
外源硒对镉胁迫下菜心Fe、Mn、Cu、Zn吸收与转运的影响
2018-03-26 10:47吴志超赵亚荣张卫杰王富华
农业环境科学学报 2018年3期
刘 帅 ,吴志超 ,赵亚荣 ,张卫杰 ,王富华 *
(1.广东省农业科学院农产品公共监测中心,广州 510640;2.农业部农产品质量安全检测与评价重点实验室,广州 510640;3.农业部农产品质量安全风险评估实验室,广州 510640)
镉(Cd)是一种生物毒性较强的重金属元素,是国际劳动卫生重金属委员会和联合国环境规划署认定的重点污染物[1]。2014年《全国土壤污染状况调查公报》显示,我国土壤环境质量堪忧,Cd污染物位点超标率达到7.0%[2]。蔬菜产业作为近几年来我国绿色食品发展迅速的产业之一,是农业发展的重点方向,而产地环境Cd污染问题是蔬菜食用安全所面临的主要问题。土壤中Cd积累到一定水平时,就会毒害土壤-植物系统,致使土壤退化,作物生长迟缓、矮小,产量品质下降[3],更为严重的是重金属Cd会通过农作物的吸收富集作用,经食物链进入人体内,影响人体健康。人体的微量元素主要通过食物摄取,蔬菜中微量元素含量直接关系到人体健康。微量元素在作物体内含量虽少,但对植物生长发育起着重要的作用,它是组成酶、维生素和生长激素的成分,直接参与了植物体的新陈代谢。如Fe直接参与电子传递、氧化胁迫、固氮作用、激素合成等过程;Mn是许多酶类的辅因子,参与PSⅡ的氧化过程和蛋白质的代谢;Cu参与质体蓝素和虫漆酶的合成,同时在逆境胁迫和细胞壁代谢中发挥重要的作用;Zn是电子传递上重要辅酶因子,与植物抗氧化性和蛋白质转录有关[4]。许多研究表明,环境中的Cd浓度超过一定限量就会影响植株对微量元素的吸收,破坏植物体内微量元素的平衡。如,当根际介质中Cd含量逐渐升高时:蕹菜根系Zn含量增加,而Fe、Mn和Cu含量则呈现“先升后降”趋势[5];同时发现,上海青地上部Fe、Cu和Zn的含量则趋于降低[6]。
硒(Se)是人类、动物必需的微量元素,缺Se会导致人群中出现大骨头病、克山病等病症[7],同时Se也是植物生长发育的有益元素,在植物抗逆境胁迫(尤其是重金属胁迫)中发挥重要作用[8]。许多研究表明,在Cd污染环境中,施Se可以增强植物抗氧化性和对重金属的耐受性,但现阶段关于Se阻控植物富集重金属效应主要表现在降低重金属生物有效性,改善蔬菜产量、品质和养分平衡,提高逆境胁迫等方面[7,9-10],而对Se是如何调控微量元素在根系吸收及在木质部运输、积累的作用机理研究还不完善。基于上述原因,本试验采用水培形式,通过研究Cd胁迫下菜心(Brassica campestris L.ssp.chinensis var.Utilis)根系吸收(共质体途径与质外体途径)和木质部运输(木质液浓度)微量元素与菜心各组织微量元素分配的对比分析,以期揭示外源Se对菜心吸收与转运微量元素的影响,对探讨Se调控蔬菜微量元素的吸收、迁移和积累及其拮抗重金属胁迫机理具有一定的借鉴价值。
1 材料与方法
1.1 植株培养
水培试验于广东省农业科学院大丰实验基地中进行,供试材料为广东省主栽菜心品种——特选种19号四九菜心,购于广东省农业科学院蔬菜研究所。菜心种子经消毒、蒸馏水浸泡处理后播于湿润的纱布上,于28℃黑暗条件下萌发育苗一周。后挑选生长一致的幼苗移栽至装有6 L营养液的培养箱中进行预培养,每盆12株,完全营养液采用Hoagland-Arnon营养液。预培养 10 d 后,开始添加 Cd(1、5 μmol·L-1)和 Se(1、5 μmol·L-1),其中 Cd 以 CdCl2的形式加入,Se以Na2SeO3的形式加入。试验共设7个处理,分别为 CK、Cd1、Cd1Se1、Cd1Se5、Cd5、Cd5Se1、Cd5Se5,每个处理重复4次。每5 d更换一次营养液,持续通气,营养液pH用HCl和NaOH调至6.5~7.0;幼苗置于光周期为16 h/8 h、温度20~25℃环境下生长。待添加各处理培养3周后收集待测样品。
