖和淀粉含量亦有所升高,该结果也印证了“反馈抑制假说”[18-19]。但也有研究表明,去果降低库强导致的可溶性糖和淀粉的积累并不导致光合末端产物合成的关键酶活性的降低[20-22]。越来越多的研究结果认为,潜在酶活性并不因为末端产物在叶片中的积累而受影响,低库强下叶光合作用调节一定存在反馈抑制以外的其他调节途径。因此,油茶库源关系调节对其碳水化合物代谢、光合作用调控的研究还需进一步对长期响应过程中叶片气体交换。叶绿素荧光、光合产物变化特征及各项参数间的相互关系上展开。
前人研究表明,油茶果实发育所需的营养物质主要是由临近的叶片提供[23],减源处理在短期内可提高留存叶片的光合效率,增加叶片可溶性糖及淀粉的积累量,但后期留存叶片光合能力的下降可造成源对库供给的同化物总量减少,引起果实品质产量下降。本试验中,叶片中可溶性糖和淀粉含量对库源比的降低而显著升高,但随处理时间的延长,缩库处理是否会影响库源间同化物的供给,并进一步影响果实品质及产量,因此,更有利于提高果实单果重和出籽率、促进茶油产量上升的最优叶果比还需进一步的研究。
[1] 彭丽丽, 姜卫兵, 韩健. 源库关系变化对果树产量及果实品质的影响[J]. 经济林研究, 2012, 30(3): 134-140.
[2] 程杰山, 王利军, 蒋爱丽, 等. 果树库源关系改变对原液光合作用的影响机制研究进展[J]. 中国农学通报, 2014, 30(19): 75-80.
[3] 朱振家, 姜成英, 史艳虎, 等. 库源比改变对油橄榄产量及源叶光合作用的调节[J]. 中国农业科学, 2015, 48(3): 546-554.
[4] Iqbal N, Masood A, Khan N A. Analyzing the significance of defoliation in growth, photosynthetic compensation and source-sink relations[J]. Photosynthetica, 2012, 50(2): 161-170.
[5] Plaut Z, Mayoral M L, Reinhold. Effect of altered sink:source ratio on Photosynthetic metabolism of source leaves[J]. Plant Physiology, 1987, 85: 786-791.
[6] Werner T, Holst K, Pōrs Y, et al. Cytokinin deficiency causes distinct changes of sink and source parameters in tobacco shoots and roots[J]. Journal of Experimental Botany, 2008, 59(10): 2659-2672.
[7] 李木英, 石庆华, 曾梅花, 等. 杂交稻主季库源性状对再生稻发育的影响[J]. 中国农学通报, 2009, 25(23): 175-181.
[8] 朱永川, 熊洪, 张林, 等. 杂交中稻齐穗期下部叶片对结实率的影响与组合间库源结构的关系[J]. 西南农业学报, 2009, 5(22): 1225-1231.
[9] 戴明宏, 赵久然, 杨国航, 等. 不同生态区和不同品种玉米的源库关系及碳氮代谢[J]. 中国农业科学, 2011, 44(8): 1585-1595.
[10] Xiao S G, Zhou P H, Heuvelink E, et al. Simulation analysis of the effects of dry matter production and partitioning in greenhouse tomato synchronous leaf pruning[J]. Scientia Agricultura Sinica, 2006, 39(10): 2154-2158.
[11] 汪玉磊, 杨劲松. 冬小麦群体库源比及其与产量的关系[J]. 土壤通报, 2009, 40(1): 140-141.
[12] 孙红春, 李存东, 张月辰, 等. 棉花源库比对中、下部果枝叶生理活性及铃重的影响[J]. 作物学报, 2008, 34(8): 1459-1463.
[13] Watari J, Kobae Y, Yamaki S, et al. Identification of sorbitol transporters expressed in the phloemof apple source leaves[J]. Plant Cell Physiol, 2004, 45(8): 1032-1041.
[14] 张志良, 瞿伟菁. 植物生理学实验指导 [M]. 北京: 高等教育出版社, 2003.
[15] 高俊凤. 植物生理学实验指导[M]. 北京: 高等教育出版社, 2006.
[16] Nebauer S G, Renau-Morata B, Guardiola J L, et al. Photosynthesis down-regulation precedes carbohydrate accumulation under sink limitation in Citrus[J]. Tree Physiol, 2011, 31: 169-177.
[17] 何方, 李纪元. 油茶产量与其相关因素的研究[J]. 经济林研究, 1990, 8(1): 1-13.
[18] Layne D R, Flore J A. End-product inhibition of photosynthesis inPrunuscerasusL. in response to whole-plant source-sink manipulation[J]. Journal of the American Society for Horticultural Science, 1995, 120(4): 583-599.
[19] Jeannette E, Reyss A, Grégory N, et al. Carbohydrate metabolism in a heat-girdled maize source leaf[J]. Plant Cell and Environment, 2000, 23(1): 61-69.
[20] Sawada S, Kuninaka M, Watanabe K, et al. The mechanism to suppress photosynthesis through end-product inhibition in single-rooted soybean leaves during acclimation to CO2enrichment[J]. Plant and Cell Physiology, 2001, 42(10): 1093-1102.
[21] Zhou R, Quebedeaux B. Changes in photosynthesis and carbohydrate metabolism in mature apple leaves in response to whole plant source-sink manipulation[J]. Journal of the American Society for Horticultural Science, 2003, 128(1): 113-119.
[22] 李卫东, 吴本宏, 李绍华, 等. 去果对桃源叶碳水化合物含量及其相关酶活性日变化的影响[J]. 园艺学报, 2006, 33(3): 605-608.
[23] 袁军, 石斌, 吴泽龙, 等. 不同库源关系对油茶光合作用及果实品质的影响[J]. 植物生理学报, 2015, 51(8): 1287-1292.
猜你喜欢
少儿科技(2021年8期)2021-01-02
婚姻与家庭·性情读本(2019年11期)2019-11-18
食品界(2019年8期)2019-10-07
保健与生活(2019年7期)2019-07-31
现代园艺(2017年13期)2018-01-19
现代园艺(2017年21期)2018-01-03
现代园艺(2017年21期)2018-01-03
中国西部(2017年4期)2017-04-26
中成药(2016年4期)2016-05-17
烟草科技(2015年8期)2015-12-20