1.2 指标测定
1.2.1 植株干重的测定
收获的全株植物用自来水冲洗,以植株基部为准分离地上部及根部。根先置于20 mmol·L-1乙二胺四乙酸钠(EDTA-Na2)浸泡20 min,经蒸馏水冲洗后与地上部组织于105℃杀青30 min,68℃下烘干,用千分之一天平称重,备用。
1.2.2 地上部、根部微量元素含量和转移系数的测定
参照Wu等[11]的方法:准确称取植株不同部位干样0.2 g左右于50 mL烧杯中,加入10 mL混酸[硝酸∶高氯酸(V/V)=4∶1],在 180 ℃消解至澄清,用蒸馏水定容到 25 mL,利用 ICP-MS(Inductively Coupled Plasma Mass Spectrometry,Agilent 7900)测定各微量元素含量(mg·kg-1)。
转移系数(TF)=地上部各微量元素含量(mg·kg-1)/根部各微量元素含量(mg·kg-1)。
1.2.3 根部质外体、共质体及细胞壁中微量元素含量的测定
参照顾明华等[12]的方法:称取已切成2~3 mm的新鲜根部样品1.0 g左右,加入10 mL 5 mmol·L-1的CaCl2溶液(pH=6.0),振荡提取2.5 h后用纱布过滤,反复提取3次,得到质外体提取液。将过滤后的残根放置于-75℃下冷冻3 d,自然解冻破坏细胞膜,按上述质外体方法提取3次,得到共质体提取液。后将残根用蒸馏水洗涤3次,加入10 mL混酸消解得细胞壁提取液。利用ICP-MS(Agilent 7900)测定根部质外体、共质体及细胞壁中各微量元素含量(mg·kg-1)。
1.2.4 木质液微量元素浓度测定
参照吴志超[13]的方法:挑选6株长势一致的菜心植株,在离根部2 cm处用刀片切去菜心地上部,断茎处套上10 cm左右橡皮管,并将已弯好的一端引流至10 mL刻度管中,起始1~2 μL丢弃以免伤流干扰,采集时间为11:00—14:00。准确吸取0.5 mL收集液并定容,利用ICP-MS(Agilent 7900)测定木质部中微量元素浓度(mg·L-1)。
1.3 数据统计分析
实验采用Excel 2013进行数据处理,采用SPSS 19.0进行方差分析(ANOVA),应用Duncan新复极差法进行显著性分析,以小写英文字母表示多重比较结果。
2 结果与分析
2.1 Se对Cd胁迫下菜心地上部、根部生物量(DW)的影响
表1反映了Cd胁迫下,Se处理对菜心地上部、根部生物量的影响。由表可知,单一低Cd胁迫可提高菜心不同组织的生物量;而高Cd胁迫下则相反,地上部和根部干重相比CK分别减少了23.9%和16.7%。Cd胁迫下施加外源性Se处理后,对菜心生物量具有促进效果,其中低Cd胁迫下施用1 μmol·L-1Se时增幅最大,地上部、根部显著增加了29.5%和41.1%;高镉胁迫时,施Se后地上部、根部生物量有增加趋势,其中5 μmol·L-1Se时与单一高镉胁迫相比显著提高了21.6%和56.3%。
2.2 Se对Cd胁迫下菜心各组织(根部和地上部)微量元素(Fe、Mn、Cu、Zn)含量及转运系数的影响
表2为施Se对低镉和高镉胁迫下菜心地上部、根部微量元素含量及转移系数的影响。从表2可知,微量元素在根部含量均高于地上部,结果符合植物对矿质元素积累规律。低Cd胁迫时,地上部Mn和Zn的含量显著增加38.8%和22.1%;而高Cd胁迫时,地上部Fe、Mn、Cu、Zn含量与 CK相比则分别降低了7.4%、16.2%、20.2%和24.3%。另外,低浓度Cd胁迫时,菜心根部Fe和Cu含量上升,而Mn和Zn的含量降低;高浓度的Cd胁迫时,根系中Mn和Zn含量显著降低36.2%和17.3%,但Cu含量显著增加69.7%。另外,各微量元素的转移系数在低Cd胁迫时普遍大于高镉胁迫,此时Mn和Zn的转移系数与CK相比分别显著增加了59.6%和23.9%;而高浓度Cd对Cu转运的抑制作用最为明显,使其转移系数降低了53.9%。
同时从表2可知,Cd胁迫下,施Se处理改变了菜心对微量元素的吸收、运输。低Cd胁迫下,施用5 μmol·L-1Se时与单一低Cd胁迫相比,地上部Fe、Cu含量分别增加了18.7%和8.9%,而根部Fe和Cu则分别减少29.5%和20.4%;同时,地上部Mn、Zn在施1 μmol·L-1Se时显著降低 20.1%、14.0%,但根部含量变化不明显。高Cd胁迫下,施用5 μmol·L-1Se时地上部 Fe、Cu、Zn含量分别显著增加 19.8%、46.7%和26.2%,Mn增加7.3%;根部Mn、Cu、Zn含量则在施Se处理后增加趋势不显著,然而Fe含量在施1 μmol·L-1Se后显著降低了17.2%。另外,表中也可看出施Se处理后,菜心组织内Fe、Cu和Zn的转移系数有增加趋势,而对Mn的转移系数无明显影响。
2.3 Se对Cd胁迫下菜心根系质外体、共质体、细胞壁Fe、Mn、Cu、Zn 含量的影响
当环境中Cd浓度超标时,会对根系产生毒害作用,表3至表6反映了Cd胁迫下,施Se处理对根系微量元素在共质体、质外体和细胞壁中含量及其分配比例的影响。从表中可知,高浓度Cd胁迫下,质外体中Fe、Mn、Zn含量与CK相比分别降低了20.4%、23.8%和16.4%,共质体中则分别下降了20.7%、11.7%和27.2%,但此时根系质外体和共质体中Cu的含量却提高了48.2%和154.9%。另外,细胞壁中含量因元素种类不同表现出的差异不同,高Cd胁迫下,菜心细胞壁中Fe、Mn、Zn含量普遍降低,Cu含量却增加。同时,低Cd胁迫增强了微量元素向共质体的转移,普遍增加了共质体中各微量元素的分配比例;而高Cd胁迫时,共质体中Fe、Mn、Zn分配比例相较于CK有所降低,但Cu的分配比例从3.4%提高到7.4%。可见,镉胁迫破坏了微量元素在细胞壁中的平衡,尤其是在高Cd胁迫下。
从表3至表6中可得,低Cd胁迫下,根系各微量元素含量变化趋势随着施Se浓度的增加而不同,质外体中Mn、Cu、Zn以及共质体中Cu和Zn含量在施Se后增加差异不明显,而细胞壁中Mn含量则显著降低。施5 μmol·L-1Se时,与单一低Cd胁迫相比,共质体中Cu、Zn含量分别提高了21.2%和10.3%,细胞壁中Mn含量则降低了19.1%。高Cd胁迫时,质外体中 Fe、Cu、Zn 以及共质体中 Fe、Mn、Cu、Zn 含量随施Se浓度的增加均有不同程度的提高,说明施Se处理普遍增强了微量元素在根系中质外体和共质体途径的运输;施用 5 μmol·L-1Se时,菜心根系质外体中Fe、Cu、Zn含量与单一高Cd胁迫相比分别提高了16.9%、34.9%和26.0%,共质体中 Fe、Mn、Cu和 Zn含量则提高了13.3%、13.6%、41.9%和28.3%;另外,细胞壁中Fe和Cu含量在施Se后无明显变化,但施5 μmol·L-1Se时,细胞壁中Mn含量显著降低43.7%,且Zn含量提高了42.3%。同时,从表可知,低Cd胁迫下增大施Se浓度时,根系细胞壁中Mn和Cu的分配比例有下降趋势,质外体和共质体中的分配比例则相应增加。高Cd胁迫下,施Se处理普遍增大了质外体和共质体中Fe、Mn、Cu的分配比例,降低了细胞壁中这3种元素的比例,却增加了细胞壁中Zn的分配比例。
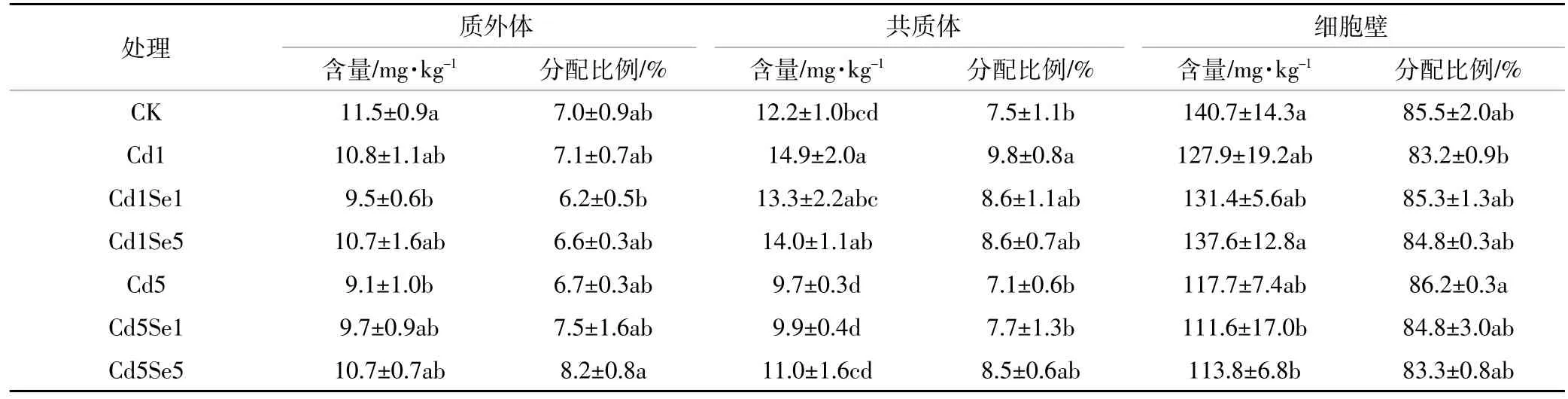
表3 根系质外体、共质体和细胞壁中Fe含量及分配比例Table 3 Concentration and distribution of Fe in the root apoplast,symplast,and cell walls
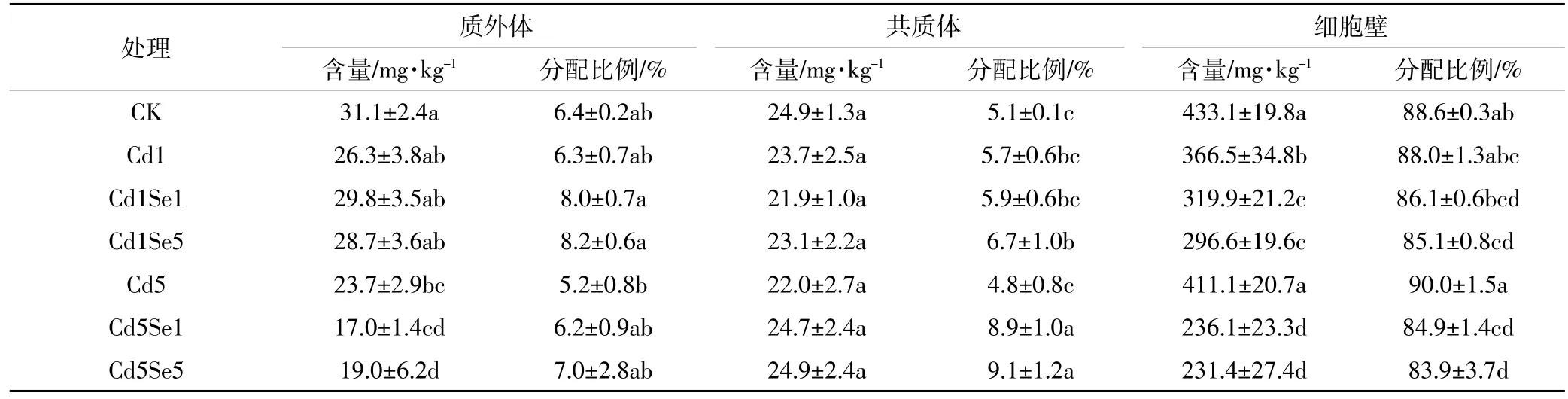
表4 根系质外体、共质体和细胞壁中Mn含量及分配比例Table 4 Concentration and distribution of Mn in the root apoplast,symplast,and cell walls
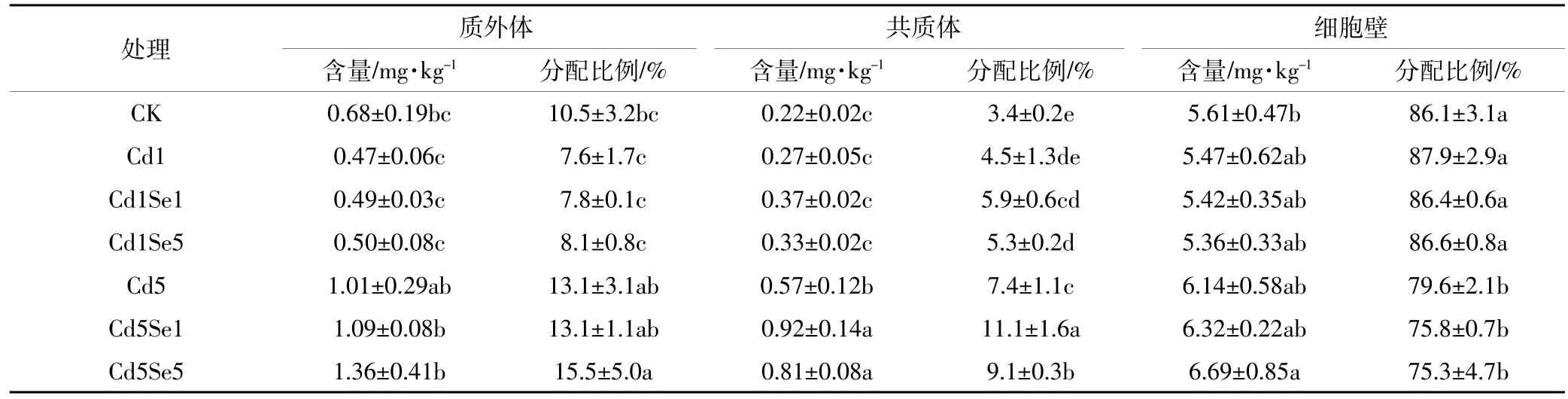
表5 根系质外体、共质体和细胞壁中Cu含量及分配比例Table 5 Concentration and distribution of Cu in the root apoplast,symplast,and cell walls
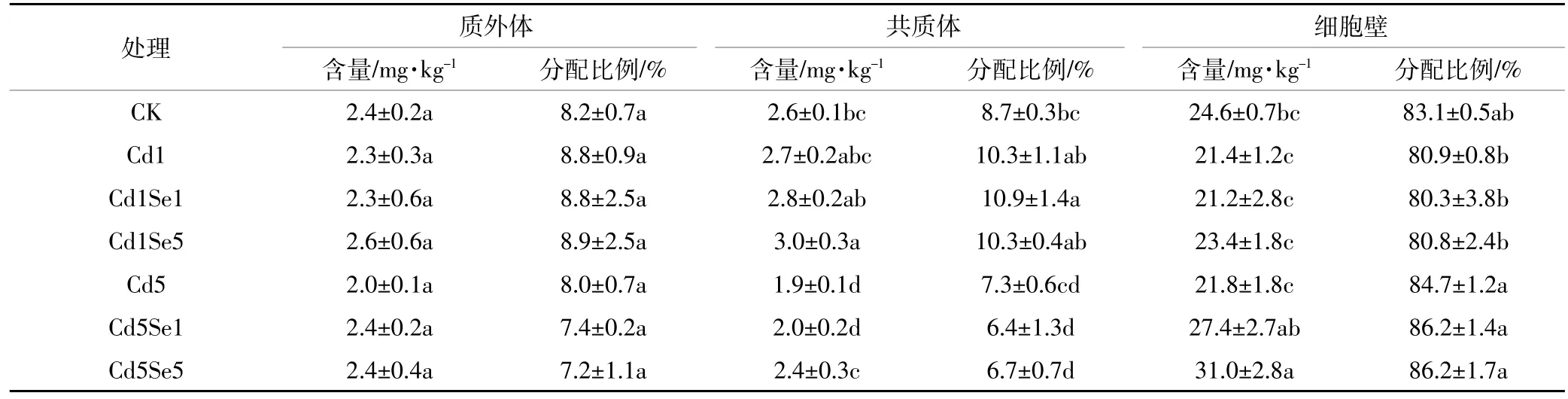
表6 根系质外体、共质体和细胞壁中Zn含量及分配比例Table 6 Concentration and distribution of Zn in the root apoplast,symplast,and cell walls
2.4 Se对 Cd胁迫下菜心木质部转运 Fe、Mn、Cu、Zn的影响
表7显示了不同施Se处理对木质液中微量元素浓度的影响。从表中可知,随着Cd胁迫加强,木质液中Fe、Mn和Zn浓度表现出“先升后降”趋势,而Cu表现降低趋势;5 μmol·L-1Cd 胁迫时木质液中 Fe、Cu、Zn浓度分别降低了6.1%、26.0%、22.4%。低Cd胁迫下施Se后,木质液中各微量元素浓度变化趋势有所不同,但都未表现出显著差异。高Cd胁迫下施Se处理后,木质液中Fe、Cu、Zn浓度有增加趋势,且施用5 μmol·L-1Se时与单一高Cd胁迫相比增加了9.8%、44.2%和16.9%,其中Cu和Zn达到显著水平,但对Mn无明显影响。
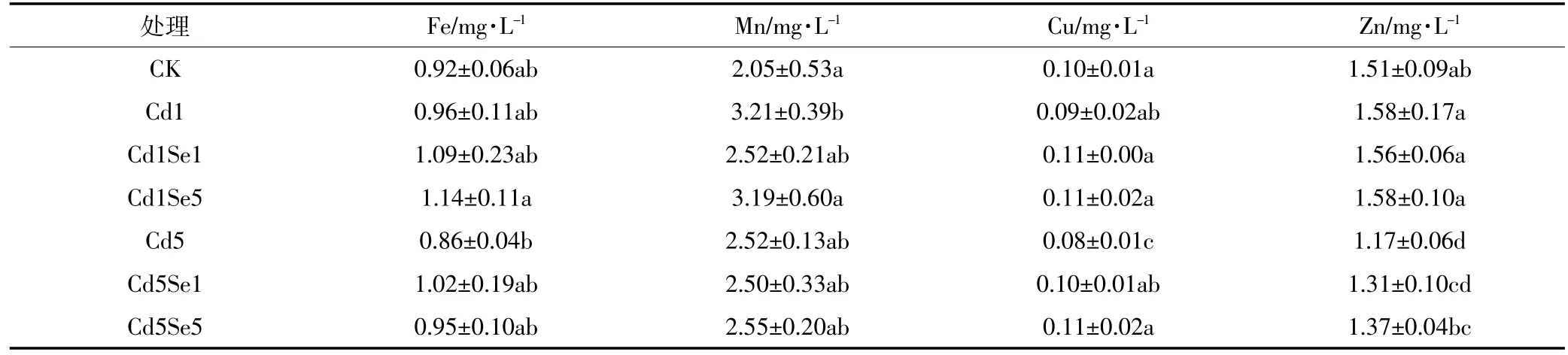
表7 木质部中微量元素的浓度Table 7 Concentration of trace elements in the xylem
3 讨论
3.1 Se对Cd胁迫下菜心生物量的影响
重金属Cd胁迫时,蔬菜生物量是反映植株耐受强度的重要指标。有研究表明,低浓度的Cd对蔬菜生长有一定的刺激作用,如1 μmol·L-1Cd能够促进小白菜的生长,提高其生物量[14]。然而,高浓度Cd会使无机营养元素流失,干扰植物生理机能,影响植物新陈代谢,进而减少其生物量[15]。另外,已有相关研究表明,Se可缓解菠菜[16]、小白菜[17]等蔬菜Cd毒害,增加作物生物量。本试验中,单一Cd处理菜心时,地上部、根部干重变化也出现“低促高抑”的现象,说明5 μmol·L-1的Cd已对菜心产生毒害作用。另外,Cd胁迫下施Se处理后,地上部、根部干重均有不同程度增加。可见,施Se对促进菜心生长、缓解镉毒害具有一定的效果。
3.2 Se对Cd胁迫下菜心组织微量元素分配的影响
蔬菜根系是吸收微量元素的重要部位,而地上部(可食部分)含量是判断营养价值高低的重要指标,微量元素平衡遭到破坏则直接影响到植株的生长发育及其质量品质。为了更好地比较菜心不同部位微量元素吸收与转运的关系,用转移系数来表示微量元素(Fe、Mn、Cu、Zn)在菜心组织中的分布特征规律,反映出根部吸收后转运到地上部的能力[18]。许多文献指出,镉胁迫能影响植物对微量元素的吸收与转运,主要是影响微量元素在体内的含量和浓度。张菊平等[19]对小白菜的研究中发现在5 μmol·L-1Cd胁迫时,地上部Mn、Zn、Cu及根部Fe、Zn含量受到抑制,且存在品种差异。Dong 等[20]则发现 10 μmol·L-1Cd 显著降低了番茄幼苗根系中Fe、Mn、Cu、Zn含量,而叶片Fe、Mn、Zn降低较少,说明Cd严重影响了根部细胞膜的选择透过性,使根系对某些微量元素的积累减少。本研究结果表明,菜心地上部对Fe、Mn、Cu、Zn以及根部对Fe、Mn的吸收在Cd胁迫下均表现出“低促高抑”的现象,说明本试验中添加的1 μmol·L-1Cd浓度并未超过菜心的耐受阈值,具有促进植株生长的效果,提高了对微量元素的吸收积累,表现出微量元素转移系数增大的现象。添加5 μmol·L-1Cd时,抑制了菜心对微量元素的吸收,可能是因为Cd抑制了根系呼吸作用,阻碍了根部物质代谢和能量转换,使细胞膜结构和功能发生变化,降低了根部对微量元素主动吸收的能力。同时,Cd被转移至叶片细胞时,会破坏叶绿体结构和功能,使叶绿素分解,并对细胞产生不同程度的毒害作用,致使植物对微量元素吸收代谢紊乱[21],向地上部转移能力下降。
施Se对植株体内微量元素吸收的影响,与植株种类、施用部位以及施用浓度等有关。许多研究显示,植物体内Se与Mn和Zn浓度表现为正相关,而与Fe表现为负相关[22]。陈平等[23]的研究结果表明,Se处理可以显著增加Cd胁迫下水稻叶片中Fe、Mn、Zn含量。刘春梅等[24]对寒地水稻的研究则得出,施Se后,叶片Zn含量下降,Fe含量增加;根系Zn、Cu含量下降,而Fe、Mn含量增加。李登超等[25]研究也表明,施Se处理增加了白菜地上部Mn和Zn含量,却降低了根部Fe、Mn、Zn元素的吸收。由此可见,Se对植物体内微量元素作用关系错综复杂,不同研究结果表现出矛盾性。本研究结果与前人结果也有不同之处:低Cd胁迫施Se促进了Fe和Cu由根部向地上部的转移;Cd对菜心产生毒害作用后,施Se对提高地上部Fe、Cu、Zn和降低根系Fe含量具有一定效果,但对根部Mn、Cu、Zn含量影响不大。可能是因为加Se后与Cd形成难溶性复合物,降低了活性氧自由基的生成及根尖细胞凋亡比例,缓解了对细胞膜的损伤,提高了抗氧化系统相关酶的活性[27];同时Se增加了菜心中叶绿素含量,使叶绿体代谢运输及能量代谢得到保障[16],恢复了植物的正常生长,进而增强地上部对微量元素的运输积累。
3.3 Se对Cd胁迫下菜心微量元素在根系及木质部中转运的影响
根系吸收微量元素经共质体途径和质外体途径运输到木质部,后经木质部到达地上部茎叶,其转移率取决于多个因素。根系共质体途径是指元素从植物根毛细胞膜通道进入,利用细胞间的胞间连丝从一个细胞质移动到另一个细胞质,经皮层、内皮层等进入根系内导管,移动速度慢;质外体途径则是根系周围的元素经根系吸收后,沿着细胞壁中的空隙进入木质部和韧皮部,这种方式阻力小、速率快。根系细胞壁其特殊的滞留作用也是影响元素吸收运输的重要机制[18,26]。一方面,土壤溶液中多数以离子态形式存在的微量元素先被吸附于根组织表面,然后经质外体途径或共质体途径进入根组织维管束的木质部导管向上运输[28],而根系细胞壁和液泡的吸收、固定作用以及共质体和质外体途径中运输效率影响着根系对微量元素的积累。已有研究表明,Cd在经根系吸收时,会在表皮细胞沉积,毒害植物根系[18],影响微量元素在根部的运输。本研究结果显示,随着Cd胁迫加强,质外体和共质体中Fe、Mn、Zn含量及分配比例降低,却使Cu含量及分配比例上升。说明Cd胁迫破坏了根系之间微量元素的平衡,使某些微量元素由经细胞壁进入质外体和共质体途径受阻,有学者认为Cd进入细胞的途径和 Ca2+、Fe2+、Mn2+、Zn2+等营养元素相似,共享在细胞膜上的离子通道/运输蛋白,如K+和Ca2+通道[29],易与其他金属元素竞争相同的载体结合位点;而Cu含量上升可能是因为Cd改变了细胞膜的通透性,导致大量Cu进入共质体造成的[30]。另外,施Se处理后,普遍提高了Fe、Mn、Cu在两种途径中的分配比例,增强了菜心微量元素在根系中质外体和共质体途径的运输。研究表明,Se可以和重金属Cd形成Se-Cd复合物,从而减少了植株对Cd的吸收,降低其生理生化毒性[10]。因此,Se缓解了Cd和其他离子的竞争关系,进而促进了微量元素进入根细胞,提高了其在共质体中分配比例。另外施Se也可以提高植物 GSH-Px、SOD、CAT、GR 等活性[16,27,31],通过提高谷胱甘肽循环途径来减轻或消除根系细胞膜因Cd引起的损伤,在限制对Cd吸收以及保护细胞膜结构和功能的完整性等方面具有重要作用[32],从而使根系微量元素含量达到动态平衡。以上结果说明,微量元素在质外体途径和共质体途径含量和分配比例的改变是Se影响根系对其吸收、转运的一个重要机制。
另一方面,根系吸收的微量元素通过木质部运输到达地上部茎叶,木质部装载、运输和卸载是影响微量元素向地上部转移的重要因素,金属离子或其化合物从根细胞装载进入木质部和卸载要经跨膜运输,这就需要通过相应载体来完成这个过程[33]。本研究结果显示,低Cd胁迫下,除Mn外,其他微量元素变化不明显,但高Cd胁迫使木质部中微量元素浓度普遍降低。可能原因是Cd胁迫达到一定程度后,影响了木质部中相关氨基酸或载体的表达,减弱了木质部装载、运输能力,进而抑制了微量元素在木质部的运输;而施Se选择性地提高了木质部中载体含量,进而增强了对某些微量元素的运输。已有研究表明,Cd处理显著改变木质液中有机酸、氨基酸的组成和含量[34],而不同有机酸种类对调控植物微量元素运输机制存在较大差异,如柠檬酸运输载体可以促进Fe从根系向地上部运输[35],尼克酰胺对番茄木质部中转运Cu过程有重要作用[36],植物Fe载体和DMA影响木质部中Fe和Zn的运输[37]。本研究显示,施Se后,木质液中Fe、Cu、Zn浓度有增加趋势,但是对Mn的吸收与转运不显著。由此推断,Cd胁迫下施Se处理可能加强了木质液中相关氨基酸和有机酸的表达,如柠檬酸、Fe载体等,从而影响元素在木质部中的转运,但是这一过程受Cd胁迫强度、施Se浓度和元素种类的影响。但是,有关Se在缓解Cd胁迫中与其他元素的相互作用的研究报告很少,另外Se对木质部中载体种类和含量的影响也有待进一步研究。
4 结论
(1)低浓度Cd对菜心具有刺激效果,对微量元素在根系吸收、木质部运输以及地上部含量积累等方面均有一定的促进作用,可提高菜心生物量。但随着Cd胁迫加强,根部对微量元素吸收紊乱,木质部浓度普遍降低,进而表现出地上部含量下降,生物量减少。
(2)施Se可以缓解Cd胁迫对菜心造成的毒害作用,低Cd胁迫施Se促进了Fe和Cu由根部向地上部的转移;而在 5 μmol·L-1Cd胁迫时,施 Se则对提高地上部Fe、Cu和Zn含量,降低根系Fe含量具有一定效果。
(3)高Cd胁迫下施Se处理,增加了质外体中Fe、Cu、Zn 以及共质体中 Fe、Mn、Cu、Zn 含量;有利于提高质外体和共质体中Fe、Mn、Cu的分配比例。
(4)施Se对低Cd胁迫下木质液中微量元素浓度的增加作用不明显;而高Cd胁迫下施Se后木质液中Fe、Cu、Zn的浓度有上升趋势,维持了地上部微量元素供应的平衡。
[1]Nordberg G F.Historical perspectives on cadmium toxicology[J].Toxicology and Applied Pharmacology,2009,238(3):192-200.
[2]环境保护部.环境保护部和国土资源部发布全国土壤污染状况调查公报[EB/OL].[2014-04-17].http://www.mlr.gov.cn/xwdt/jrxw/201404/t20140417_1312998.htm.Ministry of Environmental Protection.The ministry of environmental protection and the ministry of land and resources issued the national survey on soil pollution[EB/OL].[2014-04-17].http://www.mlr.gov.cn/xwdt/jrxw/201404/t20140417_1312998.htm.
[3]郭爱珍,陈 斌,程 曼,等.我国蔬菜重金属污染现状及防控措施[J].山西农业科学,2016,44(4):560-564.GUO Ai-zhen,CHEN Bin,CHENG Man,et al.Present situation of heavy metal pollution of vegetables in China and its prevention and control measures[J].Journal of Shanxi Agricultural Sciences,2016,44(4):560-564.
[4]Dalcorso G,Manara A,Furini A.An overview of heavy metal challenge in plants:From roots to shoots[J].Metallomics,2013,5(9):1117-1132.
[5]张 娟.镉胁迫条件下蕹菜对土壤铁锰铜锌钙镁的吸收与富集[D].福建:福建农林大学,2011.ZHANG Juan.The uptake and accumulation of Fe,Mn,Cu,Zn,Ca and Mg by water spinach(Lpomoea aquatic Forsk.)from soils under cadmium stress[D].Fujian:Fujian Agriculture University,2011.
[6]宋阿琳.小白菜对镉胁迫的响应及硅缓解镉毒害的机制[D].南京:南京农业大学,2009.SONG A-lin.Responses of Brassica chinensis L.to cadmium stress and silicon-alleviated cadmium toxicity[D].Nanjing:Nanjing Agriculture University,2009.
[7]庞晓辰,王 辉,吴泽赢,等.硒对水稻镉毒害的影响及其机制的研究[J].农业环境科学学报,2014,33(9):1679-1685.PANG Xiao-chen,WANG Hui,WU Ze-ying,et al.Alleviation by selenium of cadmium toxicity to rice and its mechanisms[J].Journal of A-gro-Environment Science,2014,33(9):1679-1685.
[8]Harabawy A,Mosleh Y.The role of vitamins A,C,E and selenium as antioxidants against genotoxicity and cytotoxicity of cadmium,copper,lead and zinc on erythrocytes of Nile tilapia[J].Ecotoxicology and Environmental Safety,2014,104:28-35.
[9]Feng R W,Wei C W,Tu S X.The roles of selenium in protecting plants againstabioticstresses[J].EnvironmentalandExperimentalBotany,2013,87(87):58-68.
[10]袁思莉,余 垚,万亚男,等.硒缓解植物重金属胁迫和积累的机制[J].农业资源与环境学报,2014,31(6):545-550.YUAN Si-li,YU Yao,WAN Ya-nan,et al.Mechanisms of selenium mitigating stress and accumulation of heavy metals in plants[J].Journal of Agricultural Resources and Environment,2014,31(6):545-550.
[11]Wu Z C,Wang F H,Liu S,et al.Comparative responses to silicon and selenium in relation to cadmium uptake,compartmentation in roots,and xylem transport in flowering Chinese cabbage(Brassica campestris L.ssp.Chinensis var.utilis)under cadmium stress[J].Environmental and Experimental Botany,2016,131:173-180.
[12]顾明华,黎晓峰.硅对减轻水稻的铝胁迫效应及其机理研究[J].植物营养与肥料学报,2002,8(3):360-366.GU Ming-hua,LI Xiao-feng.Effect of silicon on alleviation of aluminum toxicity and corresponded mechanisms in rice[J].Plant Nutrition and Fertilizer Science,2002,8(3):360-366.
[13]吴志超.高低镉积累油菜品种筛选及其生化机制研究[D].武汉:华中农业大学,2015.WU Zhi-chao.Screening of high/low cadmium accumulation Brassica napus cultivars and research on the biochemical mechanisms[D].Wuhan:Huazhong Agriculture University,2015.
[14]于方明,仇荣亮,汤叶涛,等.Cd对小白菜生长及氮素代谢的影响研究[J].环境科学,2008,29(2):506-511.YU Fang-ming,QIU Rong-liang,TANG Ye-tao,et al.Effects of cadmium on the growth and nitrogen metabolism in Brassica chinensis[J].Environmental Science,2008,29(2):506-511.
[15]Maria C,Cristina M,Jose M,et al.Cadmium toxicity affects photosynthesis and plant growth at different levels[J].Acta Physiol Plant,2013,35(4):1281-1289.
[16]郭 锋,樊文华,冯两蕊,等.硒对镉胁迫下菠菜生理特性、元素含量及镉吸收转运的影响[J].环境科学学报,2014,34(2):524-531.GUO Feng,FAN Wen-hua,FENG Liang-rui,et al.Effects of selenium(Se)on the physiological characteristics,element contents,uptake and transportation of Cd in spinach under Cd stress[J].Acta Scientise Circumstantise,2014,34(2):524-531.
[17]刘 达,涂路遥,赵小虎,等.镉污染土壤施硒对植物生长及根际镉化学行为的影响[J].环境科学学报,2016,36(3):999-1005.LIU Da,TU Lu-yao,ZHAO Xiao-hu,et al.Effect of selenium application to the cadmium-polluted rhizosphere on plant growth and chemical behavior of cadmium[J].Acta Scientise Circumstantise,2016,36(3):999-1005.
[18]王晓娟,王文斌,杨 龙,等.重金属镉(Cd)在植物体内的转运途径及其调控机制[J].生态学报,2015,35(23):7921-7929.WANG Xiao-juan,WANG Wen-bin,YANG Long,et al.Transport pathways of cadmium(Cd)and its regulatory mechanisms in plant[J].ActaEcologicaSinica,2015,35(23):7921-7929.
[19]张菊平,崔文朋,焦新菊,等.低浓度镉对小白菜生长及营养元素吸收积累的影响[J].江西农业大学学报,2011,33(1):22-28.ZHANG Ju-ping,CUI Wen-peng,JIAO Xin-ju,et al.Effect of low concentration cadmium on the growth and accumulation of nutrient elements in pakchoi[J].Acta Agriculturae University Jiangxiensis,2011,33(1):22-28.
[20]Dong J,Wu F B,Zhang G P.Influence of cadmium on antioxidant capacity and four microelement concentrations in tomato seedlings(Lycopersicon esculentum)[J].Chemosphere,2006,64(10):1659-1666.
[21]赵 巍,张联合,郁飞燕,等.镉对水稻幼苗积累微量元素的影响[J].山东农业科学,2011,11:63-66,76.ZHAO Wei,ZHANG Liao-he,YU Fei-yan,et al.Effect of cadmium on accumulation of microelements in rice seedling[J].Shandong Agricultural Sciences,2011,11:63-66,76.
[22]Arvy M P,Thiersault M,Doireau P.Relationship between selenium,micronutrients,carbohydrates,and alkaloid accumulation in Catharanthus roseus cells[J].Journalof Plant Nutrition,1995,18(8):1535-1546.
[23]陈 平,吴秀峰,张伟锋,等.硒对镉胁迫下水稻幼苗叶片元素含量的影响[J].中国生态农业学报,2006,14(3):114-117.CHEN Ping,WU Xiu-feng,ZHANG Wei-feng,et al.Effects of Se on some element contents in leaves of rice seedling under Cd stress[J].Chinese Journal of Eco-Agriculture,2006,14(3):114-117.
[24]刘春梅,罗胜国,王孟雪,等.硒对镉胁迫下寒地水稻 Cd、Zn、Fe、Cu、Mn 含量的影响[J].水土保持学报,2014,28(6):136-142.LIU Chun-mei,LUO Sheng-guo,WANG Meng-xue,et al.Effects of selenium on cadmium,zinc,iron,copper,manganese content in rice under cadmium stress in cold climate[J].Journal of Soil and Water Conservation,2014,28(6):136-142.
[25]李登超,朱祝军,徐志豪,等.硒对小白菜生长和养分吸收的影响[J].植物营养与肥料学报,2003,9(3):353-358.LI Deng-chao,ZHU Zhu-jun,XU Zhi-hao,et al.Effects of selenium on the growth and nutrient absorption of pakchoi[J].Plant Nutrition and Fertilizer Science,2003,9(3):353-358.
[26]王学华,戴 力.作物根系镉滞留作用及其生理生化机制[J].中国农业科学,2016,49(22):4323-4341.WANG Xue-hua,DAI Li.Immobilization effect and its physiology and biochemical mechanism of the cadmium in crop roots[J].Scientia AgriculturaSinica,2016,49(22):4323-4341.
[27]Wu Z C,Liu S,Zhao J,et al.Comparative responses to silicon and selenium in relation to antioxidant enzyme system and the glutathioneascorbate cycle in flowering Chinese cabbage(Brassica campestris L.ssp.Chinensis var.utilis)under cadmium stress[J].Environmental and Experimental Botany,2017,131:1-11.
[28]白艳波,李 娇,张宝龙,等.干旱胁迫对植物矿质元素影响的研究进展[J].生物技术通报,2013,31(3):15-18.BAI Yan-bo,LI Jiao,ZHANG Bao-long,et al.Research advance on effect of drought stress on mineral elements of plant[J].Biotechnology Bulletin,2013,31(3):15-18.
[29]Perfus-Barbeoch L,Leonhardt N,Vavasseur A,et al.Heavy metal toxicity:cadmium permeates through calcium channels and disturbs the plant water status[J].The Plant Journal,2002,32(4):539-548.
[30]郭 智,原海燕,奥岩松.镉胁迫对龙葵幼苗光合特性和营养元素吸收的影响[J].生态环境学报,2009,18(3):824-829.GUO Zhi,YUAN Hai-yan,AO Yan-song.Effect of cadmium on photosynthesis and nutrient elements uptake of Solanum nigrum L.seedling[J].Ecology and Environmental Sciences,2009,18(3):824-829.
[31]林匡飞,徐小清,金 霞,等.硒对水稻的生态毒理效应及临界指标研究[J].应用生态学报,2005,16(4):678-682.LIN Kuang-fei,XU Xiao-qing,JIN Xia,et al.Eco-toxicological effects of selenium and its critical value on Oryza sativa[J].Chinese Journal of Applied Ecology,2005,16(4):678-682.
[32]彭 玲,贾 芬,田小平,等.硒对油菜根尖镉胁迫的缓解作用[J].环境科学学报,2015,35(8):2597-2604.PENG Ling,JIA Fen,TIAN Xiao-ping,et al.Alleviation of cadmium stress on root tip of rape seedling by selenium[J].Acta Scientise Circumstantise,2015,35(8):2597-2604.
[33]陈 飞.大麦镉吸收与运转机制的研究[D].杭州:浙江大学,2009.CHEN Fei.Physiological and molecular mechanism of cadmium uptake and translocation in barley[D].Hangzhou:Zhejiang University,2009.
[34]Nakamura S,Akiyama C,Sasaki T,et al.Effect of cadmium on the chemical composition of xylem exudate from oilseed rape plants(Brassicanapus L.)[J].Soil Science and Plant Nutrition,2008,54(1):118-127.
[35]Morrissy J,Baxter I R,Lee J,et al.The ferroportin metal efflux proteins function in iron and cobalt homeostasis in Arabidopsis[J].Plant Cell,2009,21:3326-3338.
[36]Pich A,Scholz G.Translocation of copper and other micronutrients in tomato plants(Lycopersicon esculentum Mill.):Nicotianamine-stimulated copper transport in the xylem[J].Experimental Botany,1996,47(1):41-47.
[37]Suzuki M,Tsukamoto T,Inoue H,et al.Deoxymugineic acid increases Zn translocation in Zn-deficient rice plants[J].Plant Mol Biol,2008,66(6):609-617.
猜你喜欢
植物研究(2021年2期)2021-02-26
山西农业科学(2021年1期)2021-01-18
热带农业科学(2018年2期)2018-06-21
速读·中旬(2017年11期)2017-11-11
上海故事(2017年10期)2017-10-19
理论导刊(2017年8期)2017-08-30
饮食与健康·下旬刊(2016年9期)2016-05-14
西湖(2016年9期)2016-05-14
天津农业科学(2015年11期)2015-12-03
安徽农学通报(2014年7期)2014-04-29
